The Combination of Mitragynine and Morphine Prevents the Development of Morphine Tolerance in Mice

Abstract
:1. Introduction

2. Results and Discussion
2.1. Results
2.1.1. Antinociceptive Effect of Combinational Treatment of MG and Morphine

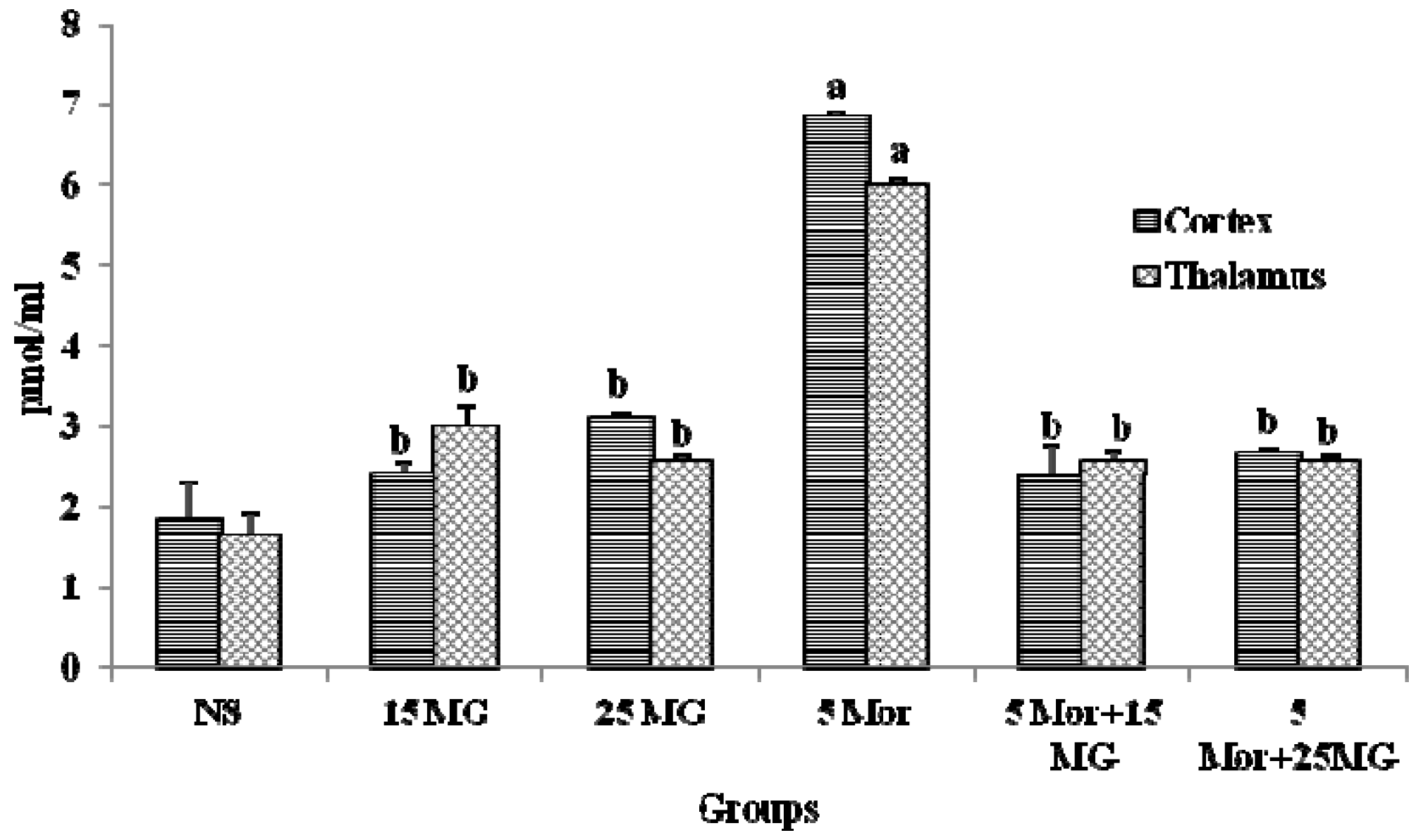
2.1.2. The Effects of MG and Morphine on the Expression of cAMP

2.1.3. The Effects of MG and Morphine on the Expression of CREB Protein

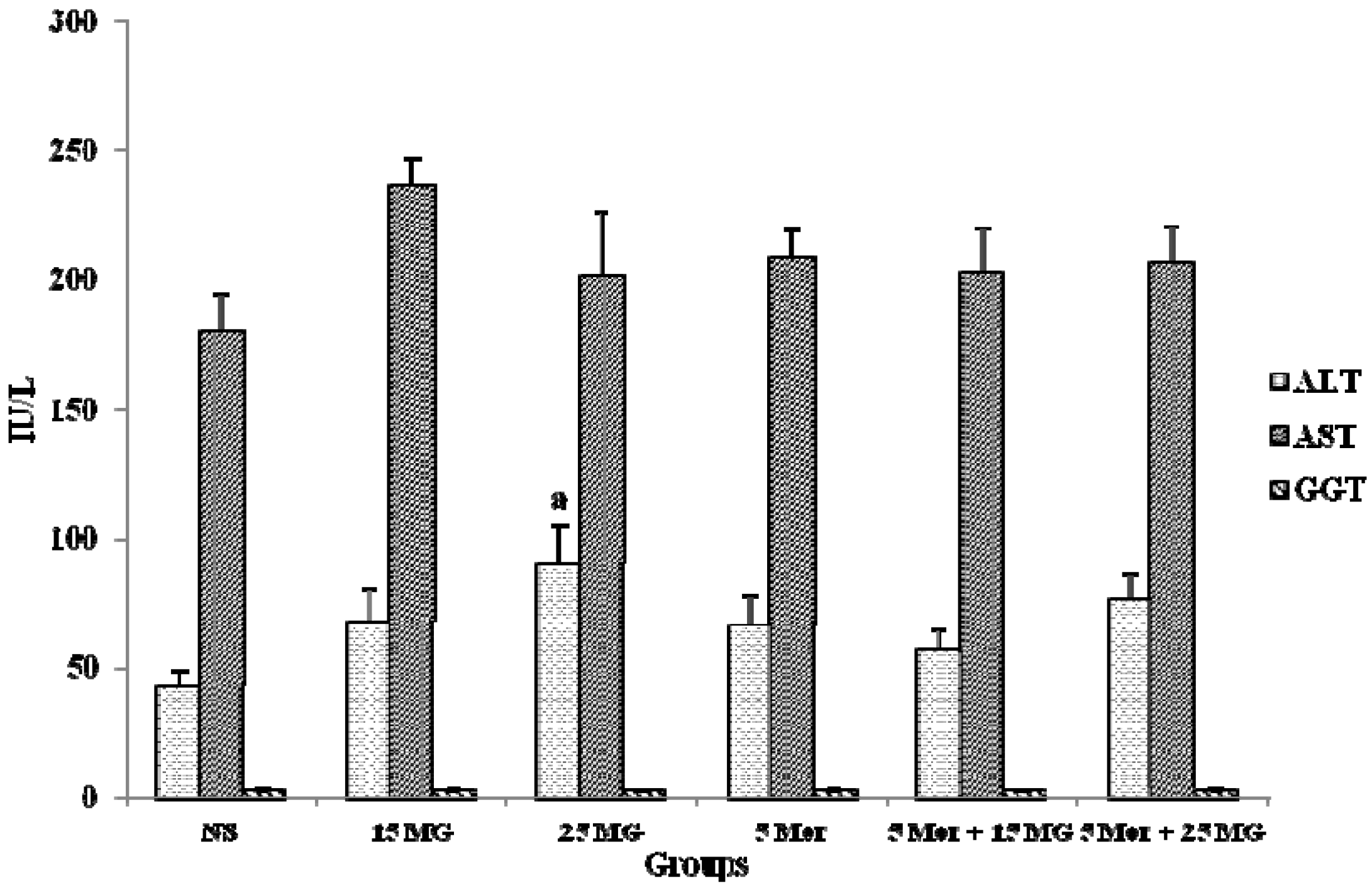
2.1.4. The Effects of MG and Morphine on liver Enzymes
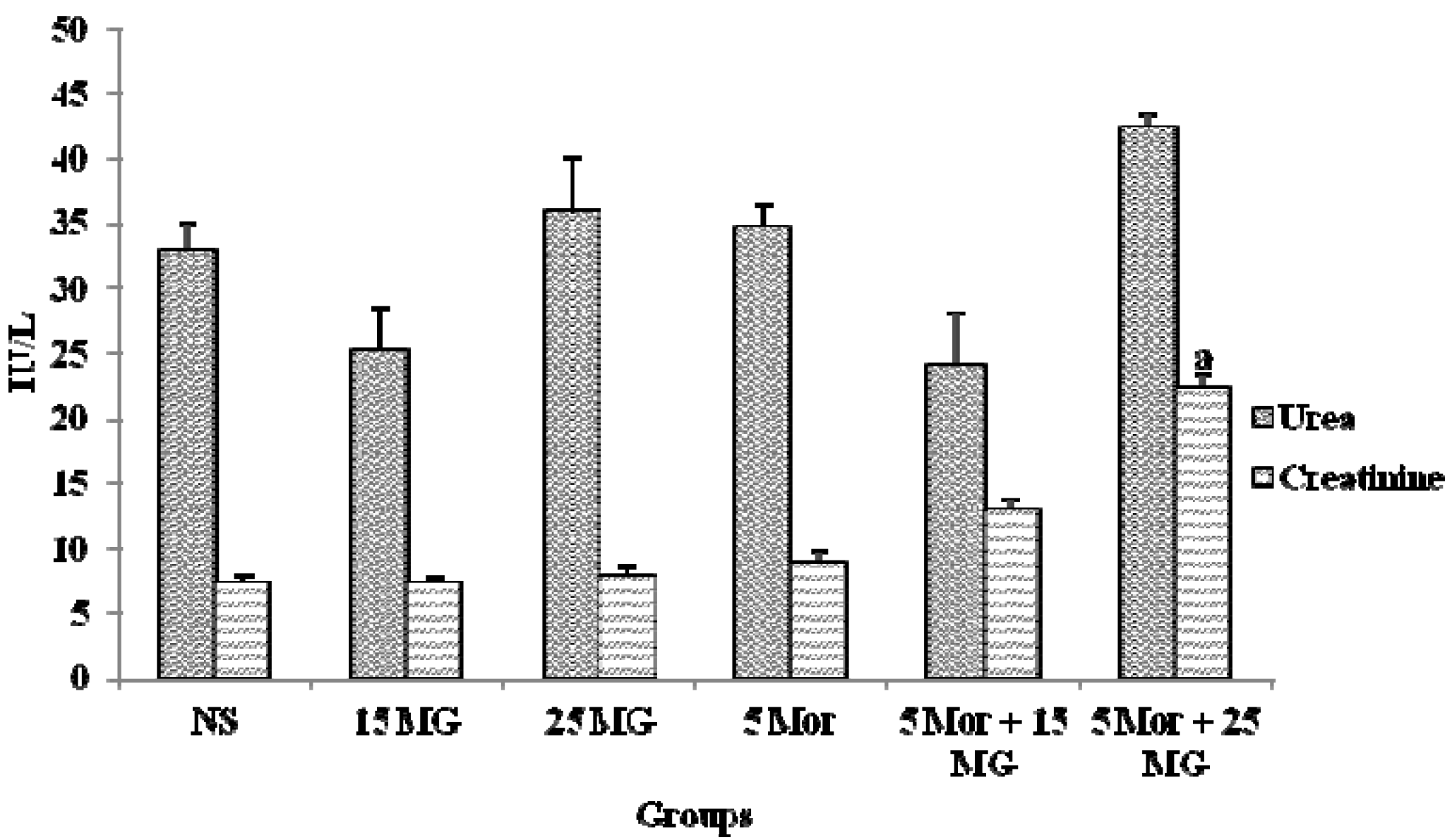
2.1.5. The Effects of MG and Morphine on Kidney Function Test


2.2. Discussion
3. Experimental
3.1. Drugs and Chemicals
3.2. Plant Materials
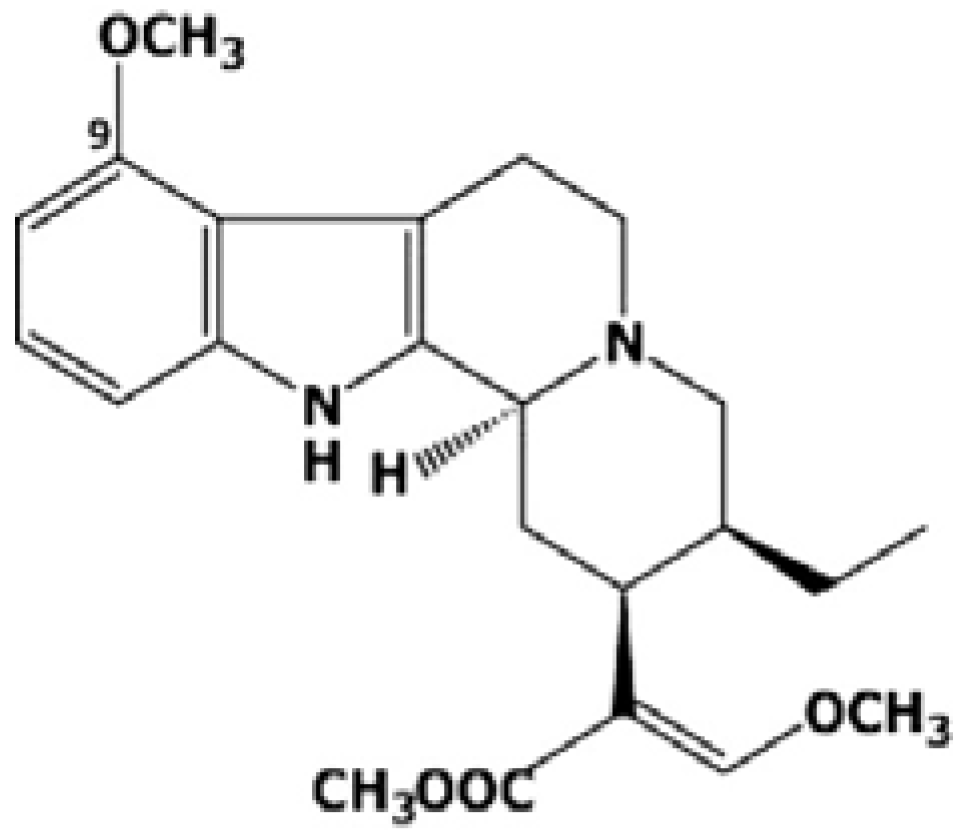
3.3. Isolation of Mitragynine from Mitragyna speciosa
3.4. Animals
3.5. Hot-Plate Test
3.6. Experimental Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Initial | Description |
|---|---|---|
| Control | NS | Normal saline |
| Treatments | 15MG | 15 mg/kg mitragynine |
| 25MG | 25 mg/kg mitragynine | |
| 5Mor | 5 mg/kg morphine | |
| 5Mor + 15MG | 5 mg/kg morphine + 15 mg/kg mitragynine | |
| 5Mor + 25MG | 5 mg/kg morphine + 25 mg/kg mitragynine |
3.7. Measurement of cAMP Level
3.8. Immunoblotting Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- McDonald, J.; Lambert, D.G. Opioid mechanisms and opioid drugs. Anaesth. Intens. Care Med. 2008, 9, 33–37. [Google Scholar] [CrossRef]
- Angeles-López, G.; Pérez-Vásquez, A.; Hernández-Luis, F.; Déciga-Campos, M.; Bye, R.; Linares, E.; Mata, R. Antinociceptive effect of extracts and compounds from Hofmeisteria schaffneri. J. Ethnopharmacol. 2010, 131, 425–432. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, Y.; Zhao, Q.; Chen, CR.; Liu, A.M.; Huang, Z.L. Curcumin exerts antinociceptive effects in a mouse model of neuropathic pain: descending monoamine system and opioid receptors are differentially involved. Neuropharmacology 2012, 62, 843–854. [Google Scholar] [CrossRef]
- Chee, J.W.; Amirul, A.A.; Majid, M.I.; Mansor, S.M. Factors influencing the release of Mitragyna speciosa crude extracts from biodegradable P(3HB-co-4HB). Int. J. Pharm. 2008, 361, 1–6. [Google Scholar] [CrossRef]
- Reanmongkol, W.; Keawpradub, N.; Sawangjaroen, K. Effects of the extracts from Mitragyna speciosa Korth. leaves on analgesic and behavioral activities in experimental animals. Songklanakarin. J. Sci. Technol. 2007, 29, 39–48. [Google Scholar]
- Peter, J.H.; Ikram, M.S. 3-dehydromitragynine: An alkaloid from Mitragyna speciosa. Phytochemistry 1986, 25, 2910–2912. [Google Scholar] [CrossRef]
- Matsumoto, K.; Takayama, H.; Ishikawa, H.; Aimi, N.; Ponglux, D.; Watanabe, K.; Horie, S. Partial agonistic effect of 9-hydroxycorynantheidine on mu-opioid receptor in the guinea-pig ileum. Life Sci. 2006, 78, 2265–2271. [Google Scholar] [CrossRef]
- Takayama, H.; Ishikawa, H.; Kurihara, M.; Kitajima, M.; Aimi, N.; Ponglux, D.; Koyama, F.; Matsumoto, K.; Moriyama, T.; Yamamoto, L.T.; et al. Studies on the synthesis and opioid agonistic activities of mitragynine-related indole alkaloids: Discovery of opioid agonists structurally different from other opioid ligands. J. Med. Chem. 2002, 45, 1949–1956. [Google Scholar]
- Takayama, H. Chemistry and pharmacology of analgesic indole alkaloids from the rubiaceous plant, Mitragyna speciosa. Chem. Pharm. Bull. (Tokyo) 2004, 52, 916–928. [Google Scholar] [CrossRef]
- Macko, E.; Weisbach, J.A.; Douglas, B. Some observations on the pharmacology of mitragynine. Archives Internationales de Pharmacodynamie et de Therapie 1972, 198, 145–151. [Google Scholar]
- Jansen, K.L.R.; Prast, C. Ethnopharmacology of kratom and the Mitragyna alkaloids. J. Ethnopharmacol. 1988, 23, 115–119. [Google Scholar] [CrossRef]
- Matsumoto, K.; Mizowaki, M.; Suchitra, T.; Murakami, Y.; Takayama, H.; Sakai, S.; Aimi, N.; Watanabe, H. Central antinociceptive effects of mitragynine in mice: Contribution of descending noradrenergic and serotonergic systems. Eur. J. Pharmacol. 1996, 317, 75–81. [Google Scholar] [CrossRef]
- Matsumoto, K.; Mizowaki, M.; Suchitra, T.; Takayama, H.; Sakai, S.; Aimi, N.; Watanabe, H. Antinociceptive action of mitragynine in mice: Evidence for the involvement of supraspinal opioid receptors. Life Sci. 1996, 59, 1149–1155. [Google Scholar] [CrossRef]
- Yamamoto, L.T.; Horie, S.; Takayama, H.; Aimi, N.; Sakai, S.; Yano, S.; Shan, J.; Pang, P.K.; Ponglux, D.; Watanabe, K. Opioid receptor agonistic characteristics of mitragynine pseudoindoxyl in comparison with mitragynine derived from Thai medicinal plant Mitragyna speciosa. Gen. Pharmacol. 1999, 33, 73–81. [Google Scholar] [CrossRef]
- Shamima, A.R.; Fakurazi, S.; Hidayat, M.T.; Hairuszah, I.; Moklas, M.A.M.; Arulselvan, P. Antinociceptive Action of Isolated Mitragynine from Mitragyna Speciosa through Activation of Opioid Receptor System. Int. J. Mol. Sci. 2012, 13, 11427–11442. [Google Scholar]
- Lauretti, G.R.; Lima, I.C.; Reis, M.P.; Prado, W.A.; Pereira, N.L. Oral ketamine and transdermal nitroglycerin as analgesic adjuvants to oral morphine therapy for cancer pain management. Anesthesiology 1999, 90, 1528–1533. [Google Scholar] [CrossRef]
- Mercadante, S. Opioid rotation for cancer pain: Rationale and clinical aspects. Cancer 1999, 86, 1856–1866. [Google Scholar] [CrossRef]
- White, P.F. The changing role of non-opioid analgesic techniques in the management of postoperative pain. Anesth. Analg. 2005, 101, S5–S22. [Google Scholar] [CrossRef]
- Sharma, S.K.; Nirenberg, M.; Klee, W.A. Morphine receptors as regulators of adenylate cyclase activity. Proc. Natl. Acad. Sci. USA 1975, 72, 590–594. [Google Scholar]
- Brandt, M.; Fischer, K.; Moroder, L.; Wunsch, E.; Hamprecht, B. Enkephalin evokes biochemical correlates of opiate tolerance and dependence in neuroblastoma x glioma hybrid cells. FEBS Lett. 1976, 68, 38–40. [Google Scholar] [CrossRef]
- Boshart, M.; Weih, F.; Schmidt, A.; Fournier, R.E.; Schütz, G.A. Cyclic AMP response element mediates repression of tyrosine aminotransferase gene transcription by the tissue-specific extinguisher locus Tse-1. Cell 1990, 61, 905–916. [Google Scholar] [CrossRef]
- Meyer, T.E.; Habener, J.F. Cyclic adenosine 3',5'-monophosphate response element binding protein (CREB) and related transcription-activating deoxyribonucleic acid-binding proteins. Endocr. Rev. 1993, 14, 269–290. [Google Scholar]
- Sheng, M.; Thompson, M.A.; Greenberg, M.E. CREB: A Ca(2+)-regulated transcription factor phosphorylated by calmodulin-dependent kinases. Science 1991, 252, 1427–1430. [Google Scholar]
- Widnell, K.L.; Russell, D.S.; Nestler, E.J. Regulation of expression of cAMP response element-binding protein in the locus coeruleus in vivo and in a locus coeruleus-like cell line in vitro. Proc. Natl. Acad. Sci. USA 1994, 91, 10947–10951. [Google Scholar]
- Coven, E.; Ni, Y.; Widnell, K.L.; Chen, J.; Walker, W.H.; Habener, J.F.; Nestler, E.J. Cell type-specific regulation of CREB gene expression: mutational analysis of CREB promoter activity. J. Neurochem. 1998, 71, 1865–1874. [Google Scholar]
- Shaw-Lutchman, T.Z.; Barrot, M.; Wallace, T.; Gilden, L.; Zachariou, V.; Impey, S.; Duman, R.S.; Storm, D.; Nestler, E.J. Regional and cellular mapping of cAMP response element-mediated transcription during naltrexone-precipitated morphine withdrawal. J. Neurosci. 2002, 22, 3663–3672. [Google Scholar]
- Chartoff, E.H.; Papadopoulou, M.; Konradi, C.; Carlezon, W.A., Jr. Dopamine-dependent increases in phosphorylation of cAMP response element binding protein (CREB) during precipitated morphine withdrawal in primary cultures of rat striatum. J. Neurochem. 2003, 87, 107–118. [Google Scholar] [CrossRef]
- Raffa, R.B. Pharmacology of oral combination analgesics: Rational therapy for pain. J. Clin. Pharm. Ther. 2001, 26, 257–264. [Google Scholar] [CrossRef]
- Du Pen, A.; Shen, D.; Ersek, M. Mechanisms of opioid-induced tolerance and hyperalgesia. Pain Manag. Nurs. 2007, 8, 113–121. [Google Scholar] [CrossRef]
- Wecker, L.; Crespo, L.M.; George, D.; Carl, F.; Stephanie, W. Brody’s Human Pharmacology Molecular to Clinical; Mosby Elsevier: Philadelpia, PA, USA, 2010; pp. 74–78. [Google Scholar]
- Nicholson, G.P. Treatment of anterior superior shoulder instability with a reverse ball and socket prosthesis. Oper. Technol. Orthop. 2003, 13, 235–241. [Google Scholar] [CrossRef]
- Noble, F.; Cox, B.M. Differential desensitization of mu- and delta-opioid receptors in selected neural pathways following chronic morphine treatment. Br. J. Pharmacol. 1996, 117, 161–169. [Google Scholar] [CrossRef]
- Sim, L.J.; Selley, D.E.; Dworkin, S.I.; Childers, S.R. Effects of chronic morphine administration on mu opioid receptor-stimulated [35S] GTPgammaS autoradiography in rat brain. J. Neurosci. 1996, 16, 2684–2692. [Google Scholar]
- Collier, H.O.; Francis, D.L. Morphine abstinence is associated with increased brain cyclic AMP. Nature 1975, 255, 159–162. [Google Scholar]
- Nestler, E.J. Genes and addiction. Nat. Genet. 2000, 26, 277–281. [Google Scholar] [CrossRef]
- Matsumoto, K.; Horie, S.; Ishikawa, H.; Takayama, H.; Aimi, N.; Ponglux, D.; Watanabe, K. Antinociceptive effect of 7-hydroxymitragynine in mice: Discovery of an orally active opioid analgesic from the Thai medicinal herb Mitragyna speciosa. Life Sci. 2004, 74, 2143–2155. [Google Scholar] [CrossRef]
- Sim-Selley, L.J.; Selley, D.E.; Vogt, L.J.; Childers, S.R.; Martin, T.J. Chronic heroin self-administration desensitizes mu opioid receptor-activated G-proteins in specific regions of rat brain. J. Neurosci. 2000, 20, 4555–4562. [Google Scholar]
- Yabaluri, N.; Medzihradsky, F. Down-regulation of mu-opioid receptor by full but not partial agonists is independent of G protein coupling. Mol. Pharmacol. 1997, 52, 896–902. [Google Scholar]
- Nestler, E.J. Cellular responses to chronic treatment with drugs of abuse. Crit. Rev. Neurobiol. 1993, 7, 23–39. [Google Scholar]
- Bilecki, W.; Przewłocki, R. Effect of opioids on Ca2+/cAMP responsive element binding protein. Acta Neurobiol. Exp. (Wars). 2000, 60, 557–567. [Google Scholar]
- Bilecki, W.; Wawrzczak-Bargiela, A.; Przewlocki, R. Activation of AP-1 and CRE-dependent gene expression via mu-opioid receptor. J. Neurochem. 2004, 90, 874–882. [Google Scholar] [CrossRef]
- Beggs, S.; Micheal, WS. A straightjacket for pain? Cell 2010, 143, 505–507. [Google Scholar] [CrossRef]
- Jurgen, S. Understanding LTP in Pain Pathway. Mol. Pain 2007, 3, 100–106. [Google Scholar]
- Simone, C.; Bosshard, C.B.; Matthias, T.; Wyss, T.M.; Bruno, W.; Markus, R. Assessment of Pain responses to innocuous and noxious electrical forepaw stimulation in mice using BOLD fMRI. Pain 2010, 655–663. [Google Scholar]
- Guitart, X.; Thompson, M.A.; Mirante, C.K.; Greenberg, M.E.; Nestler, E.J. Regulation of cyclic AMP response element-binding protein (CREB) phosphorylation by acute and chronic morphine in the rat locus coeruleus. J. Neurochem. 1992, 58, 1168–1171. [Google Scholar] [CrossRef]
- Ozsoy-Sacan, O.; Yanardag, R.; Orak, H.; Ozgey, Y.; Yarat, A.; Tunali, T. Effects of parsley (Petroselinum crispum) extract versus glibornuride on the liver of streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2006, 104, 175–181. [Google Scholar] [CrossRef]
- Singh, S.N.; Vats, P.; Suri, S.; Shyam, R.; Kumria, M.M.; Ranganathan, S.; Sridharan, K. Effect of an antidiabetic extract of Catharanthus roseus on enzymic activities in streptozotocin induced diabetic rats. J. Ethnopharmacol. 2001, 76, 269–277. [Google Scholar] [CrossRef]
- Mulder, G.J. Pharmaceutical Toxicology: Safety Sciences of Drugs; Dencker, L., Ed.; RPS Publishing: London, UK, 2000; pp. 137–160. [Google Scholar]
- Sample Availability: Samples of the mitragynine compounds are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fakurazi, S.; Rahman, S.A.; Hidayat, M.T.; Ithnin, H.; Moklas, M.A.M.; Arulselvan, P. The Combination of Mitragynine and Morphine Prevents the Development of Morphine Tolerance in Mice. Molecules 2013, 18, 666-681. https://doi.org/10.3390/molecules18010666
Fakurazi S, Rahman SA, Hidayat MT, Ithnin H, Moklas MAM, Arulselvan P. The Combination of Mitragynine and Morphine Prevents the Development of Morphine Tolerance in Mice. Molecules. 2013; 18(1):666-681. https://doi.org/10.3390/molecules18010666
Chicago/Turabian StyleFakurazi, Sharida, Shamima Abdul Rahman, Mohamad Taufik Hidayat, Hairuszah Ithnin, Mohamad Aris Mohd Moklas, and Palanisamy Arulselvan. 2013. "The Combination of Mitragynine and Morphine Prevents the Development of Morphine Tolerance in Mice" Molecules 18, no. 1: 666-681. https://doi.org/10.3390/molecules18010666