Impact of Soil Field Water Capacity on Secondary Metabolites, Phenylalanine Ammonia-lyase (PAL), Maliondialdehyde (MDA) and Photosynthetic Responses of Malaysian Kacip Fatimah (Labisia pumila Benth)

Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolics and Flavonoids

{kind=link}
{kind=link}
{kind=link}
| Varieties | Evapotranspiration | Total Phenolics | Total Flavonoids | Anthocyanin |
|---|---|---|---|---|
| replacement | (mg gallic acid/g dry weight) | (mg rutin/g dry weight) | (mg petunidin/ g fresh weight) | |
| Alata | 100% | 0.81 ± 0.01 d | 0.52 ± 0.01 c | 0.72 ± 0.01 d |
| 75% | 1.23 ± 0.12 c | 0.67 ± 0.03 b | 0.81 ± 0.02 c | |
| 50% | 1.75 ± 0.02 b | 0.75 ± 0.13 a | 0.92 ± 0.01 a | |
| 25% | 1.72 ± 0.14 b | 0.64 ± 0.12 b | 0.90 ± 0.04 a | |
| Pumila | 100% | 0.79 ± 0.13 d | 0.57 ± 0.15 c | 0.69 ± 0.01 d |
| 75% | 1.27 ± 0.03 c | 0.69 ± 0.16 b | 0.79 ± 0.03 c | |
| 50% | 1.89 ± 0.03 a | 0.75 ± 0.11 a | 0.97 ± 0.05 a | |
| 25% | 1.83 ± 0.07 a | 0.71 ± 0.06 a | 0.88 ± 0.04 b |
2.2. Anthocyanins
| Parameters | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1.Total phenolics | 1.000 | |||||||||||
| 2.Total flavonoids | 0.987 * | 1.000 | ||||||||||
| 3.Anthocyanin | 0.876 * | 0.888 * | 1.000 | |||||||||
| 4.Photosynthesis | –0.879 * | –0.855 * | –0.871 * | 1.000 | ||||||||
| 5.AQY | –0.876 * | –0.901 * | –0.899 * | 0.978 * | 1.000 | |||||||
| 6.Dark respiration | 0.934 * | 0.876 * | 0.789 * | 0.234 | 0.113 | 1.000 | ||||||
| 7.fv/fm | –0.874 * | –0.786 * | –0.699 * | 0.766 * | 0.433 | –0.445 | 1.000 | |||||
| 8.RWC | –0.854 * | –0.921 * | –0.865 * | 0.897 * | 0.223 | 0.778 * | 0.899 * | 1.000 | ||||
| 9.Carbohydrate | 0.976 * | 0.986 * | 0.765 * | –0.876 * | 0.677 * | –0.765 * | –0.556 * | –0.876 * | 1.000 | |||
| 10.Chlorophyll | –0.766 * | –0.786 * | –0.865 * | -0.987 * | 0.332 | 0.344 | 0.776 * | –0.444 | –0.978 * | 1.000 | ||
| 11.PAL | 0.932 * | 0.876 * | 0.812 * | –0.781 * | 0.021 | 0.021 | 0.034 | 0.002 | 0.788 * | 0.234 | 1.000 | |
| 12.MDA | 0.911 * | 0.885 * | 0.934 * | –0.886 * | 0.032 | 0.043 | 0.001 * | 0.012 | 0.876 * | –0.002 | 0.987 * | 1.000 |
| Evapotranspiration | Net Photosynthesis | Apparent quantum yield | Dark respiration rate (Rd) | Maximum efficiency of photosystem II |
|---|---|---|---|---|
| replacement | (A) | (ɸ) | (µmol/m2/s) | (fv/fm) |
| (µmol/m2/s) | (µmol/m2/s) | |||
| 100% | 10.75 ± 0.01 a | 0.08 ± 0.01 a | 4.61 ± 0.21 d | 0.84 ± 0.01 a |
| 75% | 7.21 ± 0.22 b | 0.06 ± 0.02 b | 12.71 ± 0.23 c | 0.81 ± 0.02 b |
| 50% | 3.41 ± 0.12 c | 0.04 ± 0.07 c | 14.75 ± 0.13 b | 0.72 ± 0.06 c |
| 25% | 1.72 ± 0.13 d | 0.01 ± 0.04 d | 15.64 ± 0.14 a | 0.66 ± 0.04 d |
2.3. Leaf gas Exchange Properties
2.4. Relative Leaf Water Content
| Varieties | Evapotranspiration | Relative leaf water | Sucrose content |
|---|---|---|---|
| replacement | content (%) | mg/g dry weight | |
| Alata | 100% | 89.31 ± 1.01 b | 27.52 ± 3.01 d |
| 75% | 87.32 ± 0.12 b | 32.41 ± 0.93 c | |
| 50% | 79.21 ± 0.32 c | 64.31 ± 0.83 b | |
| 25% | 72.72 ± 0.74 d | 63.64 ± 1.12 b | |
| Pumila | 100% | 94.79 ± 0.93 a | 27.57 ± 3.15 d |
| 75% | 85.27 ± 0.54 b | 35.69 ± 5.16 c | |
| 50% | 77.89 ± 1.02 c | 67.75 ± 2.11 a | |
| 25% | 70.83 ± 2.07 d | 66.32 ± 0.07 a |
2.5. Soluble Sugar (Sucrose) Content
2.6. Total Chlorophyll Content

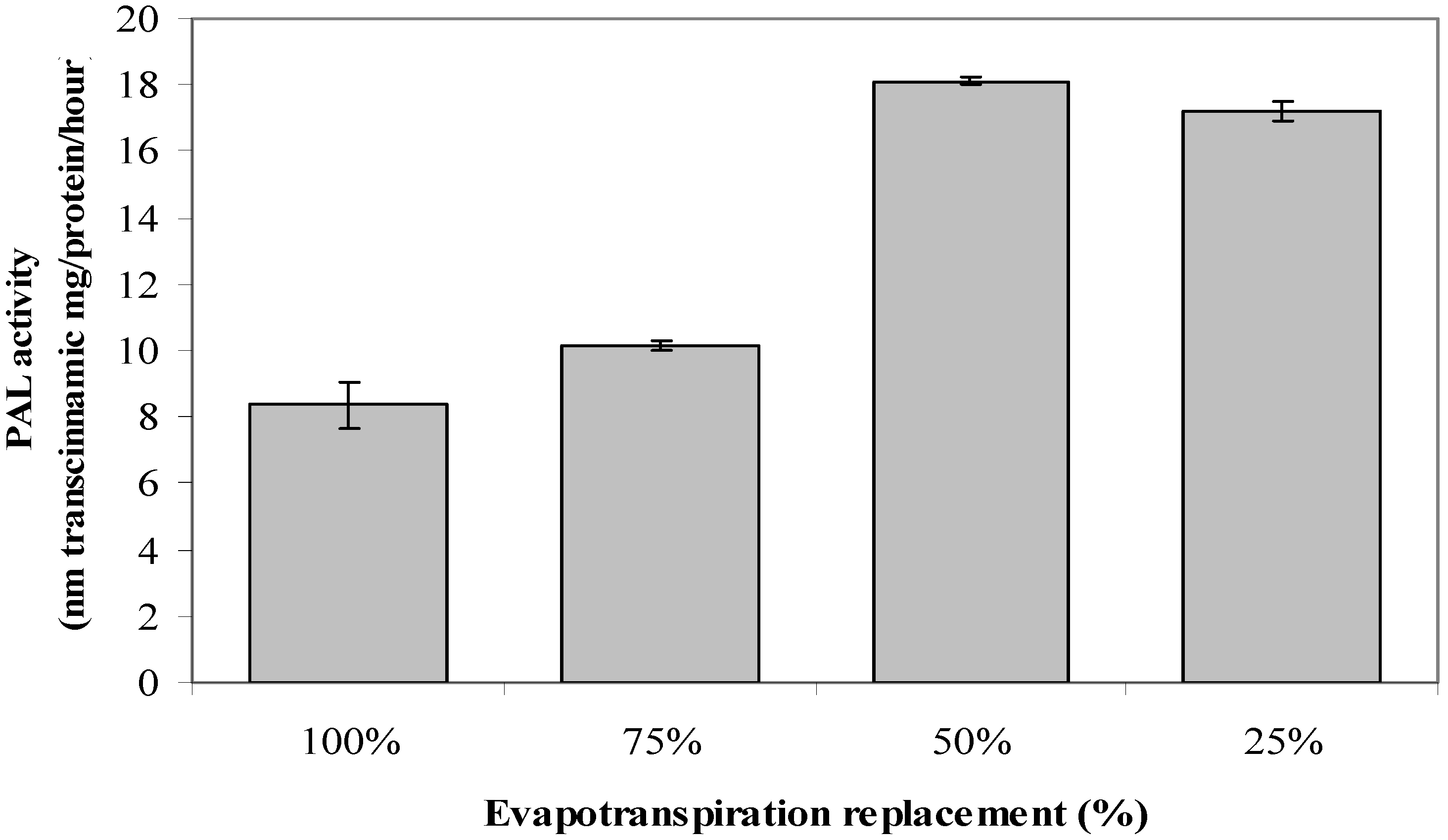
2.7. Phenylalanine Ammonia-Lyase Activity

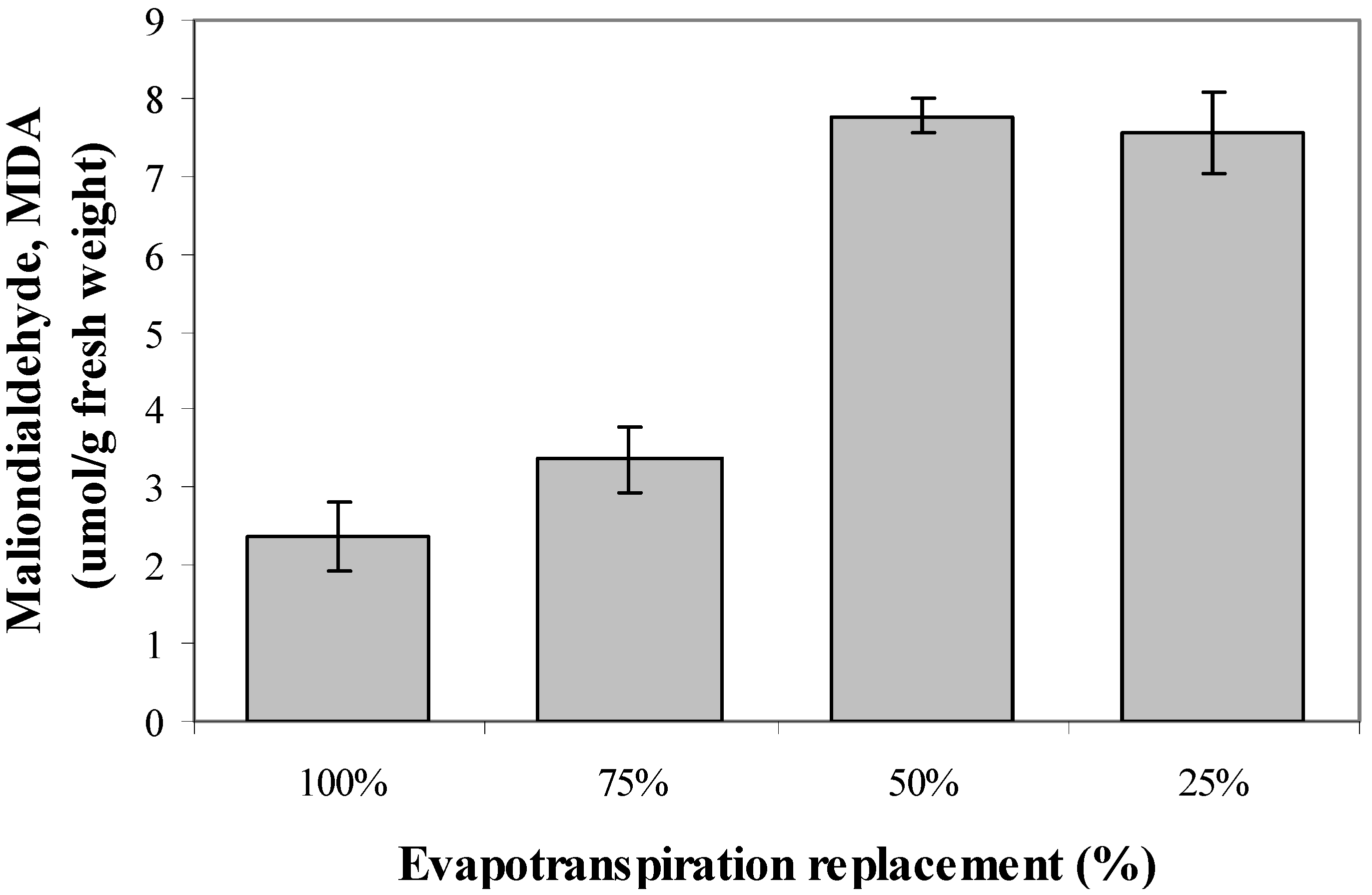
2.8. Lipid Peroxidation Activity

3. Experimental
3.1. Plant Material and Maintenance
3.2. Total Phenolics and Flavonoids Quantification
3.3. Anthocyanins Content
3.4. Leaf Gas Exchange Measurement
3.5. Chlorophyll Fluorescences Measurement
3.6. Relative Leaf Water Content
3.7. Sucrose Determination
3.8. Chlorophyll Content
3.9. Phenylalanine-Ammonia-Lyase (PAL) Activity Determination
3.10. Malondialdehyde (MDA) Content Determination
3.11. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Harborne, J.B. Introduction to Ecological Biochemistry,3rd ed.; Academic Press: London, UK, 1993. [Google Scholar]
- Heath, M.C.; Boller, T. Biotic interactions levels of complexity in plant interactions with herbivores, pathogens and mutualists. Curr. Opin. Plant Biol. 2002, 5, 277–278. [Google Scholar] [CrossRef]
- Jain, D.C.; Mathur, A.K.; Gupta, M.M.; Singh, A.K.; Verma, R.K.; Gupta, A.P.; Kumar, S. Isolation of high artemisinin-yielding clones of Artemisia annua. Phytochemistry 1996, 43, 993–901. [Google Scholar] [CrossRef]
- Ghershenzon, J. Changes in levels of plant secondary metabolites under water and nutrient stress. In Recent Advances in Phytochemistry-Phytochemical Adaptations to Stress; Timmermann, B.N., Steelin, C., Loewus, F.A., Eds.; Plenum Press: New York, NY, USA, 1984; pp. 273–220. [Google Scholar]
- Hansen, U.; Seufert, G. Terpenoid emission from Citrus sinensis (L.) OSBECK under drought stress. Phys. Chem. Earth 1999, 24, 681–687. [Google Scholar]
- Marchese, J.A.; Rehder, V.L.G. The influence of temperature in the production of artemisinin in Artemisia annua L. Braz. J. Med. Plant 2001, 4, 89–93. [Google Scholar]
- Magiero, E.C.; Assmann, J.M.; Marchese, J.A.; Capelin, D.; Paladini, M.V.; Trezzi, M.M. Allelopathic effect of Artemisia annua L. on the germination and initial development of lettuce (Lactuca sativa L.) and wild poinsettia (Euphorbia heterophylla L.) seedlings. Braz. J. Med. Plant 2009, 11, 317–324. [Google Scholar]
- Jaafar, H.Z.E.; Mohamed, H.N.B.; Rahmat, A. Accumulation and partitioning of total phenols in two varieties of Labisia pumila Benth. under manipulation of greenhouse irradiance. Acta Hort. 2008, 797, 387–392. [Google Scholar]
- Sulaiman, B.; Mansor, M.; Jaafar, A. Some medicinal plants in mount Bubu Peral. In Proceeding in Medicinal Products from Tropical Rain Forest; Shaari, K., Abdul, K.A., Ali, M., Eds.; Forest Research Institute of Malaysia: Kepong, Malaysia, 1992; pp. 37–43. [Google Scholar]
- Jamia, A.J.; Ibrahim, J.; Khairana, H.; Juriyati, H. Perkembangan Penyelidikan dan Pembangunan Kacip Fatimah; New Dimension in Complementary Health Care: Kuala Lumpur, Malaysia, 2004; pp. 13–19. [Google Scholar]
- Ferreira, J.F.S. Nutrient deficiency in the production of artemisinin, dihydroartemisinic acid, and artemisinic acid in Artemisia annua L. J. Agric. Food Chem. 2007, 55, 1686–1694. [Google Scholar] [CrossRef]
- Marchese, J.A.; Rehder, V.L.G.; Casiraghi, V.; Tedesco, A.C.; Lira, R. Flowering in plants of Artemisia annua L. standed to different conditions of photoperiod and temperature. Acta Hort. 2002, 569, 275–280. [Google Scholar]
- Putalun, W.; Luealon, W.; De-Eknamkul, W.; Tanaka, H.; Shoyama, Y. Improvement of artemisinin production by chitosan in hairy root cultures of Artemisia annua L. Biotechnol. Lett. 2007, 29, 1143–1146. [Google Scholar] [CrossRef]
- Stewart, A.J.; Chapman, W.; Jenkins, G.I.; Graham, I.; Martin, T.; Crozier, A. The effect of nitrogen and phosphorous deficiency on flavonol accumulation in plant tissues. Plant Cell Environ. 2001, 24, 1189–1197. [Google Scholar] [CrossRef]
- Winkel, S.B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Mosaleeyanon, K.; Zobayed, S.M.A.; Afreen, F.; Kozai, T. Relationships between net photosynthetic rate and secondary metabolite contents in St. John’s wort. Plant Sci. 2005, 169, 523–531. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Abdul Rahman, Z. The relationship between phenolics and flavonoids production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth.under high CO2 and nitrogen fertilization. Molecules 2011, 16, 162–174. [Google Scholar]
- Couceiro, M.A.; Afreen, F.; Zobayed, S.M.A.; Kozai, T. Variation in concentrations of major bioactive compounds of St. John’s wort: Effects of harvesting time, temperature and germoplasm. Plant Sci. 2006, 170, 128–134. [Google Scholar]
- Zobayed, S.M.A.; Afreen, F.; Kozai, T. Temperature stress can alter the photosynthetic efficiency and secondary metabolite concentrations in St. John’s wort. Plant Physiol. Bioch. 2005, 43, 977–984. [Google Scholar]
- Franz, C. Nutrient and water management for medicinal and aromatic plants. Acta Hort. 1983, 132, 203–215. [Google Scholar]
- Palevitch, D. Recent advances in the cultivation of medicinal plants. Acta Hort. 1987, 208, 29–35. [Google Scholar]
- Marchese, J.A.; Figueira, G.M. The use of pre and post-harvest technologies and good agricultural practices in the production of medicinal and aromatic plants. Braz. J. Med. Plant 2005, 7, 86–96. [Google Scholar]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: to grow or defend. Quart. Rev. Biol. 1992, 67, 283–235. [Google Scholar]
- Charles, D.I.; Simon, J.E.; Shock, C.C.; Feibert, E.B.G.; Smith, R.M. Effect of water stress and post-harvest handling on artemisinin content in the leaves of Artemisia annua L. In Proceedings of the Second International Symposium: New Crops, Exploration, Research and Commercialization, Indianapolis, USA, October 1991; Janick J. Simon, J.E., Ed.; John Wiley and Sons Inc.: New York, NY, USA, 1993; pp. 640–643. [Google Scholar]
- Jaleel, C.A.; Sankar, B.; Murali, P.V.; Gomathinayagam, M.; Lakshmanan, G.M.A.; Panneerselvam, R. Water deficit stress effects on reactive oxygen metabolism in Catharanthus roseus; impacts on ajmalicine accumulation. Colloid. Surfaces B 2008, 62, 105–111. [Google Scholar] [CrossRef]
- Lee, S.C.; Norliza, A.L.; Sze, Y.L.; Chew, T.L.; Mohd, R.S.; Ramlan, A.Z. Flavonols and phenolics from Labisia pumila (Kacip Fatimah). Food Chem. 2011, 127, 1186–1192. [Google Scholar] [CrossRef]
- Zaizuhana, S.; Puteri, J.N.M.B.; Noral’ashikin, Y.; Muhammad, H.; Rohana, A.B.; Zakiah, I. The in vivo rodent micronucleus assay of Kacip Fatimah (Labisia pumila) extract. Trop. Biomed. 2006, 23, 214–219. [Google Scholar]
- Jaafar, H.Z.E.; Mohamed, H.N.B.; Rahmat, A. Accumulation and partitioning of total phenols in two varieties of Labisia pumila Benth. under manipulation of greenhouse irradiance. Acta Hort. 2008, 797, 387–392. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.E. Photosynthetic capacity, photochemical efficiency and chlorophyll content of three varieties of Labisia pumila benth. Exposed to open field and greenhouse growing conditions. Acta Physiol. Plant 2011, 33, 2179–2185. [Google Scholar] [CrossRef]
- Fortier, E.; Desjardins, Y.; Tremblay, N.; Bélec, C.; Côté, M. Effects of exposure to soluble fraction of industrial solid waste on lipid peroxidation and DNA methylation in erythrocytes of Oreochromis niloticus, as assessed by quantification of MDA and m5dC rates. Acta Hort. 2010, 856, 55–62. [Google Scholar]
- Azhar, N.; Hussain, B.; Ashraf, M.Y.; Abbasi, K.Y. Water stress mediated changes in growth, physiology and secondary metabolites of desi ajwain (Trachyspermum Ammi L.). Pak. J. Bot. 2011, 43, 15–19. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Zaharah, A.R. Effects of nitrogen fertilization on synthesis of primary and secondary metabolites in three varieties of Kacip Fatimah (Labisia pumila Blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef]
- Oh, M.M.; Trick, H.N.; Rajashekar, C.B. Secondary metabolism and antioxidant are involved in environmental adaptation and stress tolerance in lettuce. J. Plant Physiol. 2009, 166, 180–199. [Google Scholar] [CrossRef]
- Harrison, K.; Were, L.M. Effect of gamma irradiation on total phenolics content, yield and antioxidant capacity of almond skin extract. Food Chem. 2007, 102, 932–937. [Google Scholar] [CrossRef]
- Palliotti, A.; Poni, S.; Silvestroni, O.; Tombesi, S.; Bernizzoni, F. Morpho-structural and physiological performance of Sangiovese and Montepulciano cvv. (Vitis vinifera) under non-limiting water supply conditions. Funct. Plant Biol. 2010, 38, 888–898. [Google Scholar]
- Wang, S.Y.; Jiao, H. Scavenging capacity of berry crops on superoxide radicals, hydrogen peroxide, hydroxyl radicals and singlet oxygen. J. Agric. Food Chem. 2000, 48, 5677–5684. [Google Scholar] [CrossRef]
- Wang, H.; Nair, M.G.; Strasburg, G.M.; Chang, Y.C.; Booren, A.M.; Gray, J.I.; DeWitt, D.L. Antioxidant and anti-inflammatory activities of anthocyanins and their aglycon, cyanidin, from tart cherries. J. Nat. Prod. 1999, 62, 294–296. [Google Scholar] [CrossRef]
- Tamura, H.; Yamagami, A. Antioxidative activity of monoacylated anthocyanins isolated from Muscat bailey A grape. J. Agric. Food Chem. 1994, 42, 1612–1615. [Google Scholar] [CrossRef]
- Asada, K. Production and action of active oxygen species in photosynthetic tissues. In Causes of Photooxidative Stress and Amelioration of Defense System in Plants; Foyer, C.H., Mullineaux, P.M., Eds.; CRS Press: Boca Raton, FL, USA, 1994; pp. 77–103. [Google Scholar]
- Moghaddam, S.S.; Jaafar, H.B.; Aziz, M.A.; Ibrahim, R.; Rahmat, A.B.; Philip, E. Flavonoid and leaf gas exchange responses of Centella asiatica to acute gamma irradiation and carbon dioxide enrichment under controlled environment conditions. Molecules 2011, 16, 8930–8944. [Google Scholar] [CrossRef]
- Schreiner, M.; Beyene, B.; Krumbein, A.; Stützel, H. Ontogenetic changes of 2-propenyl and 3-Lndolylmethyl glucosinolates in Brassica carinata leaves as affected by water supply. J. Agric. Food Chem. 2009, 57, 7259–7263. [Google Scholar] [CrossRef]
- Xiao, X.; Yang, F.; Zhang, S.; Korpelainen, H.; Li, C. Physiological and proteomic responses of two contrasting Populus cathayana populations to drought stress. Physiol. Plant. 2009, 136, 150–168. [Google Scholar] [CrossRef]
- Szabó, B.; Lakatos, Á.; Koszegi, T.; Botz, L. Investigation of abiogenic stress-induced alterations in the level of secondary metabolites in poppy plants (Papaver somniferum L.). Acta Biol. Hung. 2008, 59, 425–438. [Google Scholar] [CrossRef]
- Jaafar, H.Z.E. Impact of Environmental Stress on Reproductive Development in Sweet pepper (Capsicum anuum). Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1995. [Google Scholar]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Asmah, R. Antioxidant activities, total phenolics and flavonoids Content in two varieties of malaysia young ginger (Zingiber officinale Roscoe). Molecules 2010, 15, 4324–4333. [Google Scholar] [CrossRef]
- Guo, R.; Yuan, G.; Wang, Q. Effects of sucrose and mannitol accumulation of health promoting component and activity of metabolic enzyme in brocolli sprout. Sci. Hort. 2011, 128, 159–165. [Google Scholar] [CrossRef]
- Kubula, J.; Sriamoopunns, S.; Meeso, N. Phytochemical, vitamin C and sugar content of thai wild fruits. Food Chem. 2011, 126, 972–981. [Google Scholar] [CrossRef]
- Veenashi, B.R.; Muralikrishna, G. In vitro antioxidant activity of xylo-oligosharides derived from cereal and millet brans - a competitive study. Food Chem. 2011, 126, 1475–1481. [Google Scholar] [CrossRef]
- Jones, C.G.; Hartley, S.E. A protein competition model of phenolic allocation. Oikos 1999, 86, 27–44. [Google Scholar] [CrossRef]
- De-souza, A.P.; Gaspar, M.; Da-silva, E.A.; Ulian, E.C.; Waclawovsky, A.J.; Nishiyama, M.Y.; Dos, M.R.V.; Bucjkeridge, M.S. Elevated CO2 increases photosynthesis, biomass and productivity and modifies gene expression in sugarcane. Plant Cell Environ. 2008, 31, 1116. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E. The influence of carbohydrate, protein and phenylanine ammonia lyase on up-regulation of production of secondary metabolites (total phenolics and flavonoid) in Labisia pumila (Blume) Fern-Vill (Kacip Fatimah) under high CO2 and different nitrogen levels. Molecules 2011, 16, 4172–4190. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Hawa, Z.E.J. Carbon dioxide fertilization enhanced antioxidant compounds in Malaysian Kacip Fatimah (Labisia pumila Blume). Molecules 2011, 16, 6068–6081. [Google Scholar] [CrossRef]
- Flohr, L.; Fuzinatto, C.F.; Melegari, S.P.; Matias, W.G. Effects of exposure to soluble fraction of industrial solid waste on lipid peroxidation and DNA methylation in erythrocytes of Oreochromis niloticus, as assessed by quantification of MDA and m5dC rates. Ecotoxicol. Environ. Saf. 2012, 76, 63–70. [Google Scholar] [CrossRef]
- Klapwijk, D.; De Lint, P.J.A.L. Fresh weight and flowering of tomato plants as influenced by container type and watering condition. Acta Hort. 1974, 39, 237–247. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z. Reduced Photoinhibition under Low Irradiance Enhanced Kacip Fatimah (Labisia pumila Benth) Secondary Metabolites, Phenyl Alanine Lyase and Antioxidant Activity. Int. J. Mol. Sci. 2012, 13, 5290–5306. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E. Enhancement of leaf gas exchange and primary metabolites, up-regulate the production of secondary metabolites of Labisia Pumila Blume seedlings under carbon dioxide enrichment. Molecules 2011, 16, 3761–3777. [Google Scholar] [CrossRef]
- Bharti, A.K.; Khurana, J.P. Molecular characterization of transparent testa (tt) mutants of Arabidopsis thaliana (ecotype Estland) impaired in flavonoid biosynthesic pathway. Plant Sci. 2003, 165, 1321–1332. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Haniff, M.H.; Raffi, M.Y. Changes in growth and photosynthetic patterns of oil palm seedling exposed to short term CO2 enrichment in a closed top chamber. Acta Physiol. Plant 2010, 32, 305–313. [Google Scholar] [CrossRef]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurements of water deficits in leaves. New Phytol. 1950, 49, 81–87. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E. The relationship of nitrogen and C/N on secondary metabolites and antioxidant activities in three varieties of Malaysia Kacip Fatimah (Labisia pumila Blume). Molecules 2011, 16, 5514–5526. [Google Scholar]
- Jaafar, H.Z.E.; Ibrahim, M.H.; Por, L.S. Effects of CO2 enrichment on accumulation of total phenols, flavonoid and chlorophyll content in two varieties of Labisia pumila Benth. exposed to different shade levels. In Proceedings of International Conference on Balanced Nutrient Management for Tropical Agriculture, Kuantan, Pahang, Maalysia, February 2010; UPM: Kuala Lumpur, Malaysia, 2010; pp. 112–114. [Google Scholar]
- Martinez-Tellez, M.A.; Lafuente, M.T. Effects of high temperature conditioning on ethylene, phenylalanine ammonia lyase, peroxidase and polyphenol oxidase in flavedo of chilled “Fortune” mandarin fruit. J. Plant Physiol. 1997, 150, 674–678. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Asmah, R.; Zaharah, A.R. Involvement of Nitrogen on Flavonoids, Glutathione, Anthocyanin, Ascorbic Acid and Antioxidant Activities of Malaysian Medicinal Plant Labisia pumila Blume (Kacip Fatimah). Int. J. Mol. Sci. 2012, 13, 393–408. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.E. Primary, Secondary Metabolites, H2O2, Malondialdehyde and Photosynthetic Responses of Orthosiphon stimaneus Benth. to Different Irradiance Levels. Molecules 2012, 17, 1159–1176. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z. Impact of Elevated Carbon Dioxide on Primary, Secondary Metabolites and Antioxidant Responses of Eleais guineensis Jacq. (Oil Palm) Seedlings. Molecules 2012, 17, 5195–5211. [Google Scholar]
- Jaafar, H.Z.; Ibrahim, M.H.; Karimi, E. Phenolics and Flavonoids Compounds, Phenylanine Ammonia Lyase and Antioxidant Activity Responses to Elevated CO2 in Labisia pumila (Myrisinaceae). Molecules 2012, 17, 6331–6347. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jaafar, H.Z.E.; Ibrahim, M.H.; Mohamad Fakri, N.F. Impact of Soil Field Water Capacity on Secondary Metabolites, Phenylalanine Ammonia-lyase (PAL), Maliondialdehyde (MDA) and Photosynthetic Responses of Malaysian Kacip Fatimah (Labisia pumila Benth). Molecules 2012, 17, 7305-7322. https://doi.org/10.3390/molecules17067305
Jaafar HZE, Ibrahim MH, Mohamad Fakri NF. Impact of Soil Field Water Capacity on Secondary Metabolites, Phenylalanine Ammonia-lyase (PAL), Maliondialdehyde (MDA) and Photosynthetic Responses of Malaysian Kacip Fatimah (Labisia pumila Benth). Molecules. 2012; 17(6):7305-7322. https://doi.org/10.3390/molecules17067305
Chicago/Turabian StyleJaafar, Hawa Z. E., Mohd Hafiz Ibrahim, and Nur Farhana Mohamad Fakri. 2012. "Impact of Soil Field Water Capacity on Secondary Metabolites, Phenylalanine Ammonia-lyase (PAL), Maliondialdehyde (MDA) and Photosynthetic Responses of Malaysian Kacip Fatimah (Labisia pumila Benth)" Molecules 17, no. 6: 7305-7322. https://doi.org/10.3390/molecules17067305