Protective Effects of Chlorella-Derived Peptide Against UVC-Induced Cytotoxicity through Inhibition of Caspase-3 Activity and Reduction of the Expression of Phosphorylated FADD and Cleaved PARP-1 in Skin Fibroblasts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
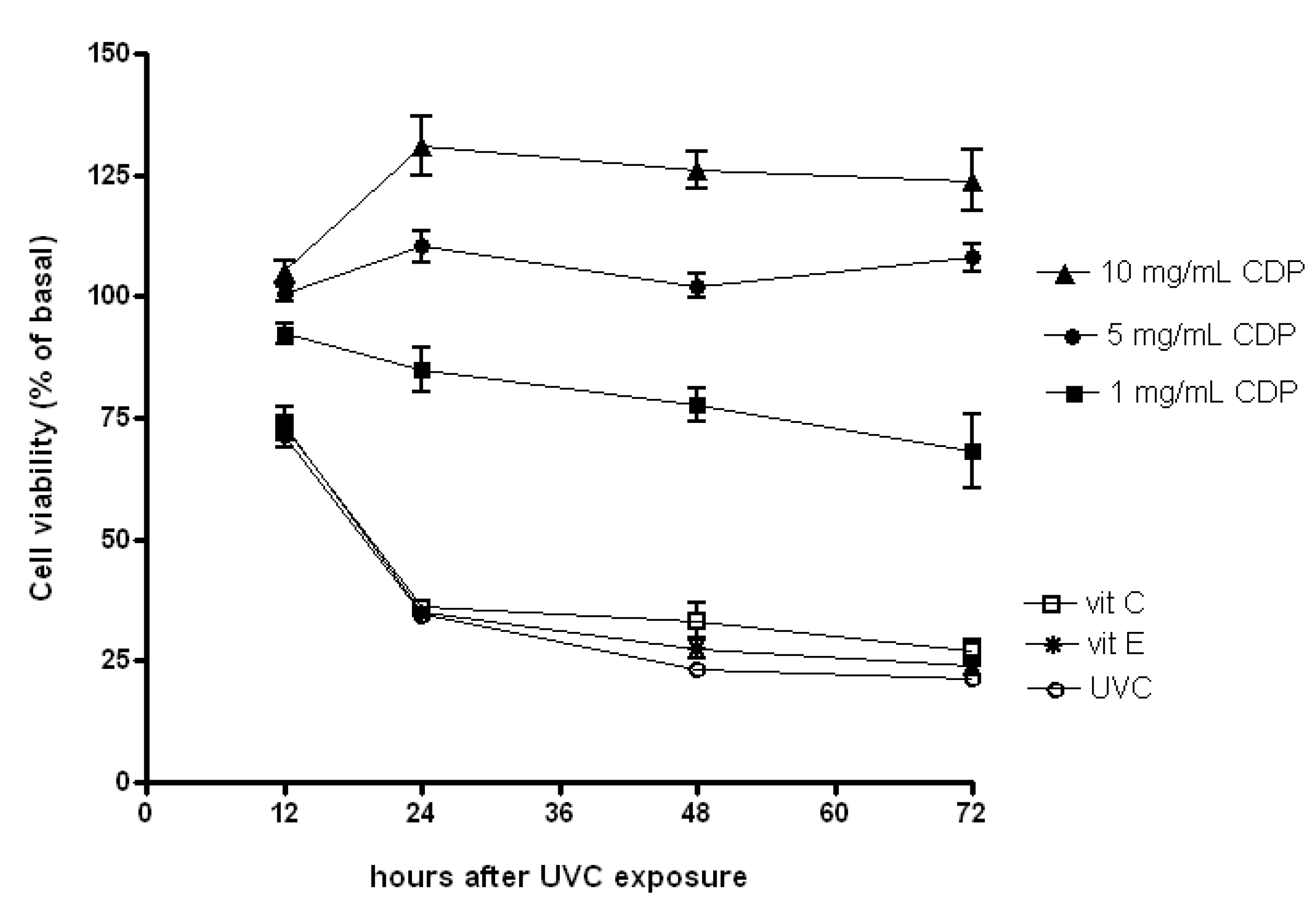
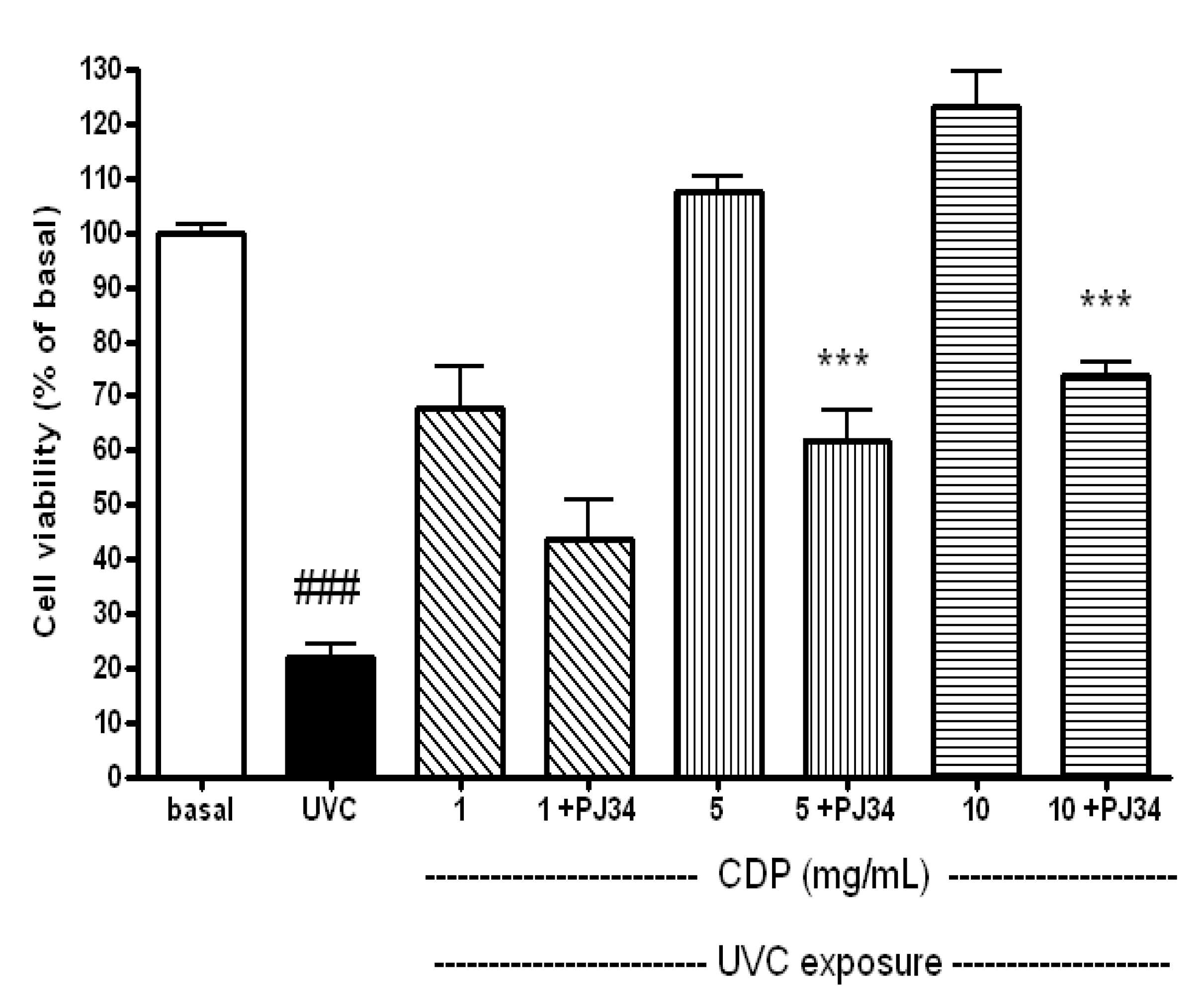
2.1. Protective Effects of CDP Against UVC-Induced Cytotoxicity in Human Skin Fibroblasts

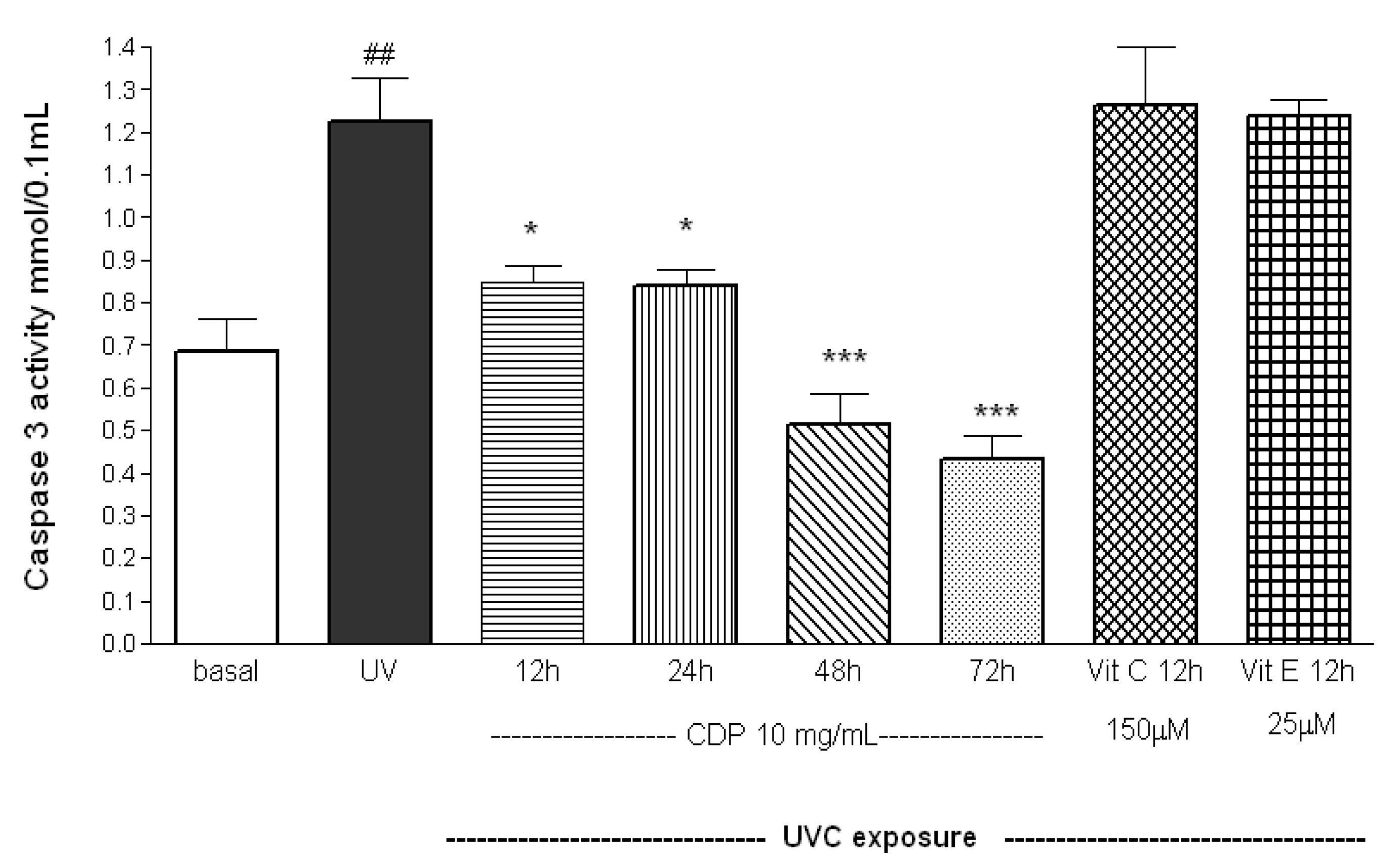
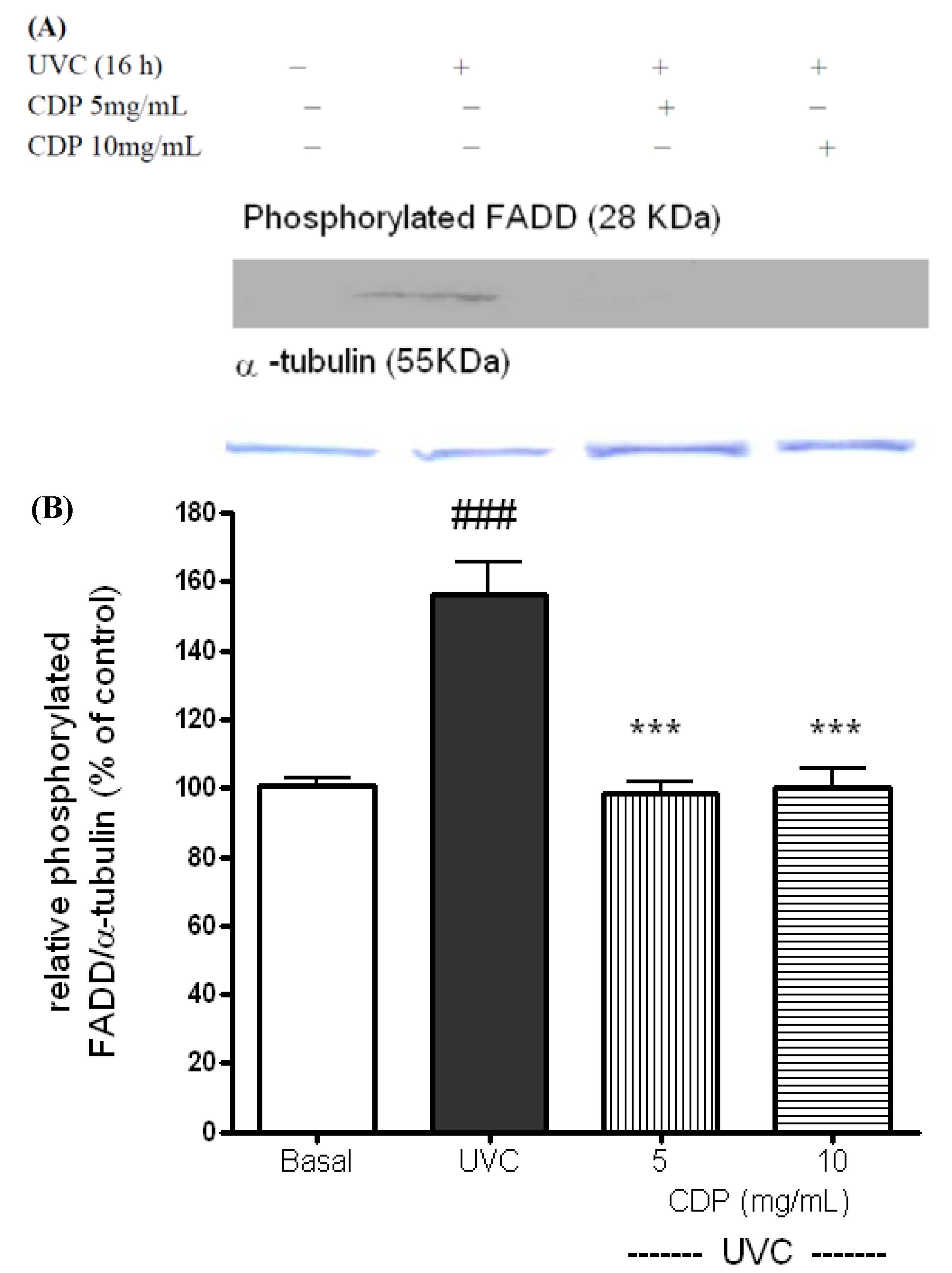
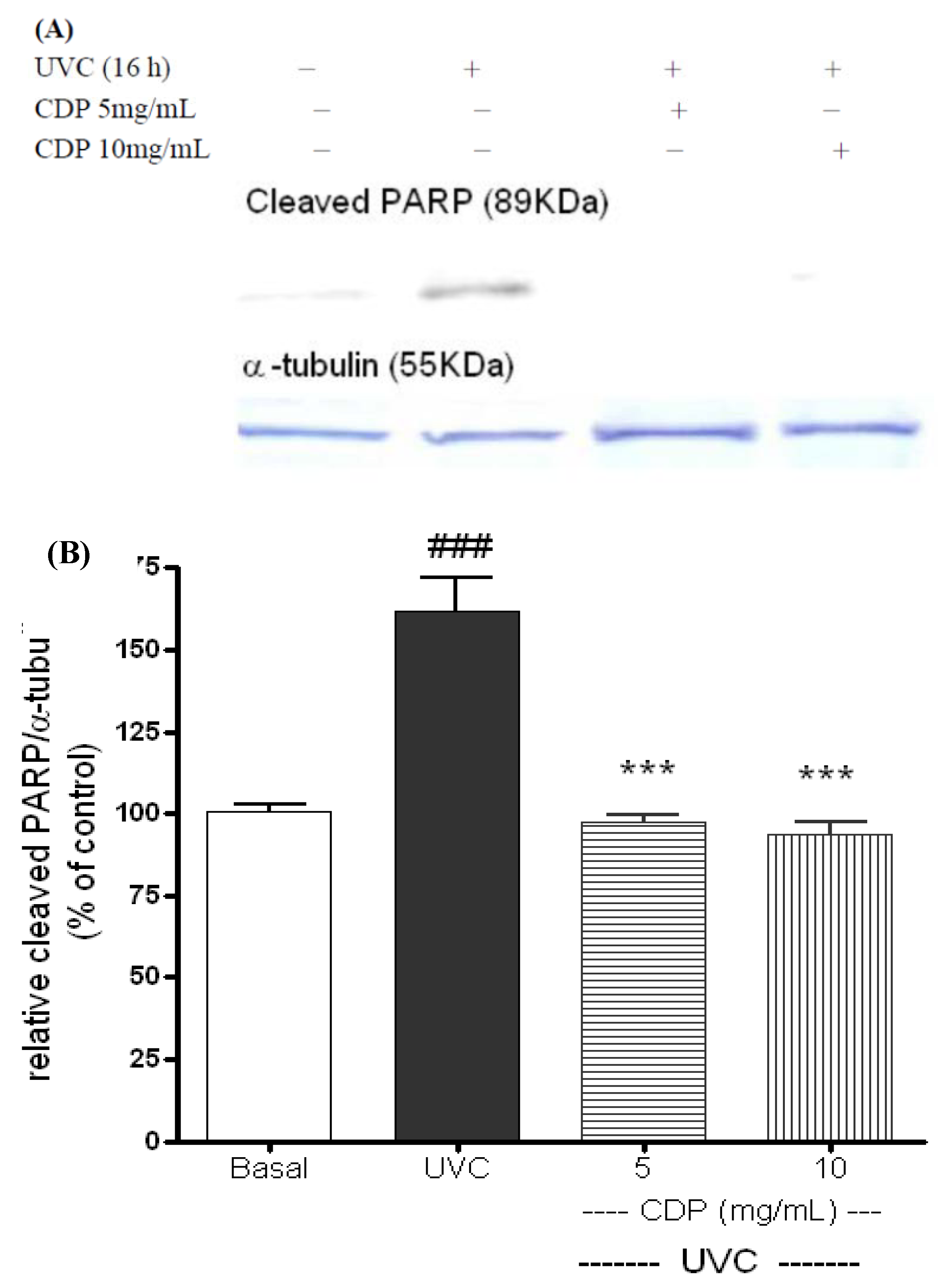
2.2. Inhibitory Effects of CDP on UVC-Induced Apoptosis Measured as Caspase-3 Activity, Phosphorylated FADD, and Cleaved PARP-1 Expression




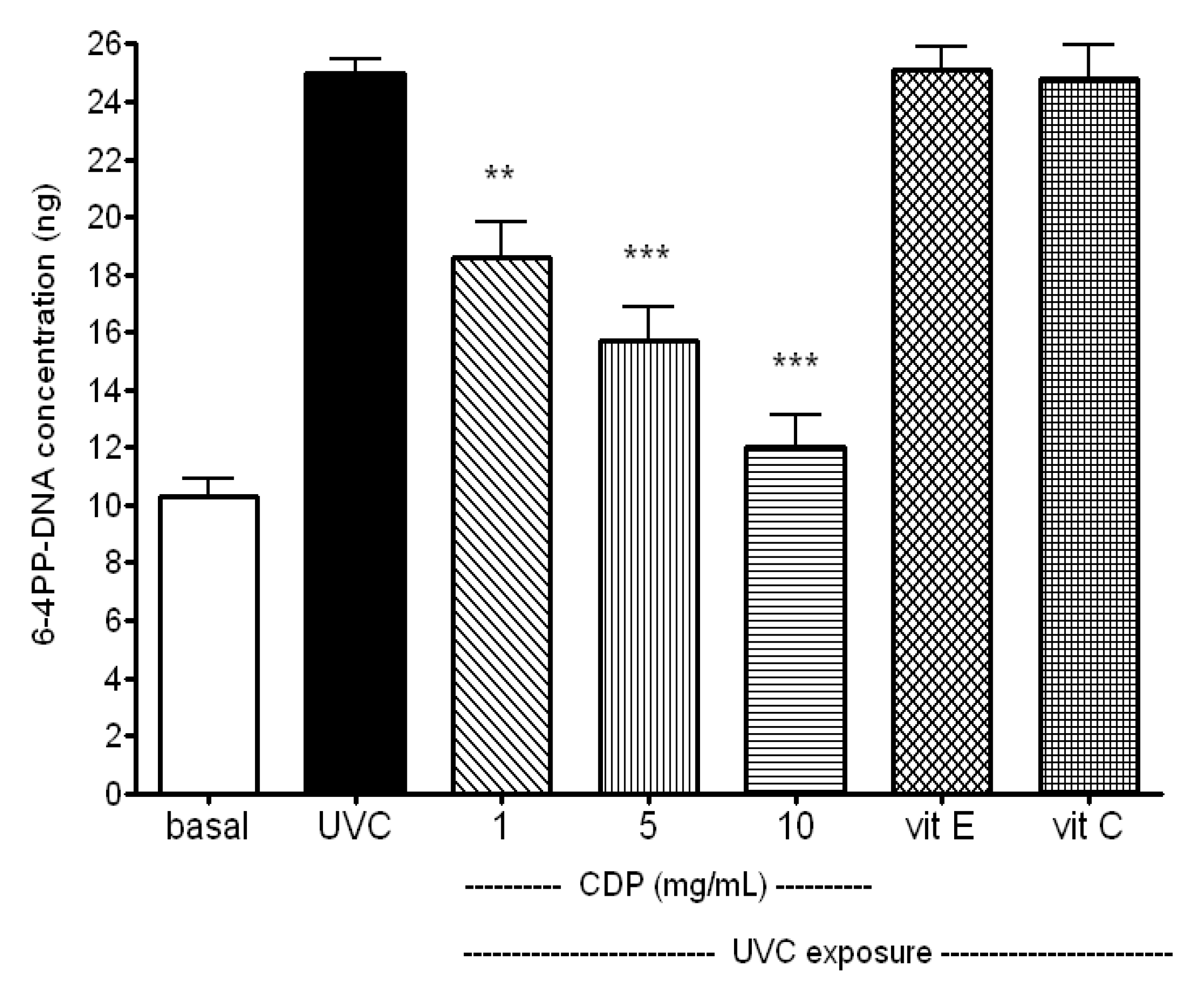
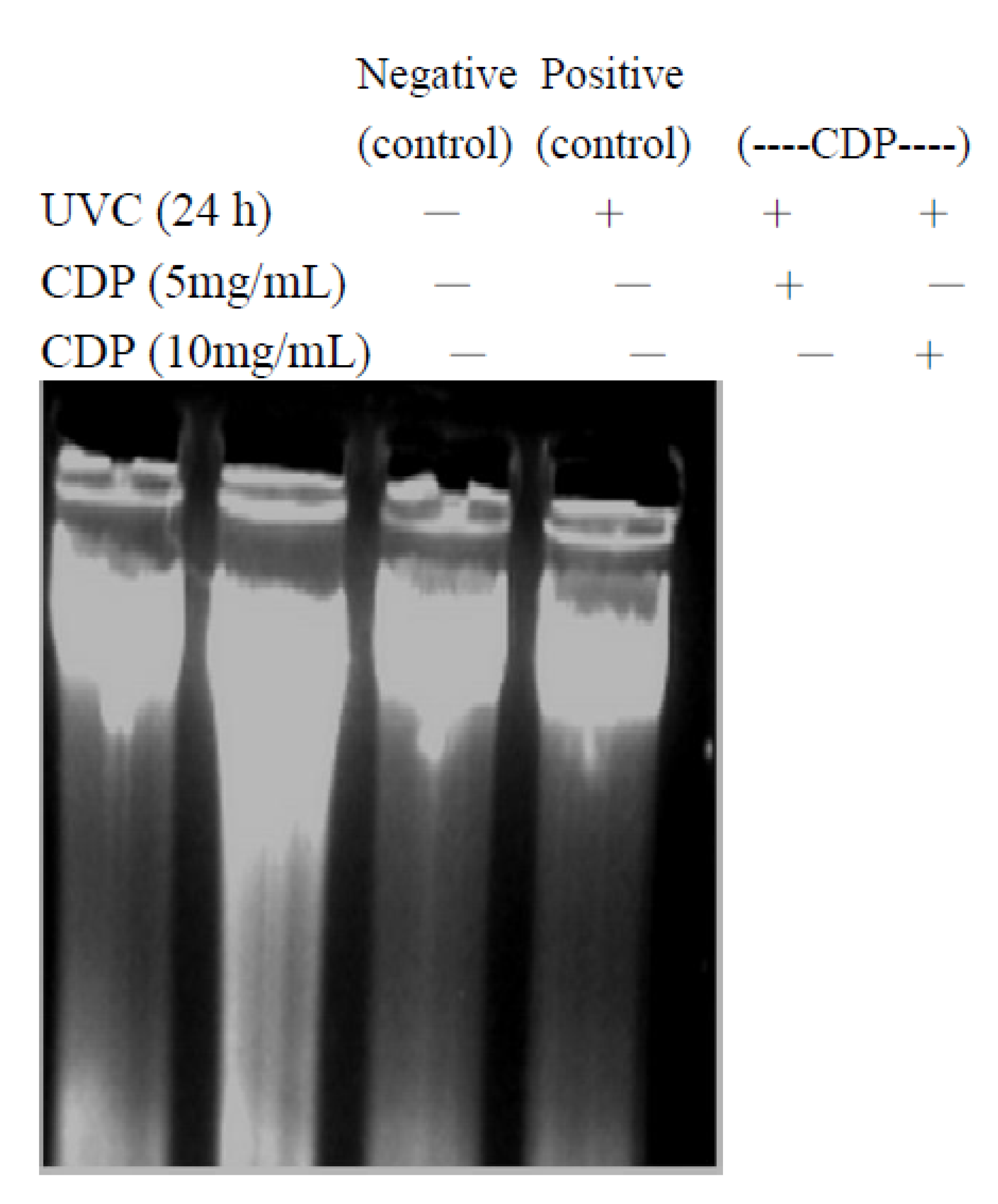
2.3. Inhibitory Effect of CDP on UVC-Induced DNA Damage


3. Experimental
3.1. Materials
3.2. Cell Culture
3.3. Chlorella-Derived Peptide (CDP) Preparation
3.4. Cell Viability Assay
3.5. Caspase-3 Activity Assay
3.6. Western Blotting Analysis of Phosphorylated FADD and Cleavage PARP-1
3.7. DNA Damage Assay
3.8. DNA Fragment Assay
3.9. Statistics
4. Conclusions
Acknowledgments
References
- Ames, B.N.; Gold, L.S.; Willett, W.C. The causes and prevention of cancer. Proc. Natl. Acad. Sci. USA 1995, 92, 5258–5265. [Google Scholar]
- Loft, X.; Poulsen, H.E. Cancer risk and oxidative DNA damage in man. J. Mol. Med. 1996, 74, 297–342. [Google Scholar] [CrossRef]
- Coyle, J.T.; Puttfarcken, P. Oxidative stress, glutamate, and neurodegenerative disorders. Science 1993, 262, 689–695. [Google Scholar]
- Beckman, K.B.; Ames, B.N. Oxidative decay of DNA. Biol. Chem. 1997, 272, 19633–19636. [Google Scholar] [CrossRef]
- Girotti, A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J. Lipid Res. 1998, 39, 1529–1542. [Google Scholar]
- Jiang, W.; Ananthaswamy, H.N.; Muller, H.K.; Kripke, M.L. p53 protects against skin cancer induction by UV-B radiation. Oncogene 1999, 18, 4247–4253. [Google Scholar] [CrossRef]
- Rehemtulla, A.; Hamilton, C.A.; Chinnaiya, A.M.; Dixit, V.M. Ultraviolet radiation-induced apoptosis is mediated by activation of CD-95 (Fas/APO-1) . J. Biol. Chem. 1997, 272, 25783–23586. [Google Scholar]
- Suda, T.; Takahashi, T.; Golstein, P.; Nagata, S. Molecular cloning and expression of the Fas ligand, a novel member of the tumor necrosis factor family. Cell 1993, 75, 1169–1178. [Google Scholar] [CrossRef]
- Los, M.; Herr, I.; Friesen, C.; Fulda, S.; Schulze-Osthoff, K.; Debatin, K.M. Cross-resistance of CD95- and drug-induced apoptosis as a consequence of deficient activation of caspase (ICE/Ced-3 protease). Blood 1997, 90, 3118–3129. [Google Scholar]
- Muzio, M.; Chinnaiyan, A.M.; Kischkel, F.C.; O’Rourke, K.; Shevchenko, A.; Ni, J.; Scaffidi, C.; Bretz, J.D.; Zhang, M.; Gentz, R.; et al. FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to the CD95 (Fas/APO-1) death-inducing signaling complex. Cell 1996, 85, 817–827. [Google Scholar] [CrossRef]
- Ernari, M.; Sakahira, H.; Yokoyama, H.; Okawa, K.; Iwamatsu, A.; Nagata, S. A caspase-activated Dnase that degrades DNA during apoptosis and its inhibitor ICAD. Nature 1998, 391, 43–50. [Google Scholar]
- Oliver, F.J.; de la Rubia, G.; Rolli, V.; Ruiz-Ruiz, M.C.; de Murcia, G.; Murcia, J.M. Importance of poly(ADP-ribose) polymerase and its cleavage in apoptosis. Lesson from an uncleavable mutant. J. Biol. Chem. 1998, 273, 33533–33539. [Google Scholar]
- Ghodgaonkar, M.M.; Zacal, N.; Kassam, S.; Rainbow, A.J.; Shah, G.M. Depletion of poly(ADP-ribose) polymerase-1 reduces host cell reactivation of a UV-damaged adenovirus-encoded reporter gene in human dermal fibroblasts. DNA Repair 2008, 7, 617–632. [Google Scholar] [CrossRef]
- Germain, L.; Lean, A.; Auger, F.; Garrel, D. Human wound healing fibroblasts have greater contractile properties than dermal fibroblasts. J. Surg. Res. 1994, 57, 268–273. [Google Scholar] [CrossRef]
- Fahey, T.; Turbeville, T.; McIntyre, K. Differential TNF secretion by wound fibroblasts compared to normal fibroblast in response to LPS. J. Surg. Res. 1995, 58, 759–764. [Google Scholar] [CrossRef]
- Dunkern, T.R.; Fritz, G.; Kaina, B. Ultraviolet light-induced DNA damage triggers apoptosis in nucleotide excision repair-deficient cells via Bcl-2 decline and caspase-3/-8 activation. Oncogene 2001, 20, 6026–6038. [Google Scholar] [CrossRef]
- Kim, K.W.; Bai, S.C.; Koo, J.-W.; Wang, X.; Kim, S.-K. Effects of dietary chlorella ellipsoidea supplementation on growth, blood characteristics, and whole-body composition in juvenile japanese flounder paralichthys olivaceus. J. World Aquacult. Soc. 2002, 33, 425–431. [Google Scholar] [CrossRef]
- Suárez, E.R.; Kralovec, J.A.; Noseda, M.D.; Ewart, H.S.; Barrow, C.J.; Lumsdena, M.D.; Grindley, T.B. Isolation, characterization and structural determination of a unique type of arabinogalactan from an immunostimulatory extract of Chlorella pyrenoidosa. Carbohydr. Res. 2005, 340, 1489–1498. [Google Scholar] [CrossRef]
- Cherng, J.Y.; Liu, C.C.; Shen, C.R.; Lin, H.H.; Shih, M.F. Beneficial effects of Chlorella-11 peptide on blocking LPS-induced macrophage activation and alleviating thermal injury-induced inflammation in rats. Int. J. Immunopathol. Pharmacol. 2010, 23, 811–820. [Google Scholar]
- Tanaka, K.; Yamada, A.; Nada, K.; Shoyama, Y.; Kubo, C.; Nomoto, K. Oral administration of a unicellular green algae, chlorella vulgaris, prevents stress-induced ulcer. Planta Med. 1997, 63, 465–466. [Google Scholar] [CrossRef]
- Sano, T.; Kumamoto, Y.; Kamiya, N.; Okuda, M.; Tanaka, Y. Effect of lipophilic extract of chlorella vulgaris on alimentary hyperlipidemia in cholesterol-fed rats. Artery 1988, 15, 217–224. [Google Scholar]
- Cherng, J.-Y.; Shih, M.-F. Preventing dyslipidemia by chlorella pyrenoidosa in rats and hamsters after chronic high fat diet treatment. Life Sci. 2005, 76, 3001–3013. [Google Scholar] [CrossRef]
- Cherng, J.-Y.; Shih, M.-F. Potential hypoglycemic effects of chlorella in streptozotocin-induced diabetic mice. Life Sci. 2005, 78, 980–990. [Google Scholar]
- Cherng, J.-Y.; Shih, M.-F. Improving glycogenesis in Streptozocin (STZ) diabetic mice after administration of green algae chlorella. Life Sci. 2006, 78, 1181–1186. [Google Scholar] [CrossRef]
- Shih, M.-F.; Cherng, J.-Y. Potential protective component of fresh grown unicellular green algae (Resilient Factor) in PMA- and UVB-induced MMP1 expression in human skin fibroblasts. Eur. J. Dermatol. 2008, 18, 303–307. [Google Scholar]
- Chen, C.-L.; Liou, S.-F.; Chen, S.-J.; Shih, M.-F. Rotective effects of Chlorella-derived peptide on UVB-induced production of MMP-1 and degradation of procollagen genes in human skin fibroblasts. Regul. Toxicol. Pharm. 2011, 60, 112–119. [Google Scholar] [CrossRef]
- Shih, M.-F.; Cherng, J.-Y. Prevention of short UV wave-induced Caspase 3 activity by water extract of Chlorella in human skin fibroblast. Acta Pharmacol. Sin. 2006, 27, 325. [Google Scholar]
- Allan, L.A.; Fried, M. p53-dependent apoptosis or growth arrest induced by different forms of radiation in U2OS cells: p21WAF1/CIP1 repression in UV induced apoptosis. Oncogene 1999, 18, 5403–5412. [Google Scholar] [CrossRef]
- Latonen, L.; Taya, Y.; Laiho, M. UV-radiation induces dose-dependent regulation of p53 response and modulates p53-HDM2 interaction in human fibroblasts. Oncogene 2001, 20, 6784–6793. [Google Scholar]
- Maki, C.G.; Howley, P.M. Ubiquitination of p53 and p21 is differentially affected by ionizing and UV radiation. Mol. Cell. Biol. 1997, 17, 355–363. [Google Scholar]
- Godar, D.E.; Lucas, A.D. Spectral dependence of UV-induced immediate and delayed apoptosis: the role of membrane and DNA damage. Photochem. Photobiol. 1995, 62, 108–113. [Google Scholar] [CrossRef]
- Li, L.; Sampat, K.; Hu, N.; Zakari, J.; Yuspa, S.H. Protein kinase C negatively regulates Akt activity and modifies UVC-induced apoptosis in mouse keratinocytes. J. Biol. Chem. 2006, 281, 3237–3243. [Google Scholar]
- Cheng, F.C.; Lin, A.; Feng, J.J.; Mizoguchi, T.; Takekoshi, H.; Hitoshi, K.; Kato, Y.; Naoki, Y. Effects of chlorella on activities of protein tyrosine phosphatases, matrix metalloproteinases, caspases, cytokine release, B and T cell proliferations, and phorbol ester receptor binding. J. Med. Food 2004, 27, 146–152. [Google Scholar]
- Ramos, A.L.; Torello, C.O.; Queiroz, M.L. Chlorella vulgaris modulates immunomyelopoietic activity and enhances the resistance of tumor-bearing mice. Nutr. Cancer 2010, 62, 1170–1180. [Google Scholar] [CrossRef]
- Lazebnik, Y.A.; Kaufmann, S.H.; Desnoyers, S.; Poirier, G.G.; Earnshaw, W.C. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 1994, 371, 346–347. [Google Scholar]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shih, M.F.; Cherng, J.Y. Protective Effects of Chlorella-Derived Peptide Against UVC-Induced Cytotoxicity through Inhibition of Caspase-3 Activity and Reduction of the Expression of Phosphorylated FADD and Cleaved PARP-1 in Skin Fibroblasts. Molecules 2012, 17, 9116-9128. https://doi.org/10.3390/molecules17089116
Shih MF, Cherng JY. Protective Effects of Chlorella-Derived Peptide Against UVC-Induced Cytotoxicity through Inhibition of Caspase-3 Activity and Reduction of the Expression of Phosphorylated FADD and Cleaved PARP-1 in Skin Fibroblasts. Molecules. 2012; 17(8):9116-9128. https://doi.org/10.3390/molecules17089116
Chicago/Turabian StyleShih, Mei Fen, and Jong Yuh Cherng. 2012. "Protective Effects of Chlorella-Derived Peptide Against UVC-Induced Cytotoxicity through Inhibition of Caspase-3 Activity and Reduction of the Expression of Phosphorylated FADD and Cleaved PARP-1 in Skin Fibroblasts" Molecules 17, no. 8: 9116-9128. https://doi.org/10.3390/molecules17089116