Induction of Apoptosis by Ethanolic Extract of Corchorus olitorius Leaf in Human Hepatocellular Carcinoma (HepG2) Cells via a Mitochondria-Dependent Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cytotoxicity of ECO on HepG2 and FL83B Cells

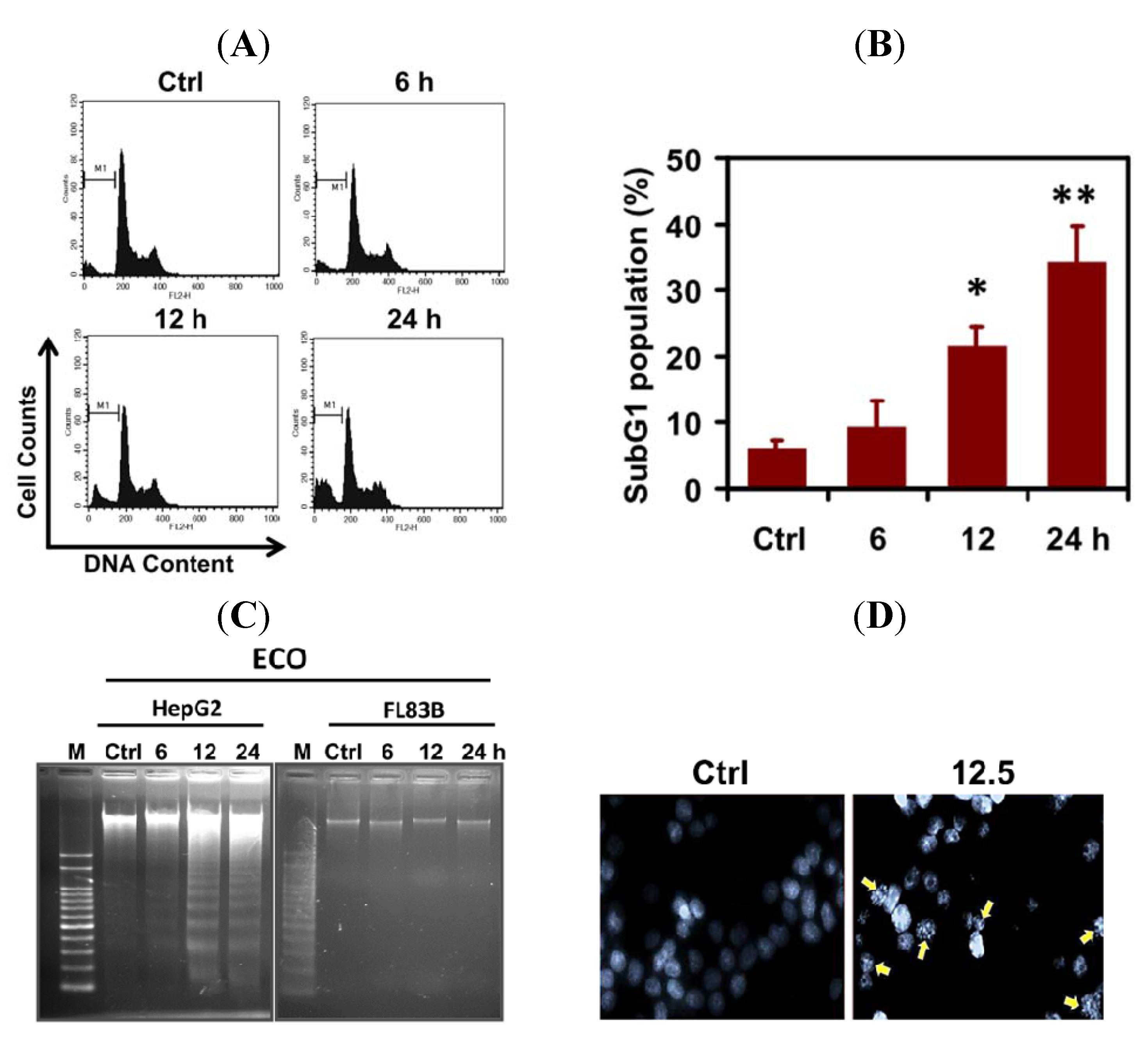
2.2. ECO-Induced Apoptosis in HepG2 Cells

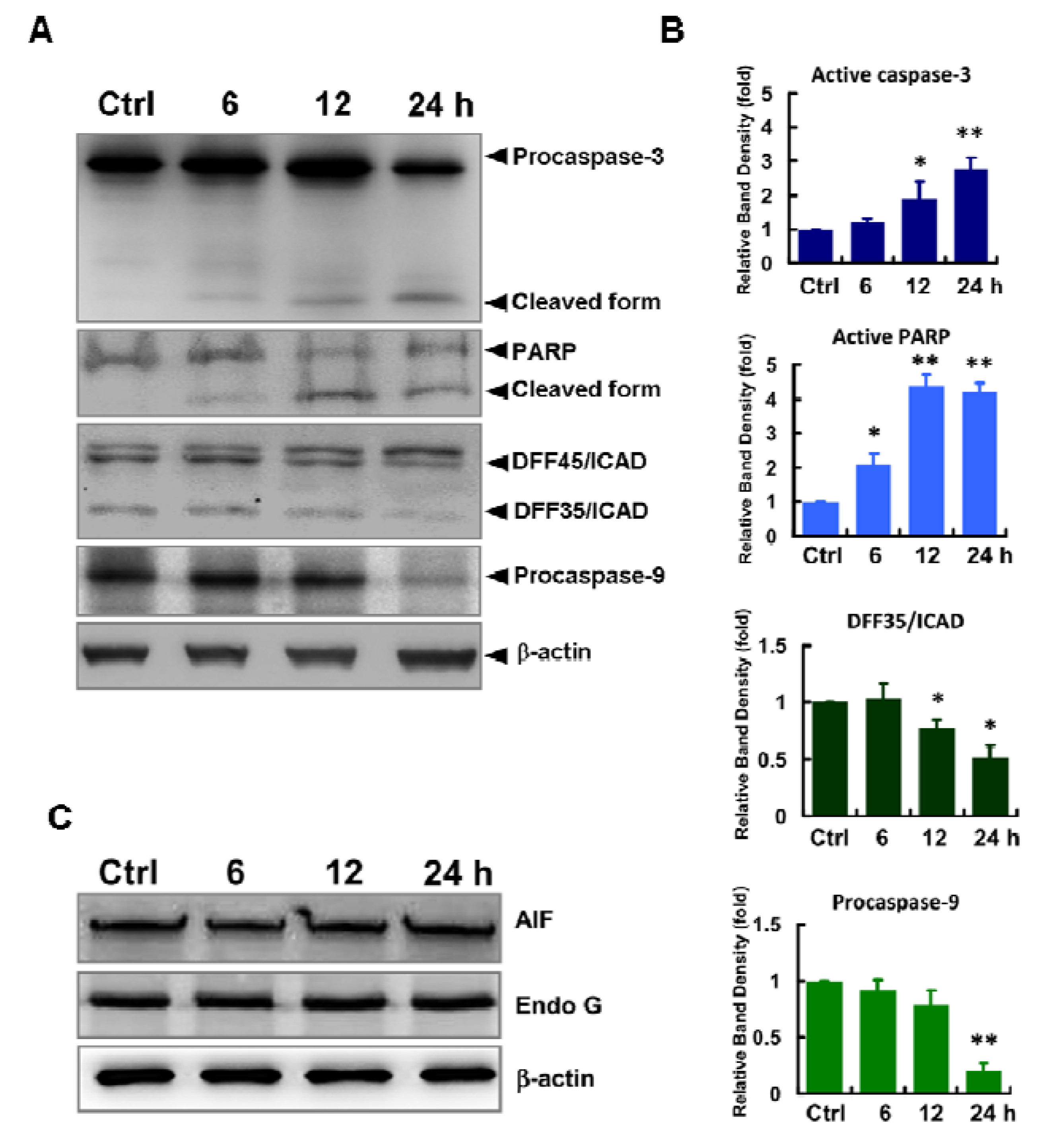
2.3. Caspases-Mediated Apoptosis in ECO-Treated HepG2 Cells
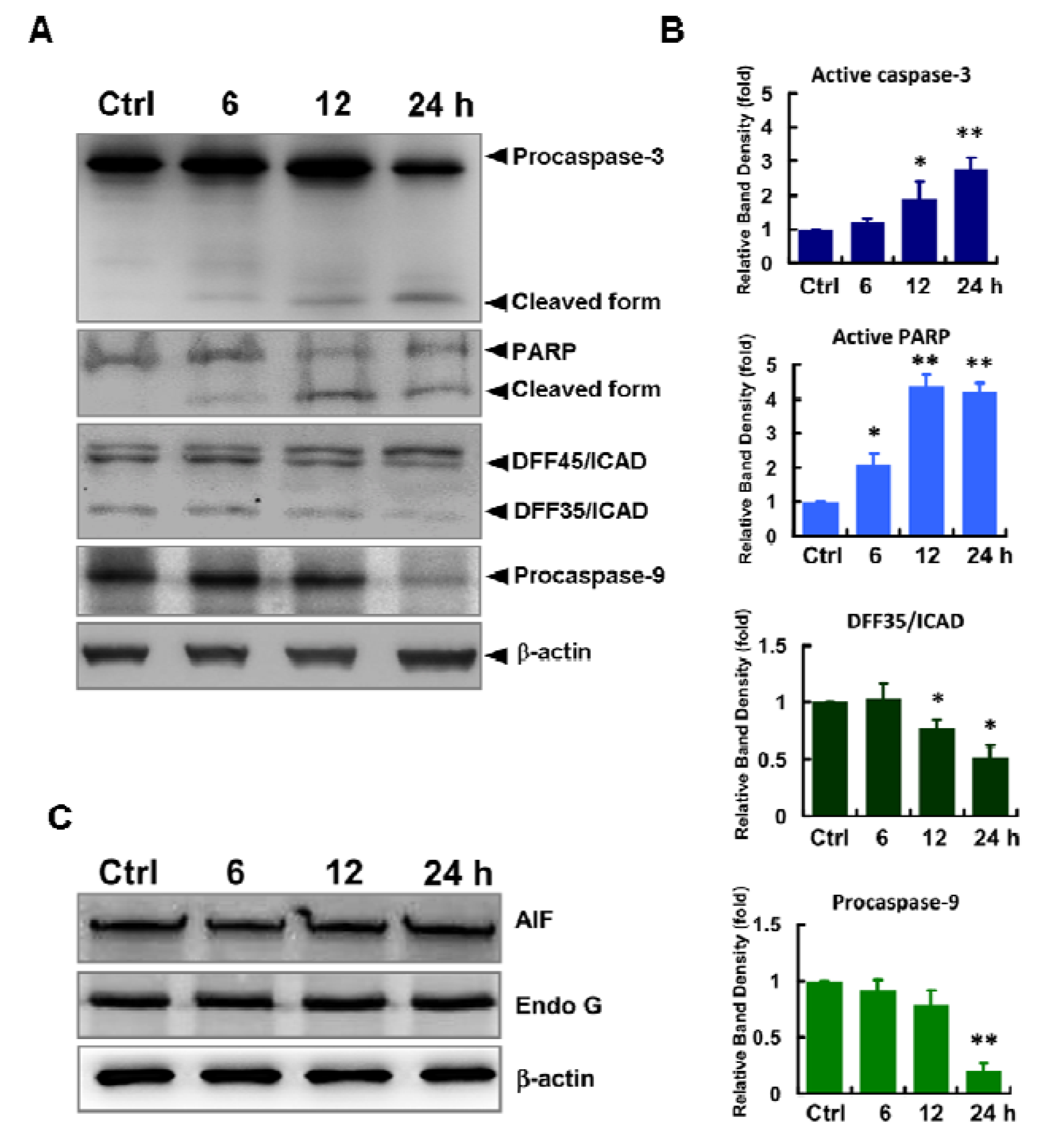
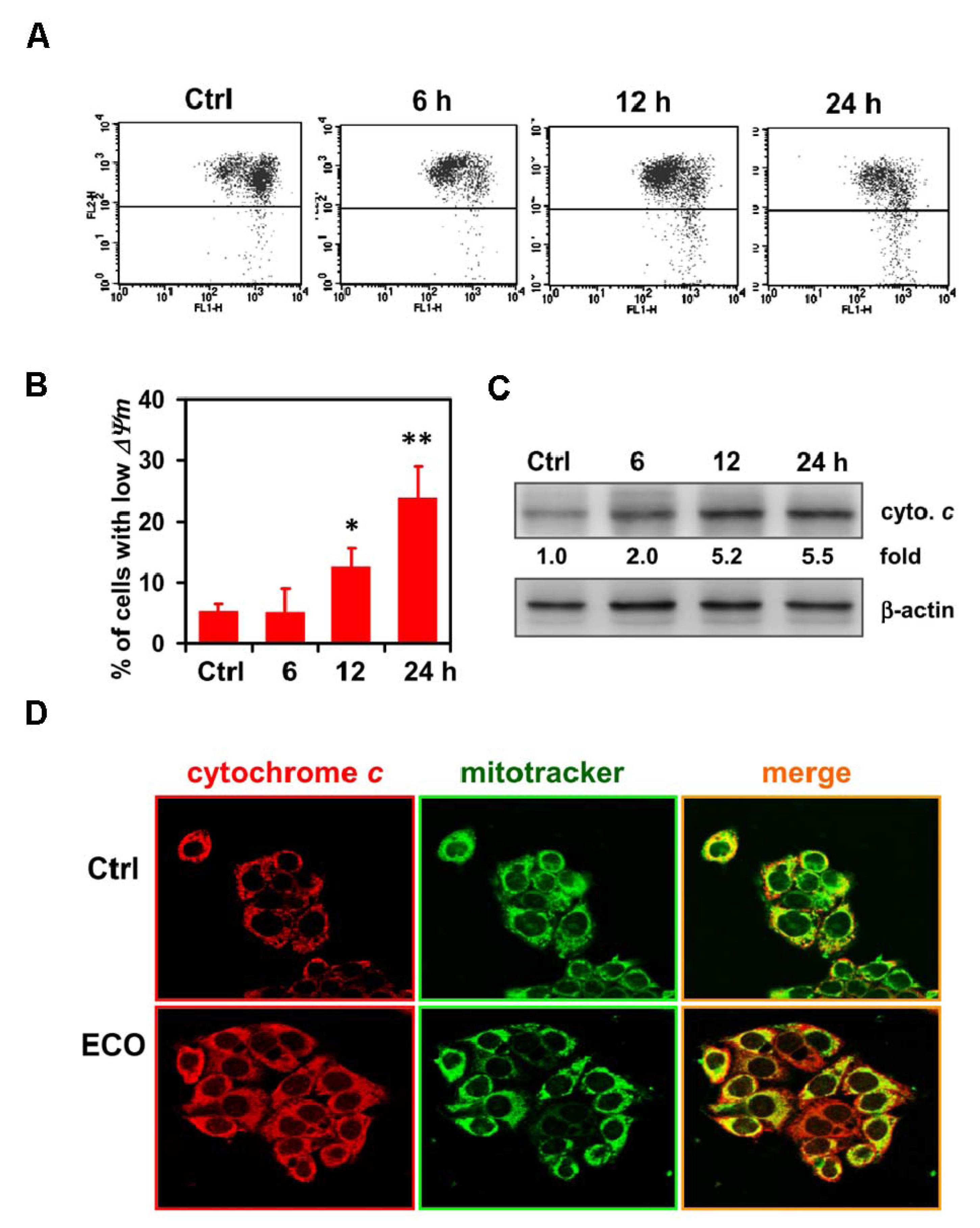
2.4. Mitochondria-Mediated Cell Death in ECO-Treated HepG2 Cells
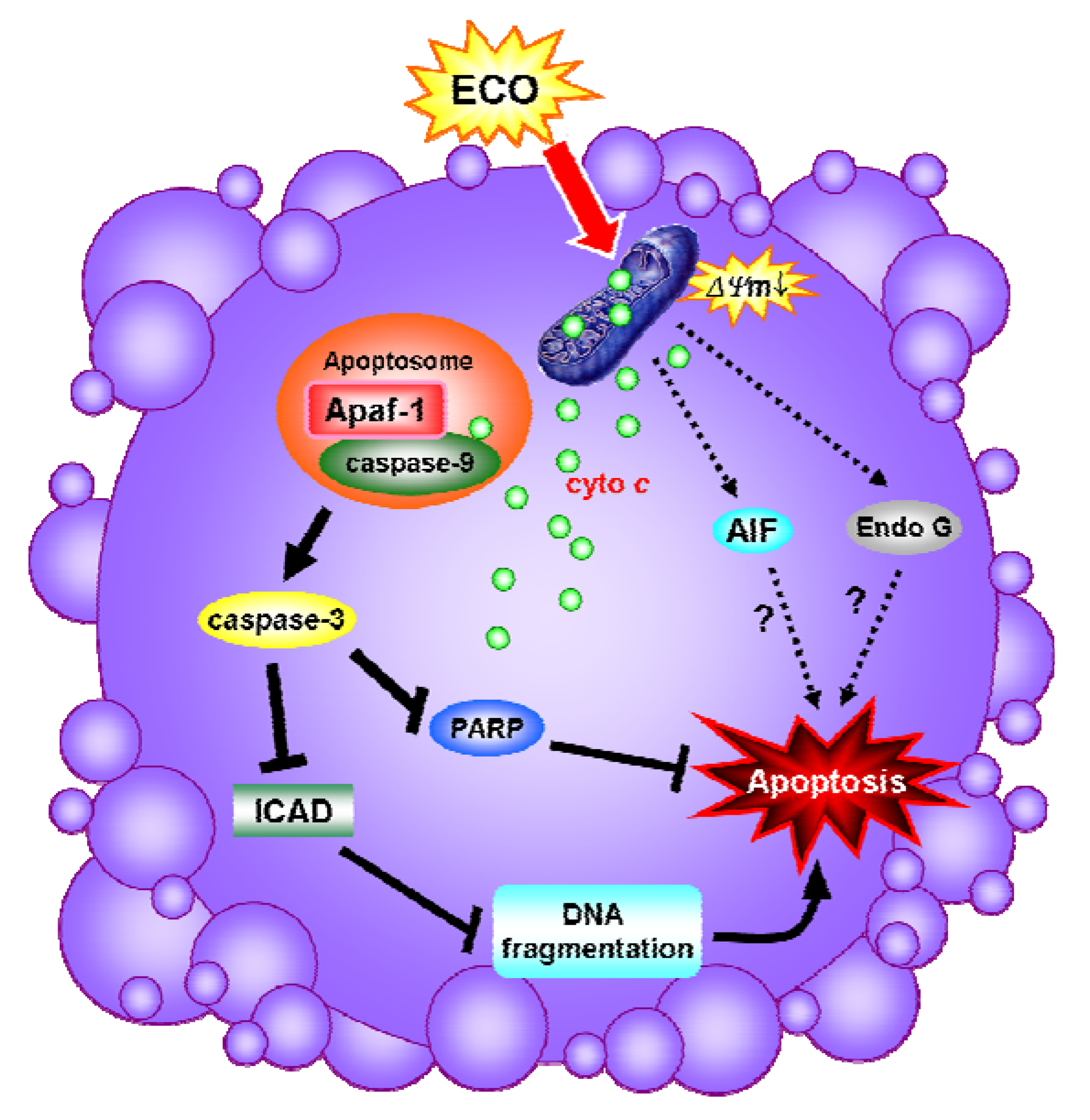
2.5. Discussion

3. Experimental
3.1. Plant Material

3.2. Cell Culture
3.3. Cell Viability Assay
3.4. Cell Cycle Analysis
3.5. DNA Fragmentation Assay
3.6. Mitochondrial Membrane Potential
3.7. Western Blot
3.8. Immunofluorescence Labeling of Cytochrome c
3.9. Statistical Analyses
4. Conclusions
Conflicts of Interest
Acknowledgments
References
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar]
- Yen, G.C.; Chen, H.Y.; Peng, H.H. Evaluation of the cytotoxicity, mutagenicity and antimutagenicity of emerging edible plants. Food Chem. Toxicol. 2001, 39, 1045–1053. [Google Scholar] [CrossRef]
- Kusamran, W.R.; Tepsuwan, A.; Kupradinun, P. Antimutagenic and anticarcinogenic potentials of some Thai vegetables. Mutat. Res. 1998, 402, 247–258. [Google Scholar] [CrossRef]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S, (Suppl. 1). [Google Scholar]
- Johnson, I.T. New approaches to the role of diet in the prevention of cancers of the alimentary tract. Mutat. Res. 2004, 551, 9–28. [Google Scholar] [CrossRef]
- Jeong, W.S.; Jun, M.; Kong, A.N. Nrf2: A potential molecular target for cancer chemoprevention by natural compounds. Antioxid. Redox Signal. 2006, 8, 99–106. [Google Scholar] [CrossRef]
- Riboli, E.; Norat, T. Epidemiologic evidence of the protective effect of fruit and vegetables on cancer risk. Am. J. Clin. Nutr. 2003, 78, 559S–569S, (Suppl. 3). [Google Scholar]
- Hardy, T.M.; Tollefsbol, T.O. Epigenetic diet: Impact on the epigenome and cancer. Epigenomics 2011, 3, 503–518. [Google Scholar]
- Azuma, K.; Nakayama, M.; Koshioka, M.; Ippoushi, K.; Yamaguchi, Y.; Kohata, K.; Yamauchi, Y.; Ito, H.; Higashio, H. Phenolic antioxidants from the leaves of Corchorus olitorius L. J. Agric. Food Chem. 1999, 47, 3963–3966. [Google Scholar]
- Ohtani, K.; Okai, K.; Yamashita, U.; Yuasa, I.; Misaki, A. Characterization of an acidic polysaccharide isolated from the leaves of Corchorus olitorius (Moroheiya). Biosci. Biotechnol. Biochem. 1995, 59, 378–381. [Google Scholar]
- Nishiumi, S.; Yabushita, Y.; Fukuda, I.; Mukai, R.; Yoshida, K.; Ashida, H. Molokhia (Corchorus olitorius L.) extract suppresses transformation of the aryl hydrocarbon receptor induced by dioxins. Food Chem. Toxicol. 2006, 44, 250–260. [Google Scholar]
- Khan, K.S.; Joergensen, R.G. Decomposition of heavy metal contaminated nettles (Urtica dioica L.) in soils subjected to heavy metal pollution by river sediments. Chemosphere 2006, 65, 981–987. [Google Scholar]
- Das, A.K.; Sahu, R.; Dua, T.K.; Bag, S.; Gangopadhyay, M.; Sinha, M.K.; Dewanjee, S. Arsenic-induced myocardial injury: Protective role of Corchorus olitorius leaves. Food Chem. Toxicol. 2010, 48, 1210–1217. [Google Scholar] [CrossRef]
- Hermann, T.; Meier, T.; Gotte, M.; Heumann, H. The 'helix clamp' in HIV-1 reverse transcriptase: A new nucleic acid binding motif common in nucleic acid polymerases. Nucleic Acids Res. 1994, 22, 4625–4633. [Google Scholar]
- Schwartsmann, G.; Ratain, M.J.; Cragg, G.M.; Wong, J.E.; Saijo, N.; Parkinson, D.R.; Fujiwara, Y.; Pazdur, R.; Newman, D.J.; Dagher, R.; Di Leone, L. Anticancer drug discovery and development throughout the world. J. Clin. Oncol. 2002, 20, 47S–59S, (Suppl. 18). [Google Scholar]
- Zeghichi, S.; Kallithraka, S.; Simopoulos, A.P. Nutritional composition of molokhia (Corchorus olitorius) and stamnagathi (Cichorium spinosum). World Rev. Nutr. Diet. 2003, 91, 1–21. [Google Scholar] [CrossRef]
- Ulivieri, C. Cell death: Insights into the ultrastructure of mitochondria. Tissue Cell 2010, 42, 339–347. [Google Scholar] [CrossRef]
- Yang, H.L.; Chen, S.C.; Chen, C.S.; Wang, S.Y.; Hseu, Y.C. Alpinia pricei rhizome extracts induce apoptosis of human carcinoma KB cells via a mitochondria-dependent apoptotic pathway. Food Chem. Toxicol. 2008, 46, 3318–3324. [Google Scholar] [CrossRef]
- Lin, Y.M.; Chen, Y.R.; Lin, J.R.; Wang, W.J.; Inoko, A.; Inagaki, M.; Wu, Y.C.; Chen, R.H. EIF3k regulates apoptosis in epithelial cells by releasing caspase 3 from keratin-containing inclusions. J. Cell Sci. 2008, 121, 2382–2393. [Google Scholar]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of poly(ADP-ribose) polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar]
- Mehmet, H. Caspases find a new place to hide. Nature 2000, 403, 29–30. [Google Scholar]
- Tait, S.W.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar]
- Otera, H.; Mihara, K. Molecular mechanisms and physiologic functions of mitochondrial dynamics. J. Biochem. 2011, 149, 241–251. [Google Scholar] [CrossRef]
- Ott, M.; Norberg, E.; Zhivotovsky, B.; Orrenius, S. Mitochondrial targeting of tBid/Bax: A role for the TOM complex? Cell Death Differ. 2009, 16, 1075–1082. [Google Scholar]
- Furumoto, T.; Wang, R.; Okazaki, K.; Hasan, A.F.M.F.; Ali, M.I.; Kondo, A.; Fukui, H. Antitumor promoters in leaves of jute (Corchorus capsularis and Cochorus olitorius). Food Sci. Technol. Res. 2002, 8, 239–243. [Google Scholar] [CrossRef]
- Komiya, T.; Kyohkon, M.; Ohwaki, S.; Eto, J.; Katsuzaki, H.; Imai, K.; Kataoka, T.; Yoshioka, K.; Ishii, Y.; Hibasami, H. Phytol induces programmed cell death in human lymphoid leukemia Molt 4B cells. Int. J. Mol. Med. 1999, 4, 377–380. [Google Scholar]
- Murakami, C.; Kumagai, T.; Hada, T.; Kanekazu, U.; Nakazawa, S.; Kamisuki, S.; Maeda, N.; Xu, X.; Yoshida, H.; Sugawara, F.; Sakaguchi, K.; Mizushina, Y. Effects of glycolipids from spinach on mammalian DNA polymerases. Biochem. Pharmacol. 2003, 65, 259–267. [Google Scholar]
- Mizushina, Y.; Hada, T.; Yoshida, H. In vivo antitumor effect of liposomes with sialyl Lewis X including monogalactosyl diacylglycerol, a replicative DNA polymerase inhibitor, from spinach. Oncol. Rep. 2012, 28, 821–828. [Google Scholar]
- Li, C.J.; Chu, C.Y.; Huang, L.H.; Wang, M.H.; Sheu, L.F.; Yeh, J.I.; Hsu, H.Y. Synergistic anticancer activity of triptolide combined with cisplatin enhances apoptosis in gastric cancer in vitro and in vivo. Cancer Lett. 2012, 319, 203–13. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, C.-J.; Huang, S.-Y.; Wu, M.-Y.; Chen, Y.-C.; Tsang, S.-F.; Chyuan, J.-H.; Hsu, H.-Y. Induction of Apoptosis by Ethanolic Extract of Corchorus olitorius Leaf in Human Hepatocellular Carcinoma (HepG2) Cells via a Mitochondria-Dependent Pathway. Molecules 2012, 17, 9348-9360. https://doi.org/10.3390/molecules17089348
Li C-J, Huang S-Y, Wu M-Y, Chen Y-C, Tsang S-F, Chyuan J-H, Hsu H-Y. Induction of Apoptosis by Ethanolic Extract of Corchorus olitorius Leaf in Human Hepatocellular Carcinoma (HepG2) Cells via a Mitochondria-Dependent Pathway. Molecules. 2012; 17(8):9348-9360. https://doi.org/10.3390/molecules17089348
Chicago/Turabian StyleLi, Chia-Jung, Shang-Yu Huang, Meng-Yu Wu, Yu-Ching Chen, Shih-Fang Tsang, Jong-Ho Chyuan, and Hsue-Yin Hsu. 2012. "Induction of Apoptosis by Ethanolic Extract of Corchorus olitorius Leaf in Human Hepatocellular Carcinoma (HepG2) Cells via a Mitochondria-Dependent Pathway" Molecules 17, no. 8: 9348-9360. https://doi.org/10.3390/molecules17089348