Influences of Dryopteris crassirhizoma Extract on the Viability, Growth and Virulence Properties of Streptococcus mutans

Abstract
:1. Introduction
2. Results and Discussion
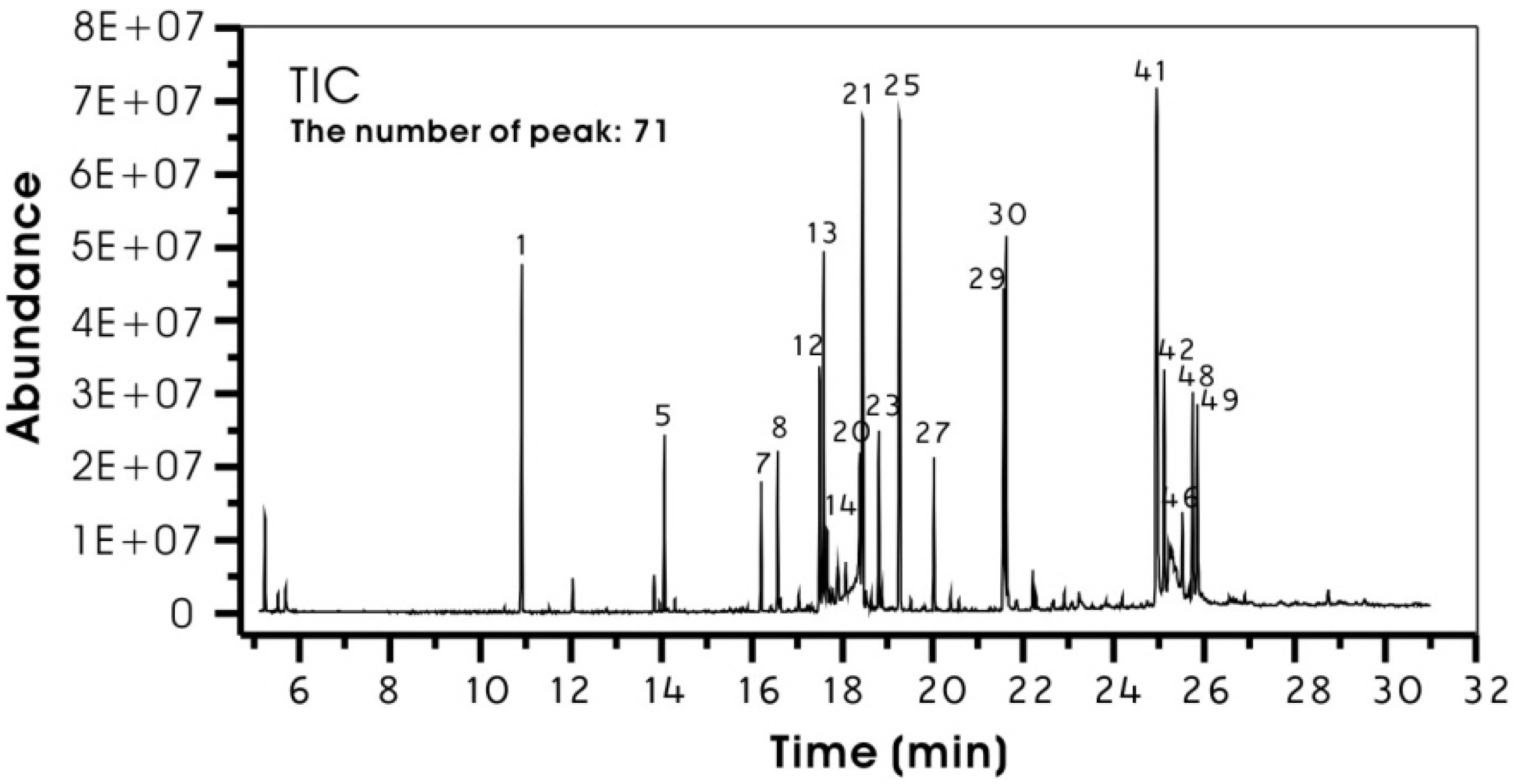
2.1. Phytochemical Composition of the Extract


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. b | Compounds | Rt c | RAP d (%) | Peak No. b | Compounds | Rt c | RAP d (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 | Glycerol | 10.91 | 4.57 | 26 | Gluconic acid | 19.52 | 0.16 | ||
| 2 | Butenedioic acid | 12.04 | 0.38 | 27 | Palmitic acid | 20.03 | 1.81 | ||
| 3 | Malic acid | 13.84 | 0.42 | 28 | Inositol | 20.40 | 0.26 | ||
| 4 | Butanal | 13.97 | 0.13 | 29 | Linoleic acid | 21.56 | 4.67 | ||
| 5 | Threitol | 14.07 | 2.01 | 30 | Oleic acid | 21.62 | 5.27 | ||
| 6 | Proline | 14.30 | 0.17 | 31 | Stearic acid | 21.67 | 0.33 | ||
| 7 | U.I | 16.21 | 1.53 | 32 | U.I | 21.86 | 0.14 | ||
| 8 | Xylitol | 16.58 | 1.97 | 33 | U.I | 22.21 | 0.53 | ||
| 9 | Ribitol | 16.63 | 0.17 | 34 | U.1 | 22.28 | 0.20 | ||
| 10 | Xylonic acid | 17.04 | 0.29 | 35 | U.I | 22.67 | 0.12 | ||
| 11 | Propanoic acid | 17.24 | 0.10 | 36 | Arachidonic acid | 22.91 | 0.25 | ||
| 12 | Fructose | 17.50 | 3.14 | 38 | U.I | 23.08 | 0.21 | ||
| 13 | Fructose | 17.59 | 4.83 | 39 | U.I | 23.24 | 0.56 | ||
| 14 | Citric acid | 17.67 | 1.62 | 40 | U.I | 23.83 | 0.19 | ||
| 15 | Benzoic acid | 17.72 | 0.30 | 41 | Sucrose | 24.95 | 11.84 | ||
| 16 | U.I | 17.79 | 0.54 | 42 | Lactulose | 25.12 | 3.77 | ||
| 17 | U.I | 17.89 | 0.76 | 43 | U.I | 25.22 | 1.42 | ||
| 18 | U.I | 17.92 | 0.59 | 44 | U.I | 25.28 | 1.93 | ||
| 19 | U.I | 18.08 | 1.55 | 45 | U.I | 25.35 | 0.60 | ||
| 20 | U.I | 18.39 | 5.51 | 46 | Maltose | 25.52 | 1.69 | ||
| 21 | Glucose | 18.45 | 9.28 | 47 | U.I | 25.66 | 0.31 | ||
| 22 | Galactose | 18.53 | 0.46 | 48 | U.I | 25.75 | 3.05 | ||
| 23 | Mannitol | 18.81 | 2.26 | 49 | U.I | 25.85 | 3.03 | ||
| 24 | Glucitol | 18.88 | 0.37 | 50 | U.I | 25.94 | 0.20 | ||
| 25 | Glucose | 19.26 | 9.86 | Total RAP of identified compounds: | 73.91 | ||||
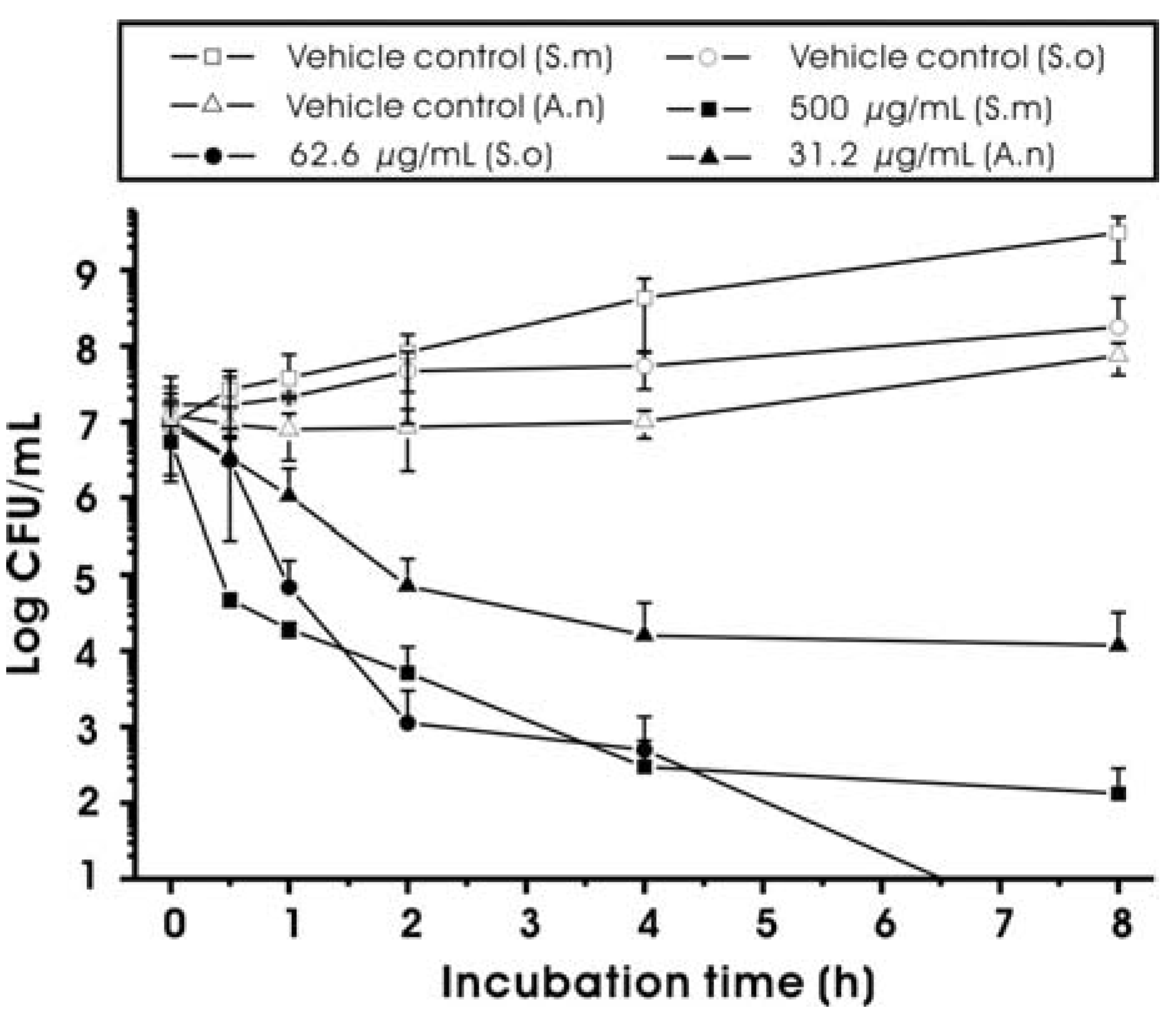
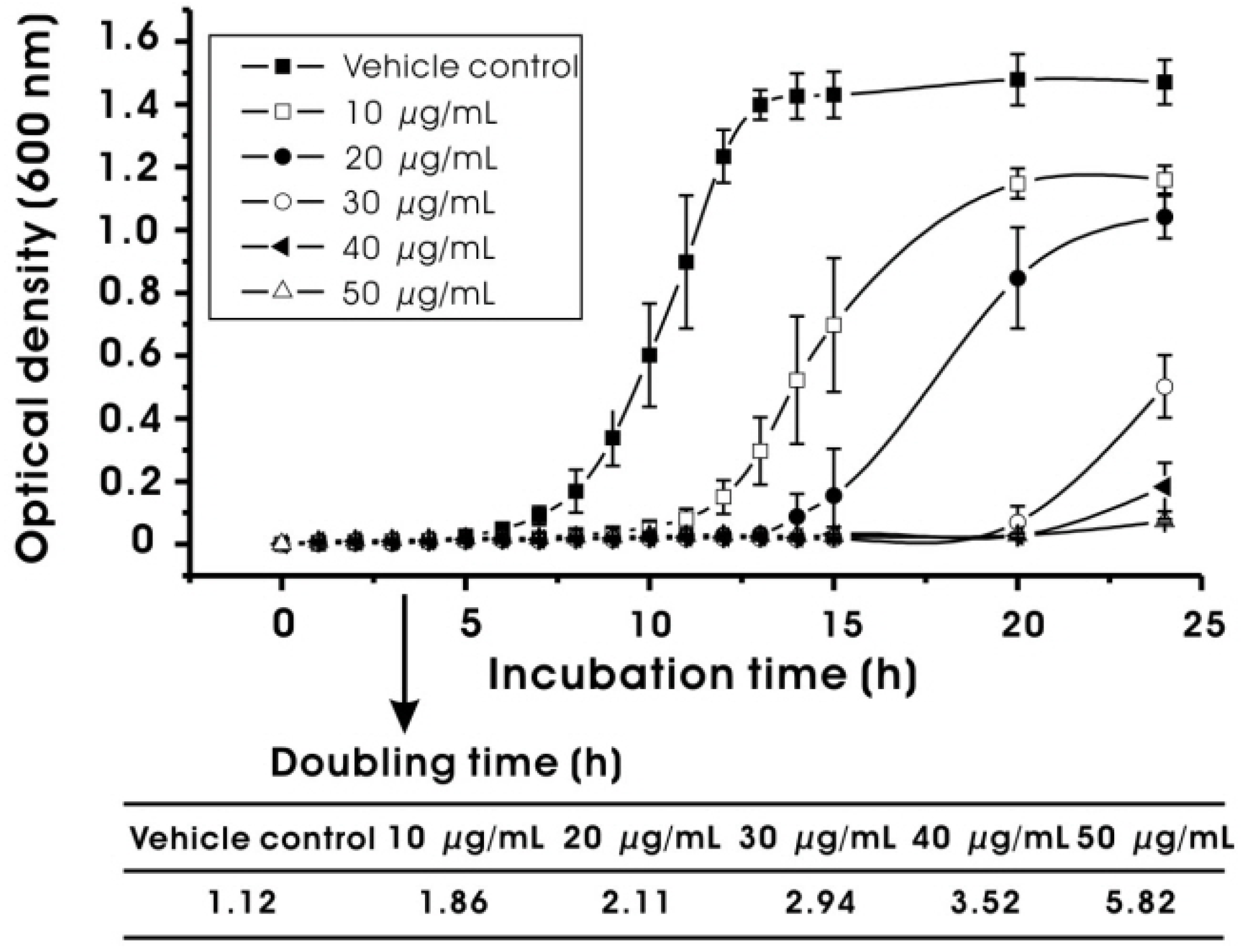
2.2. Effect on Bacterial Viability and Growth
| Test bacteria | MIC (μg/mL) | MBC (μg/mL) |
|---|---|---|
| Streptococcus mutans UA159 | 62.5 | 250 |
| Streptococcus oralis 35037 | 15.6 | 31.3 |
| Actinomyces naeslundii 12104 | 7.8 | 15.6 |


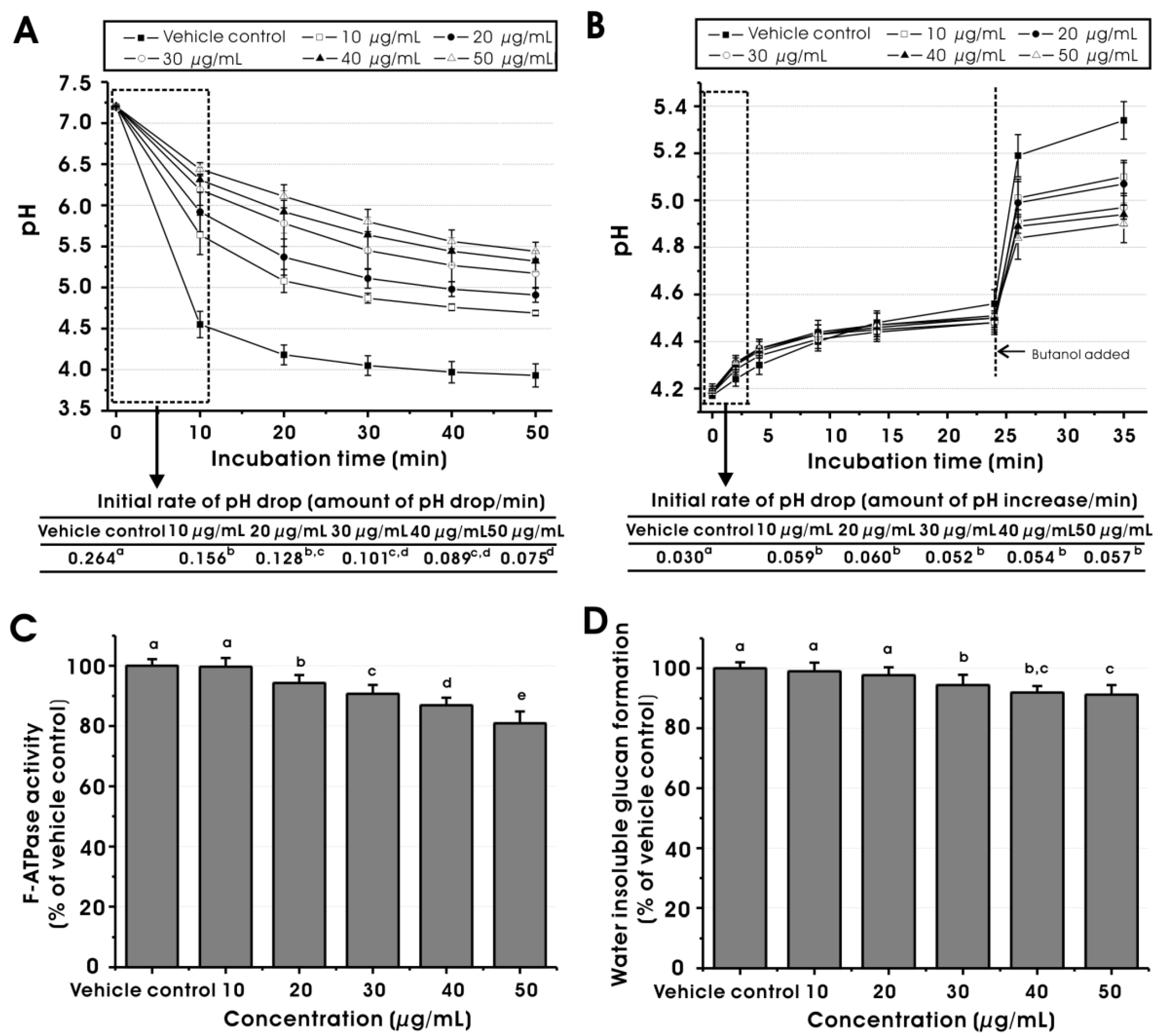
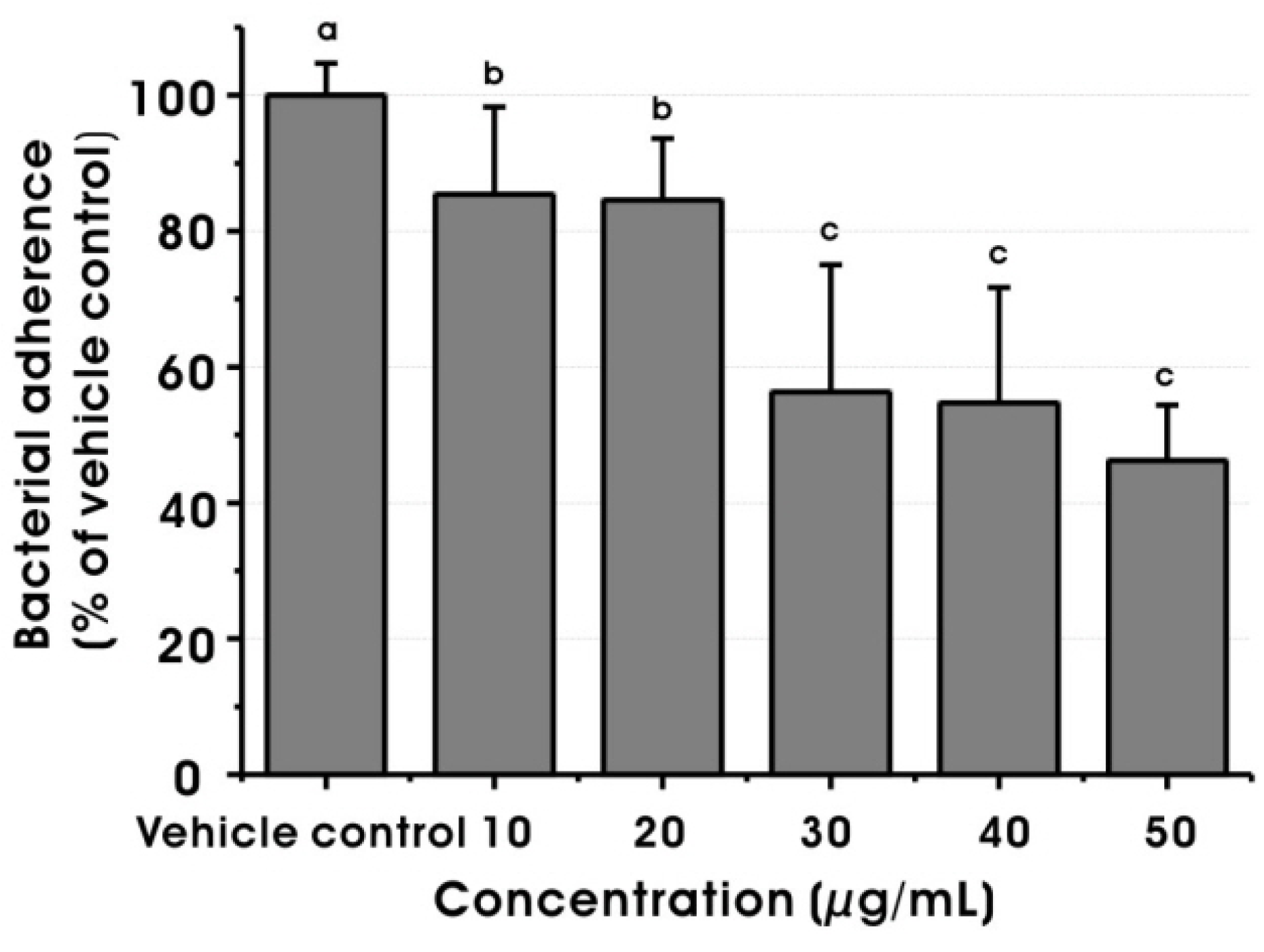
2.3. Effect on Virulence Properties of S. mutans at Sub-MIC Levels


3. Experimental
3.1. Bacterial Strains and Culture Medium
3.2. Plant Extraction and Chemical Characterization by GC-MS
3.3. Effect on Bacterial Viability and Growth
3.3.1. Determination of MIC and MBC
3.3.2. Time-kill Assay
3.3.3. Bacterial Growth Rate at Sub-MIC Levels
3.4. Effect on Virulence Properties of S. Mutans at Sub-MIC Levels
3.4.1. Glycolytic pH-drop Assay
3.4.2. Proton Permeability Assay
3.4.3. F-ATPase Activity Assay
3.4.4. GTF Preparation and GTF Activity Assay
3.4.5. Sucrose-dependent Adherence Assay
3.5. Statistical Analyses
4. Conclusions
Acknowledgments
References
- Marsh, P.D. Are dental diseases examples of ecological catastrophes? Microbiology 2003, 149, 279–294. [Google Scholar] [CrossRef]
- Quivey, R.G., Jr.; Kuhnert, W.L.; Hahn, K. Adaptation of oral streptococci to low pH. Adv. Microb. Physiol. 2000, 42, 239–274. [Google Scholar] [CrossRef]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar]
- Jeon, J.G.; Rosalen, P.L.; Falsetta, M.L.; Koo, H. Natural products in caries research: Current (limited) knowledge, challenges and future perspective. Caries Res. 2011, 45, 243–263. [Google Scholar] [CrossRef]
- Lee, H.B.; Kim, J.C.; Lee, S.M. Antibacterial activity of two phloroglucinols, flavaspidic acidsAB and PB, from Dryopteris crassirhizoma. Arch. Pharm.Res. 2009, 32, 655–659. [Google Scholar] [CrossRef]
- Chang, S.H.; Bae, J.H.; Hong, D.P.; Choi, K.D.; Kim, S.C.; Her, E.; Kim, S.H.; Kang, C.D. Dryopteris crassirhizoma has anti-cancer effects through both extrinsic and intrinsic apoptotic pathways and G0/G1 phase arrest in human prostate cancer cells. J. Ethnopharmacol. 2010, 130, 248–254. [Google Scholar] [CrossRef]
- Namba, T.; Tsunezuka, M.; Hattori, M. Dental caries prevention by traditional Chinese medicines. Planta Med. 1982, 44, 100–106. [Google Scholar] [CrossRef]
- Gao, Z.; Ali, Z.; Zhao, J.; Qiao, L.; Lei, H.; Lu, Y.; Khan, I.A. Phytochemical investigation of the rhizomes of Dryopteris crassirhizoma. Phytochem. Lett. 2008, 1, 188–190. [Google Scholar] [CrossRef]
- Almeida, L.S.; Murata, R.M.; Yatsuda, R.; Santos, M.H.; Nagem, T.J.; Alencar, S.M.; Koo, H.; Rosalen, P.L. Antimicrobial activity of Rheedia brasiliensis and 7-epiclusianone against Streptococcus mutans. Phytomedicine 2008, 15, 886–891. [Google Scholar] [CrossRef]
- Khan, R.; Islam, B.; Akram, M.; Shakil, S.; Ahmad, A.; Ali, S.M.; Siddiqui, M.; Khan, A.U. Antimicrobialactivity of fiveherbalextractsagainstmultidrugresistant (MDR) strains of bacteria and fungus of clinical origin. Molecules 2009, 14, 586–597. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Amato, I.; Ingenito, A.; Zarrelli, A.; Pinto, G.; Pollio, A. Plant polyphenols and their anti-cariogenicproperties: A review. Molecules 2011, 16, 1486–1507. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Simionato, M.R.; Mayer, M.P.; Novo, N.F.; Zelante, F. Effects of subinhibitory concentrations of chemical agents on hydrophobicity and in vitro adherence of Streptococcus mutans and Streptococcus sanguis. Caries Res. 1994, 28, 335–341. [Google Scholar] [CrossRef]
- Bachrach, G.; Jamil, A.; Naor, R.; Tal, G.; Ludmer, Z.; Steinberg, D. Garlic allicin as a potential agent for controlling oral pathogens. J. Med. Food 2011, 14, 1338–1343. [Google Scholar] [CrossRef]
- Halket, J.M.; Waterman, D.; Przyborowska, A.M.; Patel, R.K.; Fraser, P.D.; Bramley, P.M. Chemical derivatization and mass spectral libraries in metabolic profiling by GC/MS and LC/MS/MS. J. Exp. Bot. 2005, 56, 219–243. [Google Scholar]
- Kanani, H.H.; Klapa, M.I. Data correction strategy for metabolomics analysis using gaschromatography-mass spectrometry. Metab. Eng. 2007, 9, 39–51. [Google Scholar] [CrossRef]
- Tsugawa, H.; Tsujimoto, Y.; Arita, M.; Bamba, T.; Fukusaki, E. GC/MS based metabolomics: Development of a data mining system for metabolite identification by using soft independent modeling of class analogy (SIMCA). BMC Bioinformatics 2011, 12, 131. [Google Scholar] [CrossRef]
- Marsh, P.D. Microbiological aspects of the chemical control of plaque and gingivitis. J. Dent. Res. 1992, 71, 1431–1438. [Google Scholar] [CrossRef]
- Rios, J.L.; Recio, M.C.; Villar, A. Screening methods for natural products with antimicrobial activity: A review of the literature. J. Ethnopharmacol. 1988, 23, 127–149. [Google Scholar] [CrossRef]
- Tsai, T.H.; Tsai, T.H.; Chien, Y.C.; Chien, Y.C.; Lee, C.W.; Tsai, P.J. In vitro antimicrobial activities against cariogenic streptococci and their antioxidant capacities: A comparative study of green tea versus different herbs. Food Chem. 2008, 110, 859–864. [Google Scholar] [CrossRef]
- Kuramitsu, H.K. Virulence factors of mutans streptococci: Role of molecular genetics. Crit. Rev. Oral Biol. Med. 1993, 4, 159–176. [Google Scholar]
- Gregoire, S.; Singh, A.P.; Vorsa, N.; Koo, H. Influence of cranberry phenolics on glucan synthesis by glucosyltransferases and Streptococcus mutans acidogenicity. J. Appl. Microbiol. 2007, 103, 1960–1968. [Google Scholar] [CrossRef]
- Bender, G.R.; Sutton, S.V.; Marquis, R.E. Acid tolerance, proton permeabilities, and membrane ATPases of oral streptococci. Infect. Immun. 1986, 53, 331–338. [Google Scholar]
- Schilling, K.M.; Bowen, W.H. Glucans synthesized in situ in experimental salivary pellicle function as specific binding sites for Streptococcus mutans. Infect. Immun. 1992, 60, 284–295. [Google Scholar]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, Mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Osawa, K.; Miyazaki, K.; Shimura, S.; Okuda, J.; Matsumoto, M.; Ooshima, T. Identification of cariostatic substances in the cacao bean husk: Their anti-glucosyltransferase and antibacterial activities. J.Dent.Res. 2001, 80, 2000–2004. [Google Scholar] [CrossRef]
- Hamel, F.G. Preliminary report: Inhibition of cellular proteasome activity by free fatty acids. Metabolism 2009, 58, 1047–1049. [Google Scholar] [CrossRef]
- McLafferty, F.W. Wiley Registry™ of Mass Spectral Data, 7th ed; John Wiley & Sons, Ltd.: New York, NY, USA, 2005. [Google Scholar]
- Song, J.H.; Kim, S.K.; Chang, K.W.; Han, S.K.; Yi, H.K.; Jeon, J.G. In vitro inhibitory effects of Polygonum cuspidatum on bacterial viability and virulence factors of Streptococcus mutans and Streptococcus sobrinus. Arch. Oral Biol. 2006, 51, 1131–1140. [Google Scholar] [CrossRef]
- Khalichi, P.; Cvitkovitch, D.G.; Santerre, J.P. Effect of composite resin biodegradation products on oral streptococcal growth. Biomaterials 2004, 25, 5467–5472. [Google Scholar] [CrossRef]
- Belli, W.A.; Buckley, H.D.; Marquis, R.E. Weak acid effects and fluoride inhibition of glycolysis by Streptococcus mutans GS-5. Can. J. Microbiol. 1995, 41, 785–791. [Google Scholar] [CrossRef]
- Phan, T.N.; Buckner, T.; Sheng, J.; Baldeck, J.D.; Marquis, R.E. hysiologic actions of zinc related to inhibition of acid and alkali production by oral streptococci in suspensions and biofilms. Oral Microbiol. Immunol. 2004, 19, 31–38. [Google Scholar] [CrossRef]
- Bencini, D.A.; Shanley, M.S.; Wild, J.R.; O’Donovan, G.A. New assay for enzymatic phosphate release: Application to aspartate transcarbamylase and other enzymes. Anal. Biochem. 1983, 132, 259–264. [Google Scholar]
- Pandit, S.; Kim, H.J.; Kim, J.E.; Jeon, J.G. Separation of an effective fraction from turmeric against Streptococcus mutans biofilms by the comparison of curcuminoid content and anti-acidogenic activity. Food Chem. 2011, 126, 1565–1570. [Google Scholar] [CrossRef]
- Sample Availability: Samples are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ban, S.-H.; Kim, J.-E.; Pandit, S.; Jeon, J.-G. Influences of Dryopteris crassirhizoma Extract on the Viability, Growth and Virulence Properties of Streptococcus mutans. Molecules 2012, 17, 9231-9244. https://doi.org/10.3390/molecules17089231
Ban S-H, Kim J-E, Pandit S, Jeon J-G. Influences of Dryopteris crassirhizoma Extract on the Viability, Growth and Virulence Properties of Streptococcus mutans. Molecules. 2012; 17(8):9231-9244. https://doi.org/10.3390/molecules17089231
Chicago/Turabian StyleBan, Suk-Ho, Jeong-Eun Kim, Santosh Pandit, and Jae-Gyu Jeon. 2012. "Influences of Dryopteris crassirhizoma Extract on the Viability, Growth and Virulence Properties of Streptococcus mutans" Molecules 17, no. 8: 9231-9244. https://doi.org/10.3390/molecules17089231