Pregnenolone Sulfate: From Steroid Metabolite to TRP Channel Ligand

1
Department of Pharmacology and Experimental Therapy, Institute of Experimental and Clinical Pharmacology and Toxicology, Eberhard Karls University, Tübingen 72074, Germany
2
Hospitals and Clinics, Interfaculty Center of Pharmacogenomics and Drug Research, University of Tübingen, Tübingen 72074, Germany
Molecules 2013, 18(10), 12012-12028; https://doi.org/10.3390/molecules181012012
Submission received: 2 September 2013
/
Revised: 20 September 2013
/
Accepted: 21 September 2013
/
Published: 27 September 2013
(This article belongs to the Special Issue Steroids)
Abstract
:Pregnenolone sulfate is a steroid metabolite with a plethora of actions and functions. As a neurosteroid, pregnenolone sulfate modulates a variety of ion channels, transporters, and enzymes. Interestingly, as a sulfated steroid, pregnenolone sulfate is not the final- or waste-product of pregnenolone being sulfated via a phase II metabolism reaction and renally excreted, as one would presume from the pharmacology textbook knowledge. Pregnenolone sulfate is also the source and thereby the starting point for subsequent steroid synthesis pathways. Most recently, pregnenolone sulfate has been functionally “upgraded” from modulator of ion channels to an activating ion channel ligand. This review will focus on molecular aspects of the neurosteroid, pregnenolone sulfate, its metabolism, concentrations in serum and tissues and last not least will summarize the functional data.
1. Introduction
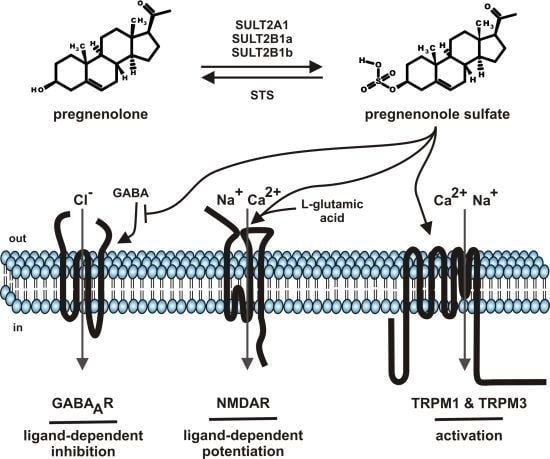
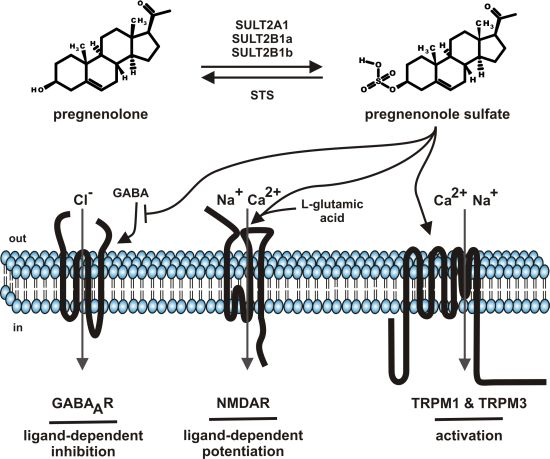
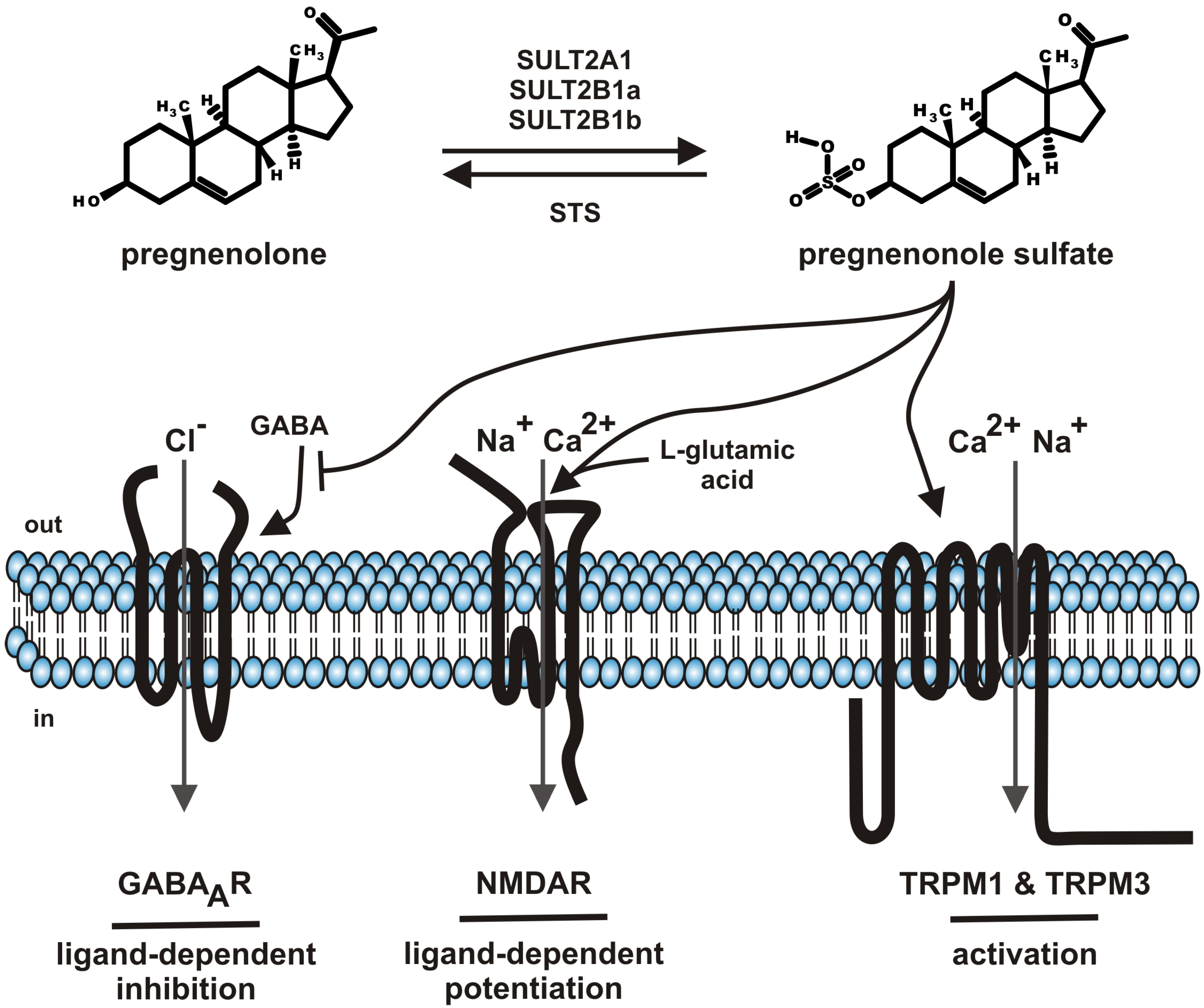
Steroids comprise a group of chemical related hormones mediating their function by modulation of transcriptional activity. Steroids like cortisol, estradiol, testosterone, or aldosterone bind to intracellular cytosolic glucocorticoid receptor, estrogen receptor, androgen receptor or mineralocorticoid receptor, respectively. The receptor-hormone complex subsequently translocates from the cytosol to the cell nucleus and regulates transcriptional activity. The genomic effects of steroid hormone application are detectable with some delay. Beside the genomic effects of steroids, fast, membrane-delimited non-genomic effects of steroids have been described [1]. The question of specificity dominated this field. On one hand, reports showed that such effects, e.g., modulation of GABAA receptor by neuroactive steroids depends on the cholesterol content of the plasma membrane by using methyl-β-cyclodextrin as tool to interfere with the endogenous cholesterol content of the plasma membrane [2]. On the other hand, a growing spectrum of steroid compounds, intermediates from steroid synthesis, metabolites (sulfate-, acetate conjugates), most of them designated as neurosteroids, helped to unravel the diversity of specific effects of membrane-delimited steroid actions [3,4,5]. This review will focus on one particular neurosteroid, pregnenolone sulfate (PregS, Figure 1) as it has been functionally “upgraded” from modulator of ion channels to an activating ion channel ligand by the discovery that PregS activates TRPM3, an ion channel of the TRP superfamily [6]. The review will sum data on metabolism, serum and tissue concentrations and functional data on PregS.
2. Pregnenolone Sulfate—Synthesis and Metabolism
Steroid sulfates are formed by sulfonation of steroids by cytosolic sulfotransferase enzymes (SULT) [7,8]. So far more than 40 distinct cytosolic sulfotransferase enzymes have been identified from which three enzymes (SULT1E, SULT2B1, SULT2A1) selectively sulfonate steroids with distinct profiles of steroid specificities. SULT1E, SULT2B1, SULT2A1 mediate the sulfonation of estrogen, nonaromatic steroids and sterols, respectively. PregS is generated by SULT2A1, SULT2B1a and SULT2B1b (Figure 1) [8]. Based on the NCBI EST profiles, SULT2A1 mRNA is transcribed preferentially in the adrenal gland, liver, parathyroid gland, whereas the SULT2B1 transcripts show reduced mRNA counts widely distributed in many tissues like ascites, brain, connective tissue, eye, heart, intestine, kidney, lung, mammary gland, pancreas, placenta, prostate, and testis. With respect to distribution of sulfotransferases in brain, recently the frog SULT enzyme has been localized to the GnRH-innervated regions of the magnocellular nucleus and anterior preoptic area [9]. The broad tissue distribution of SULT enzyme argues for the generation of PregS in many tissues and its different tissue–specific roles.
Differences in SULT2B1 enzymes result from splice events leading to two different mRNAs encoding N-terminally diverging proteins SULT2B1a and SULT2B1b [10]. The splice differences are accompanied by distinct substrate selectivities. SULT2B1b is highly selective for cholesterol being the cholesterol sulfate forming enzyme in the skin, liver, and elsewhere, SULT2B1a ignores cholesterol and prefers pregnenolone [8]. Pregnenolone is synthesized from cholesterol by a mitochondrially localized cytochrome P450 side-chain cleavage enzyme (scc, CYP11A1) [4,11]. This central step in steroidogenesis is rate limiting and essential. Diethylstilbestrol, tamoxifen, cocaine, and other drugs have been described to interfere with steroidogenesis by downregulation of cytochrome P450 side-chain cleavage enzyme [12,13,14]. SULT2B1a expression is regulated by l-glutamic acid via AMPA receptor-mediated NO signaling [15], which is interesting as PregS potentiates l-glutamic acid transduction via N-methyl-d-aspartate receptors (NMDAR) (see below).
In the context of xenobiotic metabolisms, sulfonation is discussed as an endpoint leading to increased water solubility and enhanced renal clearance of the metabolite [16]. However, steroid sulfates additionally represent ion channel modulators and storage intermediates. As source and building block for steroid synthesis, steroid sulfates had to enter cells for sulfohydrolase reaction. The passive passage and diffusion of steroid sulfates are hampered by hydrophilicity of the sulfate moiety, for which reason cellular uptake of steroid sulfates is dependent on transporters (see below) [17,18]. The steroid releasing reaction is mediated by steroid sulfatase (STS) an enzyme encoded by a X-chromosomally localized gene [19,20,21]. The physiological impact of STS can be anticipated by the fact that the congenital deficit of STS is causing X-linked recessive ichthyosis [22]. The disease affecting males in an incidence of 1: 2000–6000, is characterized by dry, scaly skin due to deletions or mutations in the STS gene and subsequent deficits in steroid hormone function [22]. In addition, biochemical studies using radioactive labeling approaches showed the release of pregnenolone and subsequent steroid metabolites from PregS [23,24,25,26].
Figure 1.
Summary of pregnenolone sulfate synthesis, metabolism and functions.

SULT: cytosolic sulfotransferase enzymes; STS: steroid sulfatase; GABA: gamma-aminobutyric acid; GABAAR: ionotropic receptor of gamma-aminobutyric acid; NMDAR: ionotropic receptor of l-glutamic acid, N-methyl-D-aspartate subtype; TRPM: members of the melastatin-like transient receptor potential channel (TRPM) subfamily; out: outside the cell, extracellular; in: inside the cell, intracellular.
3. Pregnenolone Sulfate Levels in Serum and Tissues
Human serum PregS concentrations change during lifespan from µM concentrations during birth to nM concentrations in adults [27]. The mean PregS concentrations during birth are 3.06 µM (♀) and 2.5 µM (♂) in the umbilical cord and 1.85 µM (♀) and 2.2 µM (♂) in fetal serum [27]. The PregS levels decline rapidly during the first two weeks to 1.02 µM (♀) and 1.4 µM (♂), reaching levels of 56 nM (♀) and 32 nM (♂) in the first year [27]. These levels are more or less stable during childhood and then double during adolescence, reaching levels of 130 nM (♀) and 140 nM (♂) in adults [27]. The high PregS levels during birth are indicative for the role of PregS during pregnancy and birth, for which reason the PregS levels during this period induced a variety of studies analyzing PregS levels during pregnancy and birth [28,29,30,31,32,33,34,35]. PregS in maternal plasma (P), umbilical artery (UA), umbilical vein (UV) rises from 930 nM (P), 3.8 µM (UA), 3.6 µM (UV) during pregnancy to 1250 nM (P), 4.8 µM (UA), 3.9 µM (UV) during birth [29,33]. A study in twins revealed that PregS plays an important role for the onset of parturition [34]. The increase of PregS before and during parturition is so far published a unique feature in humans. The increase in concentration prior parturition seems to be a species–specific regulation mechanism, as in contrast in sheep, the PregS concentrations rise during pregnancy and rapidly decline from day 3 before birth in fetal as well as maternal plasma [36]. However, the absence of large amplitudes during the oestrous cycle in seems to be a common feature in mammals. PregS levels in ewes show only small deviations from the mean values [37]. Values for women are only available from women suffering from premenstrual syndrome also show only small deviations in PregS levels during the oestrous cycle [38]. In contrast to the data situation on PregS in women during the oestrous cycle, several studies analyzed PregS occurrence in testis from men who have undergone orchiectomy due to prostatic carcinoma [39,40,41]. Whereas the peripheral PregS serum levels are in the range of the concentrations mentioned above, the PregS concentrations in spermatic venous blood were 3–4 fold increased [41]. In testis tissue, PregS concentrations reach 2.8 to 4.0 µmol/kg, in vas deferens and epididymis and 1.4 µmol/kg and 0.76 µmol/kg, respectively [39,40].
The dynamic regulation of PregS levels during pregnancy illustrates that steroid as well as neurosteroid levels can change due to physiological adaptation. Adrenal stimulation in children increased PregS parallel to cortisol levels enabling the usage of PregS in diagnosis of various adrenal and pituitary diseases in children [27,42]. A more general role of PregS became evident by reports of changed PregS levels in disease states [43,44,45,46,47]. For example, PregS serum concentrations are reduced in hypothyroidism and 7fold increase in hyperthyroidism [46]. In patients suffering from rheumatoid arthritis, PregS levels are 4fold reduced [45].
The occurrence of PregS in the brain has been a matter of debate influenced by imponderabilities of the analytic methods summarized in a review by Schumacher and coauthors [48]. The impact of PregS as neurosteroid in disease has been documented by a report showing that PregS tissue concentrations decreased in Alzheimer patients [49]. The correlation of high levels of key proteins implicated in the formation of plaques and neurofibrillary tangles with the decreased levels of PregS and other neurosteroids suggest a possible neuroprotective role of PregS in Alzheimer disease [49]. In human brain, PregS concentrations vary from 5 to 42 nmol/kg PregS, depending on brain region, with the highest concentrations of 35 and 42 nmol/kg PregS in striatum and hypothalamus, respectively [49]. Comparable values were published for rat brain PregS concentrations of 45 nmol/kg [50,51]. Interestingly, a study analyzing neurosteroid levels in developing rat brain upon fetal alcohol exposure revealed that fetal alcohol exposure by maternal ethanol consumption increase PregS level in brain during fetal development and last till postnatal day 5 [52]. Enhanced PregS levels were restricted to the fetal brain. The maternal brain, maternal blood, placenta, and fetal liver were not affected excluding secondary accumulation of peripherally-produced steroids [52]. In the context with recent finding of PregS-induced, TRPM3-mediated potentiation of synapse formation in Purkinje cells (see below), fetal alcohol exposure leading to enhanced PregS levels might be responsible for an inadequate neuronal architecture. Daily subcutaneous application of 20 mg/kg PregS protected the survival and maturation of newborn neuronal cells in APP/PS1 transgenic mice, a mouse model for Alzheimer disease [53]. Concurrently, the spatial cognitive performance was improved in the PregS-treated mice [53].
4. Transporter—Cellular Uptake of Pregnenolone Sulfate
It is unlikely that pregnenolone sulfate is capable of easily crossing the plasma membrane due to the hydrophilic sulfate moiety. On the other hand, for its function as source for pregnenolone and subsequent synthesis of steroid hormones like estradiol, progesterone etc., it is indispensible that the substrate (PregS) enters the cytosol to come into contact with the cytosolic localized sulfohydrolases. The transmembrane transport can be facilitated by a variety of transporter proteins [17,18]. The organic anion-transporting polypeptide (OATP-B) has been cloned from placenta as steroid conjugate transporting protein [54]. OATP-B is nowadays classified as solute carrier organic anion transporter (SLCO2B1). OATP-B/SLCO2B1 is expressed in many organs like liver, mammary gland, brain and intestine, whereas other organic anion transporting polypeptides (OATP)-A or C, are limited to special tissues (such as liver or brain) [55]. OATP-B/SLCO2B1 has been shown to be localized in the placental basolateral membrane compartment mediating estrone-sulfate uptake being inhibited by PregS [54]. The transport properties are quite complex as later studies showed that OATP-B/SLCO2B1 mediates PregS uptake, which is further enhanced in the presence of progesterone, whereas PregS blocked 28% ± 7% and 43% ± 7% of the estron sulfate uptake and dehydroepiandrosterone sulfate uptake, respectively [55].
A high capability to transport PregS has also been shown for a transport protein of another subfamily, the sodium-dependent organic anion transport (SOAT) nowadays, solute carrier family 10 (sodium/bile acid cotransporter) (SLC10A6) [56,57]. With respect to function of PregS as neurosteroid, the organic solute transporter OSTα-OSTβ is of particular interest [58]. The transport protein of the organic solute transporter family (SLC51) is expressed in the human tissues liver, ovary, adrenal gland and hippocampus. By laser capture microdissection RT-PCR analysis, mRNA transcription of OSTα-OSTβ was detected in Purkinje cells and cells in the CA region of the hippocampus murine Purkinje and hippocampal cells, the steroidogenic cells of the brain [58]. PregS as well as DHEAS represent relatively high affinity substrates for OSTα-OSTβ/SLC51. Estron sulfate uptake in the presence of PregS or DHEAS, results in inhibition of half of the transport [58].
5. Molecular Targets of Pregnenolone Sulfate
A broad range of ion channels have been shown to be modulated by PregS. Many reports remained anecdotally like the modulation of potassium channels [59,60] or nicotinic acetylcholine receptor [61] or voltage-gated sodium channels [62]. In contrast (Figure 1), the negative modulation of GABAA chloride channels, the positive modulation of glutamate response by NMDA receptors and the activation of TRPM1 as well as TRPM3 channels are well established and will be the topics of the following chapters (Table 1).

{kind=link}
{kind=link}
| Molecular target | Mode of action | EC50/IC50 values | Physiological impact |
|---|---|---|---|
| GABAA channel | inhibition | IC50 7.2 µM | long term potentiation, memory & learning, anxiolysis, general anesthesia epilepsy, musclar cramps |
| (GABA 3 µM) [63] | |||
| NMDA receptor | potentiation | EC50 33 µM | neuronal development, synapse formation |
| (NMDA 5 µM) [64] | |||
| TRPM1 | activation | melanocyte function, melanin synthesis, phototransduction | |
| TRPM3 | activation | EC50 23 µM (– 80 mV) [6] | pain modulation, insulin secretion, neuronal development |
| EC50 12 µM (+ 80 mV) [6] | |||
| [Pregnenolone EC50 15 µM | |||
| (–80 mV)] [6] | |||
| [Pregnenolone EC50 14 µM | |||
| (+80 mV)] [6] |
5.1. GABAA Channels
The clearance of barbiturate-induced hypnosis by PregS was the starting point for the characterization of interaction of PregS with the GABAA receptor [65]. Subsequent studies showed binding to and functional inhibition of GABAA receptors [66,67]. The modulation of GABAA receptors by PregS seems to be limited to a distinct subunit composition [68]. GABAA receptors reassembled from α1,2,3 + β1 + γ2 are sensitive to PregS-mediated inhibition [68]. The effect of PregS on GABAA inhibition is independent of ethanol, whereas modulation by allopregnanolone, another neurosteroid, was affected [69]. The neurosteroid effect on GABAA receptors is phylogenetically conserved. UNC-49, the C. elegans orthologue of mammalian GABAA receptors, is modulated by PregS [70]. The transcription of two splice variants of C. elegans UNC-49 resulting in the translation of UNC-49B and UNC-49C differing in neurosteroid sensitivity allowed to map the region of neurosteroid interaction [70]. Site-directed mutagenesis und functional analysis showed that a region of 6 residues in the first transmembrane domain (M1) mediates PregS modulation [70]. However transferring these finding to the mammalian GABAA receptor orthologue showed quite different results [71]. Therefore, it is unclear whether the PregS interaction site is localized elsewhere or formed by M1 in conjunction with structure-stabilizing amino acids of the other transmembrane segments (M2, M3, M4). GABA as neurotransmitter plays a central role in the neuronal network allowing the pharmacological modulation to achieve anxiolysis and general anesthesia or interfere in epilepsy or muscular cramps. Therefore, a variety of central as well as peripheral effects of PregS have been linked to its ability to interfere with GABA function [5].
5.2. NMDA Receptors
PregS is an allosteric modulator at the l-glutamic acid receptor [63]. With respect to selectivity, it is remarkable that selectively N-methyl-d-aspartate (NMDA) receptors are modulated in a positive way, whereas α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPAR) or kainate receptors are negative regulated [63]. The half-maximal concentration of PregS (EC50 value) in the presence of 5 µM NMDA is 33 µM (Table 1) [64]. The effects of PregS can be shifted to nanomolar concentrations in the presence of the NMDA receptor antagonists, ifenprodil or dizocilpine (MK-801) [72,73]. NMDA receptors assemble as obligate heteromers that may be drawn from NR1, NR2A, NR2B, NR2C, NR2D, NR3A and NR3B subunits [74]. Splicing events increase variability of the NMDA receptor family [74]. The magnitude of PregS-mediated effect tested on the NR1a/NR2A and NR1a/NR2B heteromers vary showing a larger effect on NR1a/NR2A [75]. Further studies revealed that PregS potentiated NMDA-induced currents of NR1/NR2A and NR1/NR2B receptors, whereas it was inhibitory at NR1/NR2C and NR1/NR2D receptors [76,77]. The effect of PregS on NMDA receptors is dependent on protein phosphorylation as protein kinase inhibitors as well as protein phosphatase inhibitors can interfere with the PregS-mediated modulation of NMDA receptors [78]. Additionally, NMDA receptor function is modulated by several endogenous molecules, including zinc, polyamines, protons, and sulfated neurosteroids. Zinc, polyamines, and phenylethanolamines exert their modulatory role by inferring with inhibition via protons. Proton-dependent inhibition depends on aspartate residue (D690 in NR1B) in the extracellular loop adjacent to the M3 transmembrane domain [79]. Detailed analysis showed corresponding domains in NR2A and revealed that additional residues are involved in proton-dependent inhibition [80]. PregS binding was also mapped to S2 extracellular loop however to another region adjacent to the M4 transmembrane domain. [81], which argues a unique mechanism for NMDA receptor enhancement independent of the proton sensor. In addition to the acutely affecting modulation of NMDA receptor, PregS enhance NMDA receptor function by increasing NMDA receptor insertion in the cell surface [82]. The enhanced integration of NMDA receptor proteins in the plasma membrane is mediated via a non-canonical, pertussis toxin–sensitive, G protein–coupled, and calcium-dependent mechanism independent of NMDA receptor activation [82].
Glutamate receptors play a central role in neuronal regulation [83]. Therefore, a variety of reports describe PregS-mediated potentiation of NMDA receptors which are involved in many processes like learning and memory [84,85,86], central regulation of body functions [87] or effects of drugs like ethanol [87], opioids [88]. In many of these reports, the effects have been assigned based on the use of a spectrum of pharmacological tools designated to interfere with NMDA receptors. However, some cautions is appropriate with respect to findings that PregS activates TRPM1 and TRPM3 [6,89]. Starting from the finding that PregS potentiates spontaneous glutamate release onto neonatal Purkinje cells during a period of active glutamatergic synapse formation [90], we showed that the potentiation is mediated by the direct activation of TRPM3 channels expressed in Purkinje cells [91].
5.3. TRP Channels (TRPM1, TRPM3)
TRPM1 and TRPM3 are members of the mammalian melastatin-like transient receptor potential (TRPM) channel subfamily including eight members based on their homology to melastatin (TRPM1), a putative tumor suppressor involved in the pathophysiology of melanoma [92]. Proteins of the TRPM channel, the classic transient receptor potential channel (TRPC), the vanilloid-like transient receptor potential channel (TRPV), the ankyrin-rich transient receptor potential channel (TRPA), the transient receptor potential channel of the polycystin group (TRPP), and the transient receptor potential channel of the mucolipin group (TRPML) constitute the superfamily of TRP channels [93]. The ion pore of TRP channels is formed by a homo- or heterotetrameric complex of four TRP channel proteins. The multimerization described so far showed that heteromerization is possible in the limited range of closely related TRP channels [94,95,96]. TRPM proteins assemble into ion conducting channels that respond to a variety of stimuli including temperature, osmolarity, various chemical signals, change in membrane voltage, oxidative stress, and intracellular calcium [97]. Studies on TRP channel function are currently handicapped by the situation that the blockers available mostly represent pan TRP channel blocker hindering direct assignment of TRP channel-mediated functions [98,99].
TRPM3 is a polymodal ion channel activated by a variety of different stimuli like hypotonicity [100], sphingolipids [101], steroids [6,102], nifedipine [6], and heat [103]. The steroid-dependent modulation of TRPM3 is complex. Pregnenolone sulfate, pregnenolone and epipregnanolone sulfate stimulate TRPM3 activity whereas progesterone inhibits PregS-stimulated TRPM2 activity [6,102,104]. The concentration-response curves of PregS and pregnenolone reveal comparable potency of both steroids (Table 1), whereas both activating ligands showed dramatic differences in efficacy. The intrinsic activity of PregS is more than ten-fold higher than the intrinsic activity of pregnenolone [6]. With respect to the hydrophilicity of the sulfate moiety, it is interesting that TRPM3 is activated selectively by extracellular application of PregS, intracellular infusion of PregS via the patch pipette failed to induce TRPM3-mediated currents [6]. The activation of TRPM3 is remarkable as the temperature profile shows a continuous increase in TRPM3 activity in the range of 27 °C to 43 °C [103], which is contrast to TRPV1 showing increased activity beyond a threshold of 40 °C [105]. However, the most fascinating feature of the heat-dependent modulation of TRPM3 activity is the sensitization towards PregS [103]. At normal human body temperature, TRPM3 can be activated by PregS concentrations in the range of 100 to 500 nM corresponding to physiologic PregS serum concentrations (see above) [103]. Electrophysiological characterization revealed permeability of TRPM3 for calcium, sodium, magnesium, manganese and zinc ions [100,101,106,107]. Expression profile and subsequent functional analysis provides evidence for TRPM3 function in vascular smooth muscle contraction [108], modulation of glucose-induced insulin release from pancreatic islets [6,109], detection of noxious heat in dorsal root ganglia [103], and activation of oligodendrocytes and neurons in developing brains [91,110].
TRPM1, the founding member of the TRPM channel subfamily was discovered in a screen looking for transcriptional regulation in melanoma [92,111]. The absence of TRPM1 in malign melanoma guided the authors to name the TRP channel identified melastatin. Difficulties in cDNA cloning and expression as well as occurrence of many splice variants hampered the characterization of TRPM1 [112]. The protein was mainly studied in the context of melanocytes [113] until its role in mammalian phototransduction was discovered [114]. In electroretinograms (ERG) of TRPM1−/− mice the so-called b-wave, the measure for ON bipolar cells was missing [114]. Immunohistochemistry revealed expression of TRPM1 in rod ON bipolar cells and initiated a series of studies characterizing TRPM1 in the context of glutamate-induced signal transduction comprising metabotropic glutamic acid receptor (mGluR6), heteromeric Go proteins, and RGS proteins [115,116,117,118,119,120]. As TRPM1 like TRPM3 is activated by PregS [89] it is interesting to note the ability of retina for neurosteroidogenesis, which has been shown in rat retina [121]. Furthermore, it is possible that the neurotoxic effects in retina induced by PregS [122,123,124] are mediated by TRPM1-mediated calcium entry.
6. Conclusions
The aspects summarized here illustrated the versatile features of PregS from pregnenolone metabolite to source of pregnenolone as mother of steroids in subsequent steroid synthesis pathways to modulatory impact on ion channels. The latter is still matter of debate in the cases of GABAA channel and NMDA receptor activation. The nanomolar concentrations found in most tissues and situations on one hand and the micromolar concentrations used in the experiments on the other are critically discussed with respect to whether or not the experiments will mimic physiologic relevant pathways. Even the experiments showing effect of PregS in nanomolar concentrations in the presence of NMDA receptor blocker represent not really physiologic conditions. Currently, it could not be excluded that the modulation of GABAA channels and NMDA receptors by PregS show a different profile in experiments performed under physiologic temperatures as it has been shown for TRPM3. At all the unsolved questions in this debate, PregS is an interesting ligand and provide a broad field for future studies dissolving the physiologic role of PregS in the brain and peripheral organs.
Conflicts of Interest
The author declares no conflict of interest.
References
- Wehling, M.; Losel, R. Non-genomic steroid hormone effects: membrane or intracellular receptors? J. Steroid Biochem. Mol. Biol. 2006, 102, 180–183. [Google Scholar] [CrossRef]
- Sooksawate, T.; Simmonds, M.A. Influence of membrane cholesterol on modulation of the GABAA receptor by neuroactive steroids and other potentiators. Br. J. Pharmacol. 2001, 134, 1303–1311. [Google Scholar] [CrossRef]
- Dubrovsky, B. Neurosteroids, neuroactive steroids, and symptoms of affective disorders. Pharmacol. Biochem. Behav. 2006, 84, 644–655. [Google Scholar] [CrossRef]
- Mellon, S.H.; Griffin, L.D. Neurosteroids: biochemistry and clinical significance. Trends Endocrinol. Metab. 2002, 13, 35–43. [Google Scholar] [CrossRef]
- Zheng, P. Neuroactive steroid regulation of neurotransmitter release in the CNS: action, mechanism and possible significance. Prog. Neurobiol. 2009, 89, 134–152. [Google Scholar] [CrossRef]
- Wagner, T.F.; Loch, S.; Lambert, S.; Straub, I.; Mannebach, S.; Mathar, I.; Düfer, M.; Lis, A.; Flockerzi, V.; Philipp, S.E.; et al. Transient receptor potential M3 channels are ionotropic steroid receptors in pancreatic beta cells. Nat. Cell Biol. 2008, 10, 1421–1430. [Google Scholar] [CrossRef]
- Falany, C.N. Enzymology of human cytosolic sulfotransferases. FASEB J. 1997, 11, 206–216. [Google Scholar]
- Strott, C.A. Sulfonation and molecular action. Endocr. Rev. 2002, 23, 703–732. [Google Scholar] [CrossRef]
- Burel, D.; Li, J.H.; Do-Rego, J.L.; Wang, A.F.; Luu-The, V.; Pelletier, G.; Tillet, Y.; Taragnat, C.; Kwon, H.B.; Seong, J.Y.; Vaudry, H. Gonadotropin-releasing hormone stimulates the biosynthesis of pregnenolone sulfate and dehydroepiandrosterone sulfate in the hypothalamus. Endocrinology 2013, 154, 2114–2128. [Google Scholar] [CrossRef]
- Her, C.; Wood, T.C.; Eichler, E.E.; Mohrenweiser, H.W.; Ramagli, L.S.; Siciliano, M.J.; Weinshilboum, R.M. Human hydroxysteroid sulfotransferase SULT2B1: two enzymes encoded by a single chromosome 19 gene. Genomics 1998, 53, 284–295. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Maeda, N.; Okumura, K.; Tanaka, E.; Suzuki, T.; Miyasho, T.; Haeno, S.; Ueda, H.; Hoshi, N.; Yokota, H. Downregulation of cytochrome P450scc as an initial adverse effect of adult exposure to diethylstilbestrol on testicular steroidogenesis. Environ. Toxicol. 2013, in press. [Google Scholar]
- Parte, P.; Balasinor, N.; Gill-Sharma, M.K.; Maitra, A.; Juneja, H.S. Temporal effect of tamoxifen on cytochrome P450 side chain cleavage gene expression and steroid concentration in adult male rats. J. Steroid Biochem. Mol. Biol. 2002, 82, 349–358. [Google Scholar] [CrossRef]
- Wu, L.; Yan, J.; Qu, S.C.; Feng, Y.Q.; Jiang, X.L. Abnormal regulation for progesterone production in placenta with prenatal cocaine exposure in rats. Placenta 2012, 33, 977–981. [Google Scholar] [CrossRef]
- Kohjitani, A.; Fuda, H.; Hanyu, O.; Strott, C.A. Regulation of SULT2B1a (pregnenolone sulfotransferase) expression in rat C6 glioma cells: relevance of AMPA receptor-mediated NO signaling. Neurosci. Lett. 2008, 430, 75–80. [Google Scholar] [CrossRef]
- Kauffman, F.C. Sulfonation in pharmacology and toxicology. Drug Metab. Rev. 2004, 36, 823–843. [Google Scholar] [CrossRef]
- Geyer, J.; Wilke, T.; Petzinger, E. The solute carrier family SLC10: more than a family of bile acid transporters regarding function and phylogenetic relationships. Naunyn Schmiedebergs Arch. Pharmacol. 2006, 372, 413–431. [Google Scholar] [CrossRef]
- Hagenbuch, B.; Meier, P.J. Organic anion transporting polypeptides of the OATP/SLC21 family: phylogenetic classification as OATP/SLCO superfamily, new nomenclature and molecular/functional properties. Pflügers Arch. 2004, 447, 653–665. [Google Scholar] [CrossRef]
- Ghosh, D. Human sulfatases: a structural perspective to catalysis. Cell. Mol. Life Sci. 2007, 64, 2013–2022. [Google Scholar] [CrossRef]
- Kriz, L.; Bicikova, M.; Hampl, R. Roles of steroid sulfatase in brain and other tissues. Physiol. Res. 2008, 57, 657–668. [Google Scholar]
- Reed, M.J.; Purohit, A.; Woo, L.W.; Newman, S.P.; Potter, B.V. Steroid sulfatase: molecular biology, regulation, and inhibition. Endocr. Rev. 2005, 26, 171–202. [Google Scholar]
- Hernandez-Martin, A.; Gonzalez-Sarmiento, R.; De Unamuno, P. X-linked ichthyosis: an update. Br. J. Dermatol. 1999, 141, 617–627. [Google Scholar] [CrossRef]
- Chibbar, R.; Mitchell, B.F. Steroid sulfohydrolase in human chorion and decidua: studies using pregnenolone sulfate and dehydroepiandrosterone sulfate as substrate. J. Clin. Endocrinol. Metab. 1990, 70, 1693–1701. [Google Scholar] [CrossRef]
- Kauffman, F.C.; Whittaker, M.; Anundi, I.; Thurman, R.G. Futile cycling of a sulfate conjugate by isolated hepatocytes. Mol. Pharmacol. 1991, 39, 414–420. [Google Scholar]
- Payne, A.H.; Jaffe, R.B. Androgen formation from pregnenolone sulfate by the human fetal ovary. J. Clin. Endocrinol. Metab. 1974, 39, 300–304. [Google Scholar] [CrossRef]
- Stone, B.A.; Seamark, R.F.; Kelly, R.W.; Deam, S. Production of steroids and release of prostaglandins by spherical pig blastocysts in vitro. Aust. J. Biol. Sci. 1986, 39, 283–293. [Google Scholar]
- de Peretti, E.; Mappus, E. Pattern of plasma pregnenolone sulfate levels in humans from birth to adulthood. J. Clin. Endocrinol. Metab. 1983, 57, 550–556. [Google Scholar] [CrossRef]
- Bicikova, M.; Klak, J.; Hill, M.; Zizka, Z.; Hampl, R.; Calda, P. Two neuroactive steroids in midpregnancy as measured in maternal and fetal sera and in amniotic fluid. Steroids 2002, 67, 399–402. [Google Scholar] [CrossRef]
- Chang, R.J.; Buster, J.E.; Blakely, J.L.; Okada, D.M.; Hobel, C.J.; Abraham, G.E.; Marshall, J.R. Simultaneous comparison of Δ5-3β-hydroxysteroid levels in the fetoplacental circulation of normal pregnancy in labor and not in labor. J. Clin. Endocrinol. Metab. 1976, 42, 744–751. [Google Scholar] [CrossRef]
- Conrad, S.H.; Pion, R.J.; Kitchin, J.D., 3rd. Pregnenolone sulfate in human pregnancy plasma. J. Clin. Endocrinol. Metab. 1967, 27, 114–119. [Google Scholar] [CrossRef]
- Hill, M.; Parizek, A.; Klak, J.; Hampl, R.; Sulcova, J.; Havlikova, H.; Lapcik, O.; Bicikova, M.; Fait, T.; Kancheva, R.; et al. Neuroactive steroids, their precursors and polar conjugates during parturition and postpartum in maternal and umbilical blood: 3.3β-hydroxy-5-ene steroids. J. Steroid Biochem. Mol. Biol. 2002, 82, 241–250. [Google Scholar] [CrossRef]
- Klak, J.; Hill, M.; Parizek, A.; Havlikova, H.; Bicikova, M.; Hampl, R.; Fait, T.; Sulcova, J.; Pouzar, V.; Kancheva, R.; Starka, L. Pregnanolone isomers, pregnenolone and their polar conjugates around parturition. Physiol. Res. 2003, 52, 211–221. [Google Scholar]
- Mathur, R.S.; Landgrebe, S.; Moody, L.O.; Powell, S.; Williamson, H.O. Plasma steroid concentrations in maternal and umbilical circulation after spontaneous onset of labor. J. Clin. Endocrinol. Metab. 1980, 51, 1235–1238. [Google Scholar] [CrossRef]
- Norman, R.J.; Deppe, W.M.; Joubert, S.M.; Marivate, M. Umbilical artery concentrations of androstenedione increased in early labour in the leading twin fetus. Br. J. Obstet. Gynaecol. 1984, 91, 776–780. [Google Scholar] [CrossRef]
- Scommegna, A.; Burd, L.; Bieniarz, J. Progesterone and pregnenolone sulfate in pregnancy plasma. Am. J. Obstet. Gynecol. 1972, 113, 60–65. [Google Scholar]
- Nathanielsz, P.W.; Elsner, C.; Magyar, D.; Fridshal, D.; Freeman, A.; Buster, J.E. Time trend analysis of plasma unconjugated and sulfoconjugated estrone and 3β-Δ5-steroids in fetal and maternal sheep plasma in relation to spontaneous parturition at term. Endocrinology 1982, 110, 1402–1407. [Google Scholar] [CrossRef]
- McKay, S.A.; Jenkin, G.; Thorburn, G.D. Peripheral plasma concentrations of pregnenolone sulphate, pregnenolone, progesterone and 20α-hydroxy-4-pregnen-3-one in ewes throughout the oestrous cycle. J. Endocrinol. 1987, 113, 231–237. [Google Scholar] [CrossRef]
- Wang, M.; Seippel, L.; Purdy, R.H.; Backstrom, T. Relationship between symptom severity and steroid variation in women with premenstrual syndrome: study on serum pregnenolone, pregnenolone sulfate, 5α-pregnane-3,20-dione and 3α-hydroxy-5α-pregnan-20-one. J. Clin. Endocrinol. Metab. 1996, 81, 1076–1082. [Google Scholar] [CrossRef]
- de la Torre, B.; Hedman, M.; Noren, S.; Sjoberg, B. Blood and tissue steroid levels and their interrelationship in men with pathological conditions of the reproductive organs. Int. J. Androl. 1986, 9, 241–249. [Google Scholar] [CrossRef]
- Leinonen, P.; Ruokonen, A.; Kontturi, M.; Vihko, R. Effects of estrogen treatment on human testicular unconjugated steroid and steroid sulfate production in vivo. J. Clin. Endocrinol. Metab. 1981, 53, 569–573. [Google Scholar] [CrossRef]
- Ruokonen, A.; Lukkarinen, O.; Vihko, R. Secretion of steroid sulfates from human testis and their response to a single intramuscular injection of 5000 IU hCG. J. Steroid Biochem. 1981, 14, 1357–1360. [Google Scholar] [CrossRef]
- de Peretti, E.; Forest, M.G.; Loras, B.; Morel, Y.; David, M.; Francois, R.; Bertrand, J. Usefulness of plasma pregnenolone sulfate in testing pituitary-adrenal function in children. Acta Endocrinol. Suppl. (Copenh) 1986, 279, 259–263. [Google Scholar]
- Bicikova, M.; Szamel, I.; Hill, M.; Tallova, J.; Starka, L. Allopregnanolone, pregnenolone sulfate, and epitestosterone in breast cyst fluid. Steroids 2001, 66, 55–57. [Google Scholar] [CrossRef]
- Hedman, M.; Nilsson, E.; de la Torre, B. Low sulpho-conjugated steroid hormone levels in systemic lupus erythematosus (SLE). Clin. Exp. Rheumatol. 1989, 7, 583–588. [Google Scholar]
- Hedman, M.; Nilsson, E.; de la Torre, B. Low blood and synovial fluid levels of sulpho-conjugated steroids in rheumatoid arthritis. Clin. Exp. Rheumatol. 1992, 10, 25–30. [Google Scholar]
- Tagawa, N.; Tamanaka, J.; Fujinami, A.; Kobayashi, Y.; Takano, T.; Fukata, S.; Kuma, K.; Tada, H.; Amino, N. Serum dehydroepiandrosterone, dehydroepiandrosterone sulfate, and pregnenolone sulfate concentrations in patients with hyperthyroidism and hypothyroidism. Clin. Chem. 2000, 46, 523–528. [Google Scholar]
- Tallova, J.; Tomandl, J.; Bicikova, M.; Simickova, M. Homocysteine in breast cyst fluid. Eur. J. Clin. Invest. 2001, 31, 623–627. [Google Scholar] [CrossRef]
- Schumacher, M.; Liere, P.; Akwa, Y.; Rajkowski, K.; Griffiths, W.; Bodin, K.; Sjovall, J.; Baulieu, E.E. Pregnenolone sulfate in the brain: a controversial neurosteroid. Neurochem. Int. 2008, 52, 522–540. [Google Scholar] [CrossRef]
- Weill-Engerer, S.; David, J.P.; Sazdovitch, V.; Liere, P.; Eychenne, B.; Pianos, A.; Schumacher, M.; Delacourte, A.; Baulieu, E.E.; Akwa, Y. Neurosteroid quantification in human brain regions: comparison between Alzheimer's and nondemented patients. J. Clin. Endocrinol. Metab. 2002, 87, 5138–5143. [Google Scholar] [CrossRef]
- Corpechot, C.; Synguelakis, M.; Talha, S.; Axelson, M.; Sjovall, J.; Vihko, R.; Baulieu, E.E.; Robel, P. Pregnenolone and its sulfate ester in the rat brain. Brain Res. 1983, 270, 119–125. [Google Scholar] [CrossRef]
- Jo, D.H.; Abdallah, M.A.; Young, J.; Baulieu, E.E.; Robel, P. Pregnenolone, dehydroepiandrosterone, and their sulfate and fatty acid esters in the rat brain. Steroids 1989, 54, 287–297. [Google Scholar] [CrossRef]
- Caldeira, J.C.; Wu, Y.; Mameli, M.; Purdy, R.H.; Li, P.K.; Akwa, Y.; Savage, D.D.; Engen, J.R.; Valenzuela, C.F. Fetal alcohol exposure alters neurosteroid levels in the developing rat brain. J. Neurochem. 2004, 90, 1530–1539. [Google Scholar] [CrossRef]
- Xu, B.; Yang, R.; Chang, F.; Chen, L.; Xie, G.; Sokabe, M.; Chen, L. Neurosteroid PREGS protects neurite growth and survival of newborn neurons in the hippocampal dentate gyrus of APPswe/PS1dE9 mice. Curr. Alzheimer Res. 2012, 9, 361–372. [Google Scholar] [CrossRef]
- St-Pierre, M.V.; Hagenbuch, B.; Ugele, B.; Meier, P.J.; Stallmach, T. Characterization of an organic anion-transporting polypeptide (OATP-B) in human placenta. J. Clin. Endocrinol. Metab. 2002, 87, 1856–1863. [Google Scholar] [CrossRef]
- Grube, M.; Kock, K.; Karner, S.; Reuther, S.; Ritter, C.A.; Jedlitschky, G.; Kroemer, H.K. Modification of OATP2B1-mediated transport by steroid hormones. Mol. Pharmacol. 2006, 70, 1735–1741. [Google Scholar] [CrossRef]
- Geyer, J.; Doring, B.; Meerkamp, K.; Ugele, B.; Bakhiya, N.; Fernandes, C.F.; Godoy, J.R.; Glatt, H.; Petzinger, E. Cloning and functional characterization of human sodium-dependent organic anion transporter (SLC10A6). J. Biol. Chem. 2007, 282, 19728–19741. [Google Scholar] [CrossRef]
- Grosser, G.; Fietz, D.; Gunther, S.; Bakhaus, K.; Schweigmann, H.; Ugele, B.; Brehm, R.; Petzinger, E.; Bergmann, M.; Geyer, J. Cloning and functional characterization of the mouse sodium-dependent organic anion transporter Soat (Slc10a6). J. Steroid Biochem. Mol. Biol. 2013, 138C, 90–99. [Google Scholar]
- Fang, F.; Christian, W.V.; Gorman, S.G.; Cui, M.; Huang, J.; Tieu, K.; Ballatori, N. Neurosteroid transport by the organic solute transporter OSTα-OSTβ. J. Neurochem. 2010, 115, 220–233. [Google Scholar] [CrossRef]
- Kobayashi, T.; Washiyama, K.; Ikeda, K. Pregnenolone sulfate potentiates the inwardly rectifying K channel Kir2.3. PLoS One 2009, 4, e6311. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, L.; Wardwell-Swanson, J. Modulation of cloned human neuronal voltage-gated potassium channels (hKv1.1 and hKv2.1) by neurosteroids. Pflügers Arch. 1998, 437, 49–55. [Google Scholar] [CrossRef]
- Kudo, K.; Tachikawa, E.; Kashimoto, T. Inhibition by pregnenolone sulfate of nicotinic acetylcholine response in adrenal chromaffin cells. Eur. J. Pharmacol. 2002, 456, 19–27. [Google Scholar] [CrossRef]
- Horishita, T.; Ueno, S.; Yanagihara, N.; Sudo, Y.; Uezono, Y.; Okura, D.; Sata, T. Inhibition by pregnenolone sulphate, a metabolite of the neurosteroid pregnenolone, of voltage-gated sodium channels expressed in Xenopus oocytes. J. Pharmacol. Sci. 2012, 120, 54–58. [Google Scholar] [CrossRef]
- Wu, F.S.; Gibbs, T.T.; Farb, D.H. Pregnenolone sulfate: a positive allosteric modulator at the N-methyl-D-aspartate receptor. Mol. Pharmacol. 1991, 40, 333–336. [Google Scholar]
- Irwin, R.P.; Maragakis, N.J.; Rogawski, M.A.; Purdy, R.H.; Farb, D.H.; Paul, S.M. Pregnenolone sulfate augments NMDA receptor mediated increases in intracellular Ca2+ in cultured rat hippocampal neurons. Neurosci. Lett. 1992, 141, 30–34. [Google Scholar] [CrossRef]
- Majewska, M.D.; Bluet-Pajot, M.T.; Robel, P.; Baulieu, E.E. Pregnenolone sulfate antagonizes barbiturate-induced hypnosis. Pharmacol. Biochem. Behav. 1989, 33, 701–703. [Google Scholar] [CrossRef]
- Majewska, M.D.; Demirgoren, S.; London, E.D. Binding of pregnenolone sulfate to rat brain membranes suggests multiple sites of steroid action at the GABAA receptor. Eur. J. Pharmacol. 1990, 189, 307–315. [Google Scholar] [CrossRef]
- Majewska, M.D.; Mienville, J.M.; Vicini, S. Neurosteroid pregnenolone sulfate antagonizes electrophysiological responses to GABA in neurons. Neurosci. Lett. 1988, 90, 279–284. [Google Scholar] [CrossRef]
- Zaman, S.H.; Shingai, R.; Harvey, R.J.; Darlison, M.G.; Barnard, E.A. Effects of subunit types of the recombinant GABAA receptor on the response to a neurosteroid. Eur. J. Pharmacol. 1992, 225, 321–330. [Google Scholar] [CrossRef]
- Akk, G.; Li, P.; Manion, B.D.; Evers, A.S.; Steinbach, J.H. Ethanol modulates the interaction of the endogenous neurosteroid allopregnanolone with the α1β2γ2L GABAA receptor. Mol. Pharmacol. 2007, 71, 461–472. [Google Scholar]
- Twede, V.; Tartaglia, A.L.; Covey, D.F.; Bamber, B.A. The neurosteroids dehydroepiandrosterone sulfate and pregnenolone sulfate inhibit the UNC-49 GABA receptor through a common set of residues. Mol. Pharmacol. 2007, 72, 1322–1329. [Google Scholar] [CrossRef]
- Baker, C.; Sturt, B.L.; Bamber, B.A. Multiple roles for the first transmembrane domain of GABAA receptor subunits in neurosteroid modulation and spontaneous channel activity. Neurosci. Lett. 2010, 473, 242–247. [Google Scholar] [CrossRef]
- Elfverson, M.; Linde, A.M.; Le Greves, P.; Zhou, Q.; Nyberg, F.; Johansson, T. Neurosteroids allosterically modulate the ion pore of the NMDA receptor consisting of NR1/NR2B but not NR1/NR2A. Biochem. Biophys. Res. Commun. 2008, 372, 305–308. [Google Scholar] [CrossRef]
- Johansson, T.; Frandberg, P.A.; Nyberg, F.; Le Greves, P. Molecular mechanisms for nanomolar concentrations of neurosteroids at NR1/NR2B receptors. J. Pharmacol. Exp. Ther. 2008, 324, 759–768. [Google Scholar]
- Alexander, S.P.; Mathie, A.; Peters, J.A. Guide to Receptors and Channels (GRAC), 5th edition. Br. J. Pharmacol. 2011, 164 (Suppl. 1), S1–324. [Google Scholar] [CrossRef]
- Ceccon, M.; Rumbaugh, G.; Vicini, S. Distinct effect of pregnenolone sulfate on NMDA receptor subtypes. Neuropharmacology 2001, 40, 491–500. [Google Scholar] [CrossRef]
- Horak, M.; Vlcek, K.; Chodounska, H.; Vyklicky, L., Jr. Subtype-dependence of N-methyl-D-aspartate receptor modulation by pregnenolone sulfate. Neuroscience 2006, 137, 93–102. [Google Scholar] [CrossRef]
- Malayev, A.; Gibbs, T.T.; Farb, D.H. Inhibition of the NMDA response by pregnenolone sulphate reveals subtype selective modulation of NMDA receptors by sulphated steroids. Br. J. Pharmacol. 2002, 135, 901–909. [Google Scholar] [CrossRef]
- Petrovic, M.; Sedlacek, M.; Cais, O.; Horak, M.; Chodounska, H.; Vyklicky, L., Jr. Pregnenolone sulfate modulation of N-methyl-D-aspartate receptors is phosphorylation dependent. Neuroscience 2009, 160, 616–628. [Google Scholar] [CrossRef]
- Kashiwagi, K.; Fukuchi, J.; Chao, J.; Igarashi, K.; Williams, K. An aspartate residue in the extracellular loop of the N-methyl-D-aspartate receptor controls sensitivity to spermine and protons. Mol. Pharmacol. 1996, 49, 1131–1141. [Google Scholar]
- Low, C.M.; Lyuboslavsky, P.; French, A.; Le, P.; Wyatte, K.; Thiel, W.H.; Marchan, E.M.; Igarashi, K.; Kashiwagi, K.; Gernert, K.; Williams, K.; Traynelis, S.F.; Zheng, F. Molecular determinants of proton-sensitive N-methyl-D-aspartate receptor gating. Mol. Pharmacol. 2003, 63, 1212–1222. [Google Scholar] [CrossRef]
- Jang, M.K.; Mierke, D.F.; Russek, S.J.; Farb, D.H. A steroid modulatory domain on NR2B controls N-methyl-D-aspartate receptor proton sensitivity. Proc. Natl. Acad. Sci. USA 2004, 101, 8198–8203. [Google Scholar] [CrossRef]
- Kostakis, E.; Smith, C.; Jang, M.K.; Martin, S.C.; Richards, K.G.; Russek, S.J.; Gibbs, T.T.; Farb, D.H. The Neuroactive Steroid Pregnenolone Sulfate Stimulates Trafficking of Functional N-Methyl D-Aspartate Receptors to the Cell Surface via a Noncanonical, G Protein, and Ca2+-Dependent Mechanism. Mol. Pharmacol. 2013, 84, 261–274. [Google Scholar] [CrossRef]
- Valenzuela, C.F.; Partridge, L.D.; Mameli, M.; Meyer, D.A. Modulation of glutamatergic transmission by sulfated steroids: role in fetal alcohol spectrum disorder. Brain Res. Rev. 2008, 57, 506–519. [Google Scholar] [CrossRef]
- Chen, L.; Miyamoto, Y.; Furuya, K.; Mori, N.; Sokabe, M. PREGS induces LTP in the hippocampal dentate gyrus of adult rats via the tyrosine phosphorylation of NR2B coupled to ERK/CREB [corrected] signaling. J. Neurophysiol. 2007, 98, 1538–1548. [Google Scholar] [CrossRef]
- Sabeti, J.; Nelson, T.E.; Purdy, R.H.; Gruol, D.L. Steroid pregnenolone sulfate enhances NMDA-receptor-independent long-term potentiation at hippocampal CA1 synapses: role for L-type calcium channels and sigma-receptors. Hippocampus 2007, 17, 349–369. [Google Scholar] [CrossRef]
- Sliwinski, A.; Monnet, F.P.; Schumacher, M.; Morin-Surun, M.P. Pregnenolone sulfate enhances long-term potentiation in CA1 in rat hippocampus slices through the modulation of N-methyl-D-aspartate receptors. J. Neurosci. Res. 2004, 78, 691–701. [Google Scholar] [CrossRef]
- Whittaker, M.T.; Gibbs, T.T.; Farb, D.H. Pregnenolone sulfate induces NMDA receptor dependent release of dopamine from synaptic terminals in the striatum. J. Neurochem. 2008, 107, 510–521. [Google Scholar] [CrossRef]
- Johansson, T.; Elfverson, M.; Zhou, Q.; Nyberg, F. Allosteric modulation of the NMDA receptor by neurosteroids in rat brain and the impact of long term morphine administration. Biochem. Biophys. Res. Commun. 2010, 401, 504–508. [Google Scholar] [CrossRef]
- Lambert, S.; Drews, A.; Rizun, O.; Wagner, T.F.; Lis, A.; Mannebach, S.; Plant, S.; Portz, M.; Meissner, M.; Philipp, S.E.; Oberwinkler, J. Transient receptor potential melastatin 1 (TRPM1) is an ion-conducting plasma membrane channel inhibited by zinc ions. J. Biol. Chem. 2011, 286, 12221–12233. [Google Scholar] [CrossRef]
- Zamudio-Bulcock, P.A.; Valenzuela, C.F. Pregnenolone sulfate increases glutamate release at neonatal climbing fiber-to-Purkinje cell synapses. Neuroscience 2011, 175, 24–36. [Google Scholar] [CrossRef]
- Zamudio-Bulcock, P.A.; Everett, J.; Harteneck, C.; Valenzuela, C.F. Activation of steroid-sensitive TRPM3 channels potentiates glutamatergic transmission at cerebellar Purkinje neurons from developing rats. J. Neurochem. 2011, 119, 474–485. [Google Scholar] [CrossRef]
- Duncan, L.M.; Deeds, J.; Hunter, J.; Shao, J.; Holmgren, L.M.; Woolf, E.A.; Tepper, R.I.; Shyjan, A.W. Down-regulation of the novel gene melastatin correlates with potential for melanoma metastasis. Cancer Res. 1998, 58, 1515–1520. [Google Scholar]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Bruford, E.A.; Caterina, M.J.; Clapham, D.E.; Harteneck, C.; Heller, S.; Julius, D.; et al. A unified nomenclature for the superfamily of TRP cation channels. Mol. Cell 2002, 9, 229–231. [Google Scholar] [CrossRef]
- Hellwig, N.; Albrecht, N.; Harteneck, C.; Schultz, G.; Schaefer, M. Homo- and heteromeric assembly of TRPV channel subunits. J. Cell Sci. 2005, 118, 917–928. [Google Scholar] [CrossRef]
- Hofmann, T.; Schaefer, M.; Schultz, G.; Gudermann, T. Subunit composition of mammalian transient receptor potential channels in living cells. Proc. Natl. Acad. Sci. USA 2002, 99, 7461–7466. [Google Scholar] [CrossRef]
- Hoenderop, J.G.; Voets, T.; Hoefs, S.; Weidema, F.; Prenen, J.; Nilius, B.; Bindels, R.J. Homo- and heterotetrameric architecture of the epithelial Ca2+ channels TRPV5 and TRPV6. EMBO J. 2003, 22, 776–785. [Google Scholar] [CrossRef]
- Harteneck, C. Function and pharmacology of TRPM cation channels. Naunyn Schmiedebergs Arch. Pharmacol. 2005, 371, 307–314. [Google Scholar] [CrossRef]
- Harteneck, C.; Frenzel, H.; Kraft, R. N-(p-amylcinnamoyl)anthranilic acid (ACA): A phospholipase A2 inhibitor and TRP channel blocker. Cardiovasc. Drug Rev. 2007, 25, 61–75. [Google Scholar] [CrossRef]
- Harteneck, C.; Klose, C.; Krautwurst, D. Synthetic modulators of TRP channel activity. Adv. Exp. Med. Biol. 2011, 704, 87–106. [Google Scholar] [CrossRef]
- Grimm, C.; Kraft, R.; Sauerbruch, S.; Schultz, G.; Harteneck, C. Molecular and functional characterization of the melastatin-related cation channel TRPM3. J. Biol. Chem. 2003, 278, 21493–21501. [Google Scholar]
- Grimm, C.; Kraft, R.; Schultz, G.; Harteneck, C. Activation of the melastatin-related cation channel TRPM3 by D-erythro-sphingosine. Mol. Pharmacol. 2005, 67, 798–805. [Google Scholar]
- Majeed, Y.; Agarwal, A.K.; Naylor, J.; Seymour, V.A.; Jiang, S.; Muraki, K.; Fishwick, C.W.; Beech, D.J. Cis-isomerism and other chemical requirements of steroidal agonists and partial agonists acting at TRPM3 channels. Br. J. Pharmacol. 2010, 161, 430–441. [Google Scholar] [CrossRef]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef]
- Majeed, Y.; Tumova, S.; Green, B.L.; Seymour, V.A.; Woods, D.M.; Agarwal, A.K.; Naylor, J.; Jiang, S.; Picton, H.M.; Porter, K.E.; et al. Pregnenolone sulphate-independent inhibition of TRPM3 channels by progesterone. Cell Calcium 2012, 51, 1–11. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Oberwinkler, J.; Lis, A.; Giehl, K.M.; Flockerzi, V.; Philipp, S.E. Alternative splicing switches the divalent cation selectivity of TRPM3 channels. J. Biol. Chem. 2005, 280, 22540–22548. [Google Scholar]
- Wagner, T.F.; Drews, A.; Loch, S.; Mohr, F.; Philipp, S.E.; Lambert, S.; Oberwinkler, J. TRPM3 channels provide a regulated influx pathway for zinc in pancreatic beta cells. Pflügers Arch. 2010, 460, 755–765. [Google Scholar] [CrossRef]
- Naylor, J.; Li, J.; Milligan, C.J.; Zeng, F.; Sukumar, P.; Hou, B.; Sedo, A.; Yuldasheva, N.; Majeed, Y.; Beri, D.; et al. Pregnenolone sulphate- and cholesterol-regulated TRPM3 channels coupled to vascular smooth muscle secretion and contraction. Circ. Res. 2010, 106, 1507–1515. [Google Scholar]
- Klose, C.; Straub, I.; Riehle, M.; Ranta, F.; Krautwurst, D.; Ullrich, S.; Meyerhof, W.; Harteneck, C. Fenamates as TRP channel blockers: Mefenamic acid selectively blocks TRPM3. Br. J. Pharmacol. 2011, 162, 1757–1769. [Google Scholar] [CrossRef]
- Hoffmann, A.; Grimm, C.; Kraft, R.; Goldbaum, O.; Wrede, A.; Nolte, C.; Hanisch, U.K.; Richter-Landsberg, C.; Bruck, W.; Kettenmann, H.; et al. TRPM3 is expressed in sphingosine-responsive myelinating oligodendrocytes. J. Neurochem. 2010, 114, 654–665. [Google Scholar] [CrossRef]
- Hunter, J.J.; Shao, J.; Smutko, J.S.; Dussault, B.J.; Nagle, D.L.; Woolf, E.A.; Holmgren, L.M.; Moore, K.J.; Shyjan, A.W. Chromosomal localization and genomic characterization of the mouse melastatin gene (Mlsn1). Genomics 1998, 54, 116–123. [Google Scholar] [CrossRef]
- Oancea, E.; Vriens, J.; Brauchi, S.; Jun, J.; Splawski, I.; Clapham, D.E. TRPM1 forms ion channels associated with melanin content in melanocytes. Sci. Signal. 2009, 2, ra21. [Google Scholar] [CrossRef]
- Guo, H.; Carlson, J.A.; Slominski, A. Role of TRPM in melanocytes and melanoma. Exp. Dermatol. 2012, 21, 650–654. [Google Scholar] [CrossRef]
- Shen, Y.; Heimel, J.A.; Kamermans, M.; Peachey, N.S.; Gregg, R.G.; Nawy, S. A transient receptor potential-like channel mediates synaptic transmission in rod bipolar cells. J. Neurosci. 2009, 29, 6088–6093. [Google Scholar] [CrossRef]
- Cao, Y.; Pahlberg, J.; Sarria, I.; Kamasawa, N.; Sampath, A.P.; Martemyanov, K.A. Regulators of G protein signaling RGS7 and RGS11 determine the onset of the light response in ON bipolar neurons. Proc. Natl. Acad. Sci. USA 2012, 109, 7905–7910. [Google Scholar]
- Koike, C.; Obara, T.; Uriu, Y.; Numata, T.; Sanuki, R.; Miyata, K.; Koyasu, T.; Ueno, S.; Funabiki, K.; Tani, A.; et al. TRPM1 is a component of the retinal ON bipolar cell transduction channel in the mGluR6 cascade. Proc. Natl. Acad. Sci. USA 2010, 107, 332–337. [Google Scholar] [CrossRef]
- Morgans, C.W.; Zhang, J.; Jeffrey, B.G.; Nelson, S.M.; Burke, N.S.; Duvoisin, R.M.; Brown, R.L. TRPM1 is required for the depolarizing light response in retinal ON-bipolar cells. Proc. Natl. Acad. Sci. USA 2009, 106, 19174–19178. [Google Scholar]
- Shen, Y.; Rampino, M.A.; Carroll, R.C.; Nawy, S. G-protein-mediated inhibition of the Trp channel TRPM1 requires the Gβγ dimer. Proc. Natl. Acad. Sci. USA 2012, 109, 8752–8757. [Google Scholar] [CrossRef]
- Shim, H.; Wang, C.T.; Chen, Y.L.; Chau, V.Q.; Fu, K.G.; Yang, J.; McQuiston, A.R.; Fisher, R.A.; Chen, C.K. Defective retinal depolarizing bipolar cells in regulators of G protein signaling (RGS) 7 and 11 double null mice. J. Bio.l Chem. 2012, 287, 14873–14879. [Google Scholar] [CrossRef]
- Xu, Y.; Dhingra, A.; Fina, M.E.; Koike, C.; Furukawa, T.; Vardi, N. mGluR6 deletion renders the TRPM1 channel in retina inactive. J. Neurophysiol. 2012, 107, 948–957. [Google Scholar] [CrossRef]
- Guarneri, P.; Guarneri, R.; Cascio, C.; Pavasant, P.; Piccoli, F.; Papadopoulos, V. Neurosteroidogenesis in rat retinas. J. Neurochem. 1994, 63, 86–96. [Google Scholar]
- Cascio, C.; Guarneri, R.; Russo, D.; De Leo, G.; Guarneri, M.; Piccoli, F.; Guarneri, P. Pregnenolone sulfate, a naturally occurring excitotoxin involved in delayed retinal cell death. J. Neurochem. 2000, 74, 2380–2391. [Google Scholar]
- Cascio, C.; Guarneri, R.; Russo, D.; De Leo, G.; Guarneri, M.; Piccoli, F.; Guarneri, P. A caspase-3-dependent pathway is predominantly activated by the excitotoxin pregnenolone sulfate and requires early and late cytochrome c release and cell-specific caspase-2 activation in the retinal cell death. J. Neurochem. 2002, 83, 1358–1371. [Google Scholar] [CrossRef]
- Guarneri, P.; Russo, D.; Cascio, C.; De Leo, G.; Piccoli, T.; Sciuto, V.; Piccoli, F.; Guarneri, R. Pregnenolone sulfate modulates NMDA receptors, inducing and potentiating acute excitotoxicity in isolated retina. J. Neurosci. Res. 1998, 54, 787–797. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Harteneck, C. Pregnenolone Sulfate: From Steroid Metabolite to TRP Channel Ligand. Molecules 2013, 18, 12012-12028. https://doi.org/10.3390/molecules181012012
AMA Style
Harteneck C. Pregnenolone Sulfate: From Steroid Metabolite to TRP Channel Ligand. Molecules. 2013; 18(10):12012-12028. https://doi.org/10.3390/molecules181012012
Chicago/Turabian StyleHarteneck, Christian. 2013. "Pregnenolone Sulfate: From Steroid Metabolite to TRP Channel Ligand" Molecules 18, no. 10: 12012-12028. https://doi.org/10.3390/molecules181012012