Protective Effect of Polyphenols Extract of Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) on Hypercholesterolemia-Induced Oxidative Stress in Rats

Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical in Adlay Phenolic Extract (APE)
2.2. Effect of APE on the Body Weight Gain, Food Intake, and Liver Weight of Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body weight gain (g) | Food intake (g/d) | Feed efficiency (%) | Liver weight (g) | Liver index |
|---|---|---|---|---|---|
| Control | 136 ± 19 | 31.45 ± 1.39 | 16.03 ± 2.18 | 11.86 ± 1.22 | 2.82 ± 0.20 |
| Chol | 134 ± 32 | 31.24 ± 2.41 | 15.93 ± 3.77 | 12.48 ± 1.84 | 2.99 ± 0.20 |
| Chol/LAPE | 132 ± 24 | 30.87 ± 1.15 | 15.79 ± 2.93 | 12.33 ± 0.95 | 2.96 ± 0.12 |
| Chol/MAPE | 142 ± 24 | 31.91 ± 0.97 | 16.48 ± 2.80 | 13.19 ± 1.50 | 3.10 ± 0.11 |
| Chol/HAPE | 141 ± 32 | 29.89 ± 1.76 | 17.44 ± 3.91 | 12.32 ± 1.51 | 3.04 ± 0.10 |
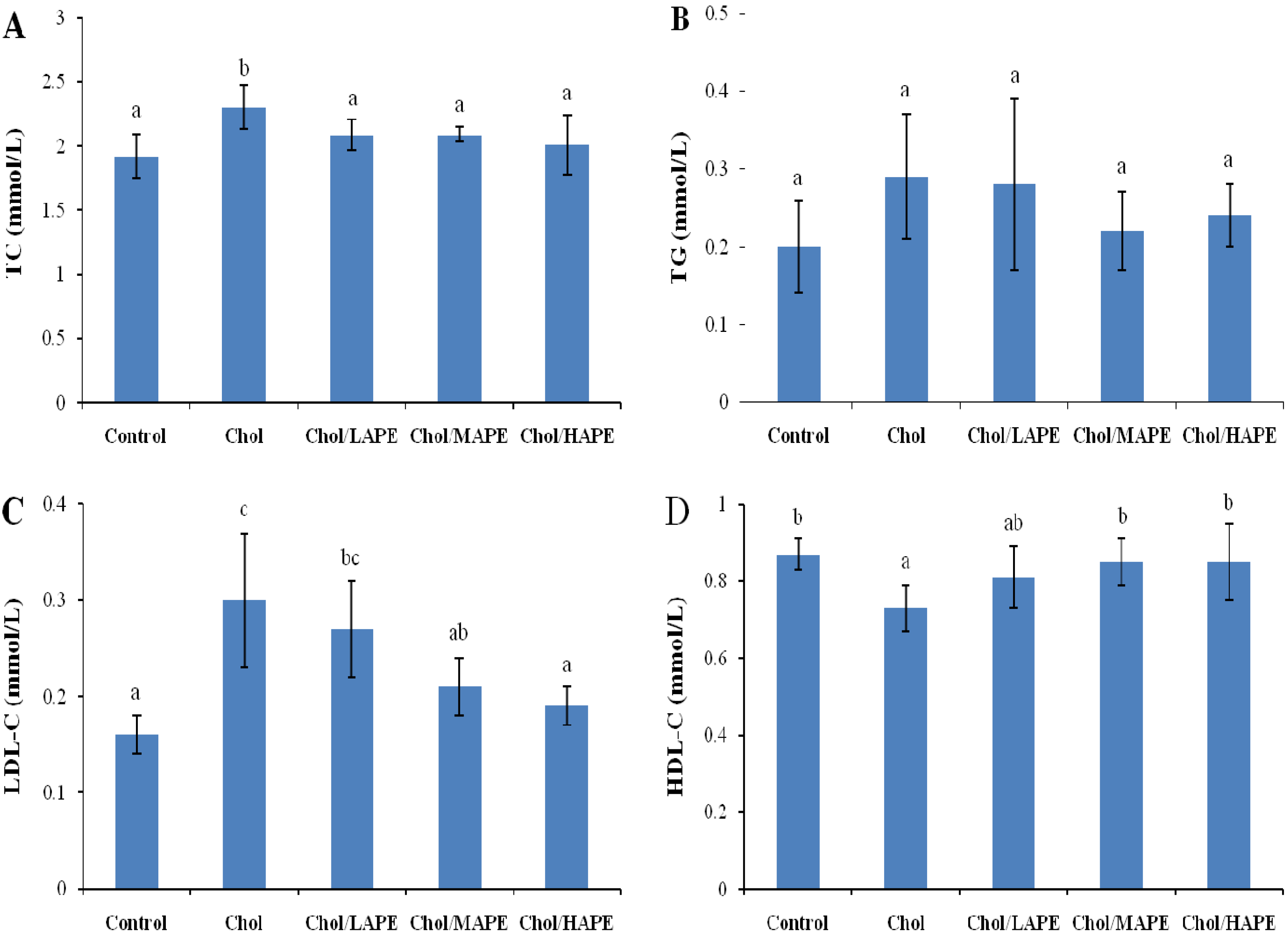
2.3. Effect of APE on Serum Lipid Profile of Rats

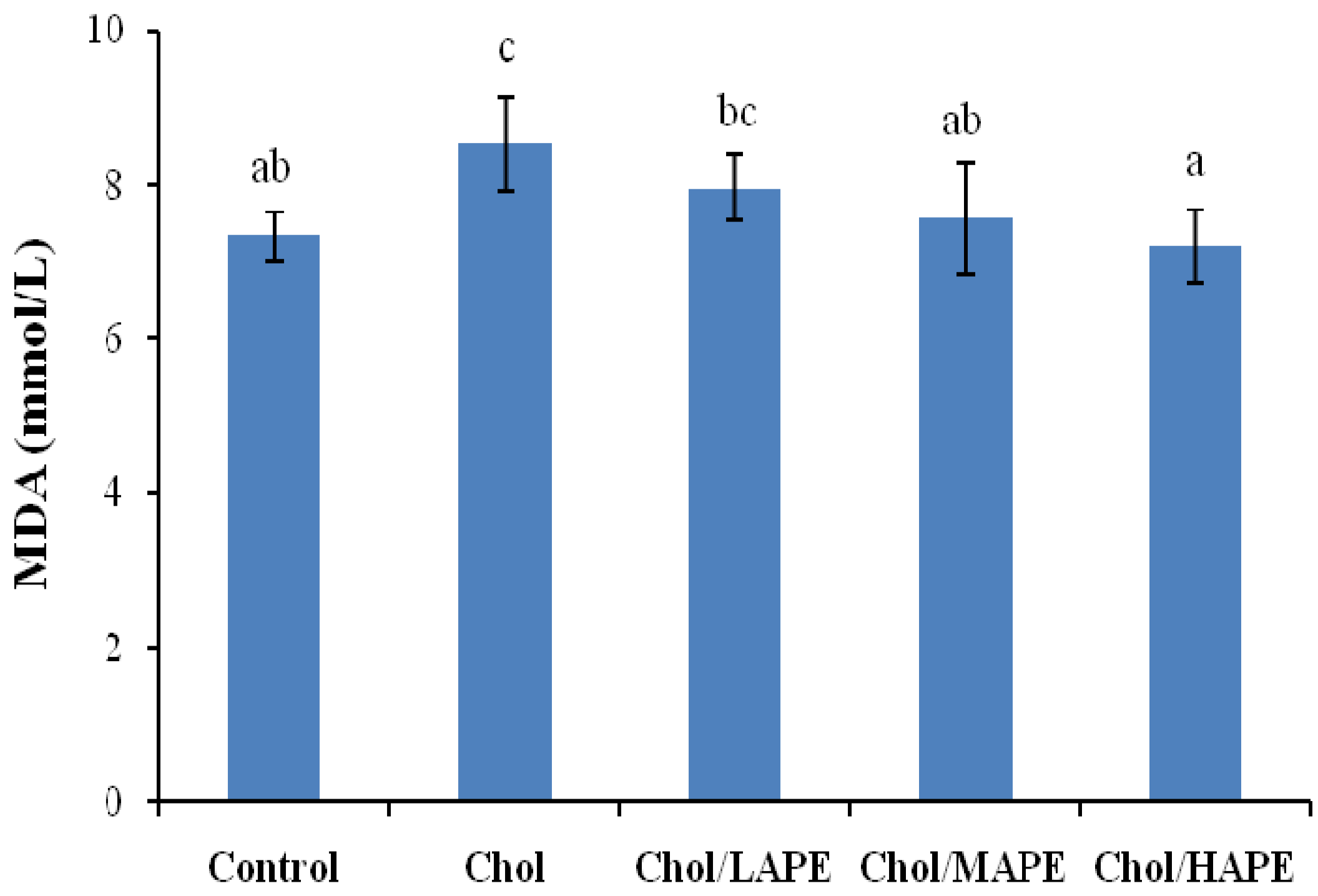
2.4. Effect of APE on Lipid Peroxidation of Hypercholesterolemic Rats

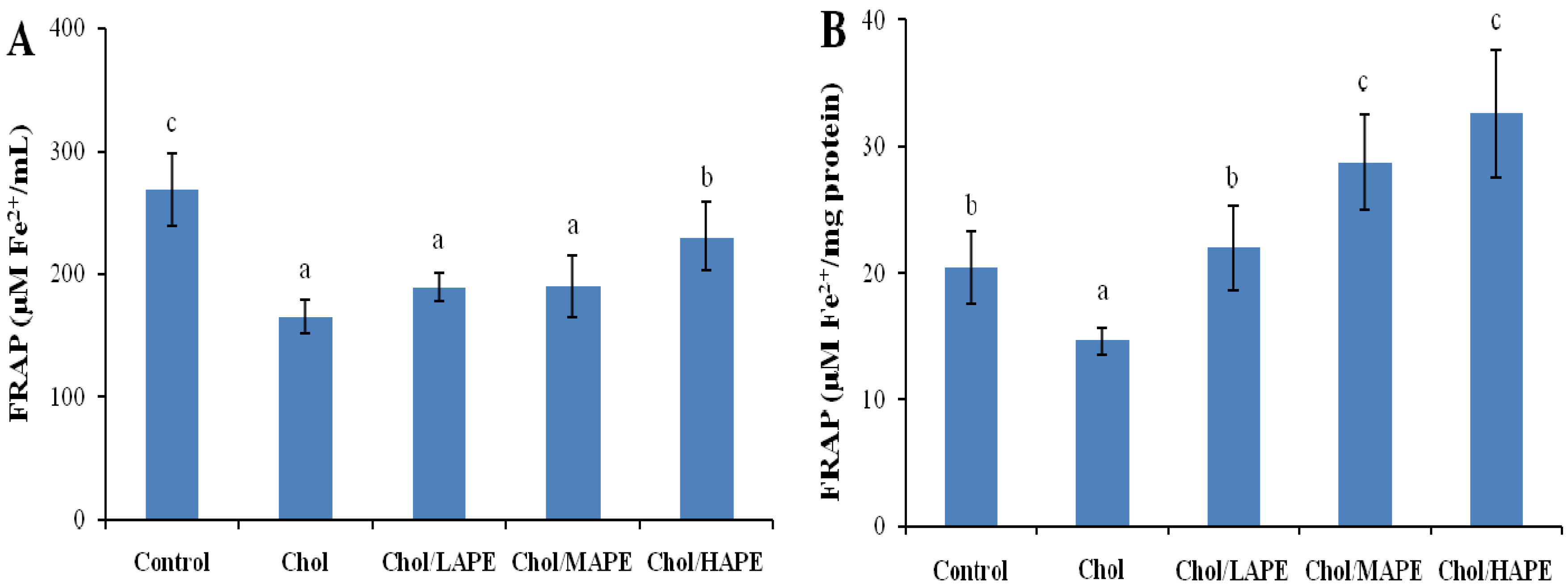
2.5. Effect of APE on the Antioxidant Status of Hypercholesterolemic Rats


3. Experimental
3.1. Chemicals
3.2. Preparation of APE
3.3. Determination of the Total Phenolic and Total Flavonoid Content
3.4. Animals and in Vivo Study Design
| Ingredients | Basal diet (g/kg) | High-cholesterol diet (g/kg) |
|---|---|---|
| Casein | 182 | 180 |
| Soybean oil | 61 | 60 |
| Wheat starch | 687 | 680 |
| Cholesterol | 0 | 10 |
| Vitamin mixture | 10 | 10 |
| Mineral mixture | 60 | 60 |
3.5. Serum Lipids Assay
3.6. Lipid Peroxidation and Antioxidant Profiles Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
Conflict of Interest
References
- Gould, A.L.; Rossouw, J.E.; Santanello, N.C.; Heyse, J.F.; Furberg, C.D. Cholesterol reduction yields clinical benefit—A new look at old data. Circulation 1995, 91, 2274–2282. [Google Scholar] [CrossRef]
- Yang, R.-L.; Shi, Y.-H.; Hao, G.; Li, W.; Le, G.-W. Increasing oxidative stress with progressive hyperlipidemia in human: Relation between malondialdehyde and atherogenic index. J. Clin. Biochem. Nutr. 2008, 43, 154–158. [Google Scholar] [CrossRef]
- Young, I.; McEneny, J. Lipoprotein oxidation and atherosclerosis. Biochem. Soc. Trans. 2001, 29, 358–362. [Google Scholar] [CrossRef]
- Zeng, Y.; Song, J.-X.; Shen, X.-C. Herbal remedies supply a novel prospect for the treatment of atherosclerosis: A review of current mechanism studies. Phytother. Res. 2012, 26, 159–167. [Google Scholar] [CrossRef]
- Huang, S.; Chen, Y.; Chiang, W. Amino acids, fatty acids and proximate composition of the seed of adlay. Food Sci. 1994, 21, 67–74. [Google Scholar]
- Li, S.Z. Compendium of Materia Medica, 1st ed; China Radio International Press: Beijing, China, 1994; pp. 1639–1643. [Google Scholar]
- Chen, H.-J.; Chung, C.-P.; Chiang, W.; Lin, Y.-L. Anti-inflammatory effects and chemical study of a flavonoid-enriched fraction from adlay bran. Food Chem. 2011, 126, 1741–1748. [Google Scholar] [CrossRef]
- Chung, C.-P.; Hsia, S.-M.; Lee, M.-Y.; Chen, H.-J.; Cheng, F.; Chan, L.-C.; Kuo, Y.-H.; Lin, Y.-L.; Chiang, W. Gastroprotective activities of adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) on the growth of the stomach cancer AGS cell line and indomethacin-induced gastric ulcers. J. Agric. Food Chem. 2011, 59, 6025–6033. [Google Scholar]
- Chung, C.-P.; Hsu, H.-Y.; Huang, D.-W.; Hsu, H.-H.; Lin, J.-T.; Shih, C.-K.; Chiang, W. Ethyl acetate fraction of adlay bran ethanolic extract inhibits oncogene expression and suppresses DMH-induced preneoplastic lesions of the colon in F344 rats through an anti-inflammatory pathway. J. Agric. Food Chem. 2010, 58, 7616–7623. [Google Scholar]
- Choi, Y.; Jeong, H.-S.; Lee, J. Antioxidant activity of methanolic extracts from some grains consumed in Korea. Food Chem. 2007, 103, 130–138. [Google Scholar]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture and U.S. Department of Health and Human Services, Dietary Guidelines for Americans, 7th edGovernment Printing Office: Washington, DC, USA, 2010; pp. 36–38.
- Chinese Nutrition Society, The Dietary Guidelines for Chinese Residents; Tibet People’s Publishing House: Lhasa, Tibet, China, 2007; pp. 51–56.
- Sato, K.; Osawa, T. Dietary supplementation with pearl barley (Adlay, Coix lacryma-jobi L. var Ma-yuen Stapf) extract increases oxidation resistance in the liver of rats ex vivo. Food Sci. Technol. Res. 2010, 16, 613–616. [Google Scholar] [CrossRef]
- Berliner, J.A.; Heinecke, J.W. The role of oxidized lipoproteins in atherogenesis. Free Radic. Biol. Med. 1996, 20, 707–727. [Google Scholar]
- Dean, R.; Fu, S.; Stocker, R.; Davies, M.J. Biochemistry and pathology of radical-mediated protein oxidation. Biochem. J. 1997, 324, 1–18. [Google Scholar]
- Assmann, G.; Gotto, A.M., Jr. HDL cholesterol and protective factors in atherosclerosis. Circulation 2004, 109, II8–II14. [Google Scholar]
- Raederstorff, D.G.; Schlachter, M.F.; Elste, V.; Weber, P. Effect of EGCG on lipid absorption and plasma lipid levels in rats. J. Nutr. Biochem. 2003, 14, 326–332. [Google Scholar] [CrossRef]
- Yazdanparast, R.; Ardestani, A.; Jamshidi, S. Experimental diabetes treated with Achillea santolina: Effect on pancreatic oxidative parameters. J. Ethnopharmacol. 2007, 112, 13–18. [Google Scholar] [CrossRef]
- Mateos, R.; Lecumberri, E.; Ramos, S.; Goya, L.; Bravo, L. Determination of malondialdehyde (MDA) by high-performance liquid chromatography in serum and liver as a biomarker for oxidative stress—Application to a rat model for hypercholesterolemia and evaluation of the effect of diets rich in phenolic antioxidants from fruits. J. Chromatogr. B 2005, 827, 76–82. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Park, S.Y.; Bok, S.H.; Jeon, S.M.; Park, Y.B.; Lee, S.J.; Jeong, T.S.; Choi, M.S. Effect of rutin and tannic acid supplements on cholesterol metabolism in rats. Nutr. Res. 2002, 22, 283–295. [Google Scholar] [CrossRef]
- Singh, P.P.; Chandra, A.; Mahdi, F.; Roy, A.; Sharma, P. Reconvene and reconnect the antioxidant hypothesis in human health and disease. IJCB 2010, 25, 225–243. [Google Scholar]
- Yazdanparast, R.; Bahramikia, S.; Ardestani, A. Nasturtium officinale reduces oxidative stress and enhances antioxidant capacity in hypercholesterolaemic rats. Chem.-Biol. Interact. 2008, 172, 176–184. [Google Scholar] [CrossRef]
- Zou, Y.P.; Lu, Y.H.; Wei, D.Z. Hypocholesterolemic effects of a flavonoid-rich extract of Hypericum perforatum L. in rats fed a cholesterol-rich diet. J. Agric. Food Chem. 2005, 53, 2462–2466. [Google Scholar]
- Han, K.-H.; Matsumoto, A.; Shimada, K.-I.; Sekikawa, M.; Fukushima, M. Effects of anthocyanin-rich purple potato flakes on antioxidant status in F344 rats fed a cholesterol-rich diet. Br. J. Nutr. 2007, 98, 914–921. [Google Scholar]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Oxidants and Antioxidants, Part A; Packer, L., Ed.; Academic Press: San Diego, CA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- He, X.; Liu, D.; Liu, R.H. Sodium borohydride/chloranil-based assay for quantifying total flavonoids. J. Agric. Food Chem. 2008, 56, 9337–9344. [Google Scholar]
- Gorinstein, S.; Leontowicz, M.; Leontowicz, H.; Najman, K.; Namiesnik, J.; Park, Y.-S.; Jung, S.-T.; Kang, S.-G.; Trakhtenberg, S. Supplementation of garlic lowers lipids and increases antioxidant capacity in plasma of rats. Nutr. Res. 2006, 26, 362–368. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovic, I. Superoxide dismutase an enzymic function for erythrocuprein (Hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, L.; Sun, J.; Yi, Q.; Wang, X.; Ju, X. Protective Effect of Polyphenols Extract of Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) on Hypercholesterolemia-Induced Oxidative Stress in Rats. Molecules 2012, 17, 8886-8897. https://doi.org/10.3390/molecules17088886
Wang L, Sun J, Yi Q, Wang X, Ju X. Protective Effect of Polyphenols Extract of Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) on Hypercholesterolemia-Induced Oxidative Stress in Rats. Molecules. 2012; 17(8):8886-8897. https://doi.org/10.3390/molecules17088886
Chicago/Turabian StyleWang, Lifeng, Jing Sun, Qida Yi, Xuefeng Wang, and Xingrong Ju. 2012. "Protective Effect of Polyphenols Extract of Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) on Hypercholesterolemia-Induced Oxidative Stress in Rats" Molecules 17, no. 8: 8886-8897. https://doi.org/10.3390/molecules17088886