Antioxidant Activity and Anti-Adipogenic Effects of Wild Herbs Mainly Cultivated in Korea


Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolic and Flavonoid Contents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Total phenolic contents (mg GAE/g) | Total flavonoid contents (mg RE/g) |
|---|---|---|
| Aster scaber extract | 183.5 ± 4.0 b | 103.9 ± 3.4 a |
| Ligularia fischeri extract | 215.8 ± 14.2 a | 86.9 ± 3.8 b |
| Kalopanax pictus extract | 75.4 ± 8.3 c | 30.5 ± 4.7 c |
| Codonopsis lanceolata extract | 14.9 ± 6.1 e | 3.8 ± 3.7 e |
| Cirsium setidens extract | 55.4 ± 7.3 d | 58.3 ± 5.3 d |
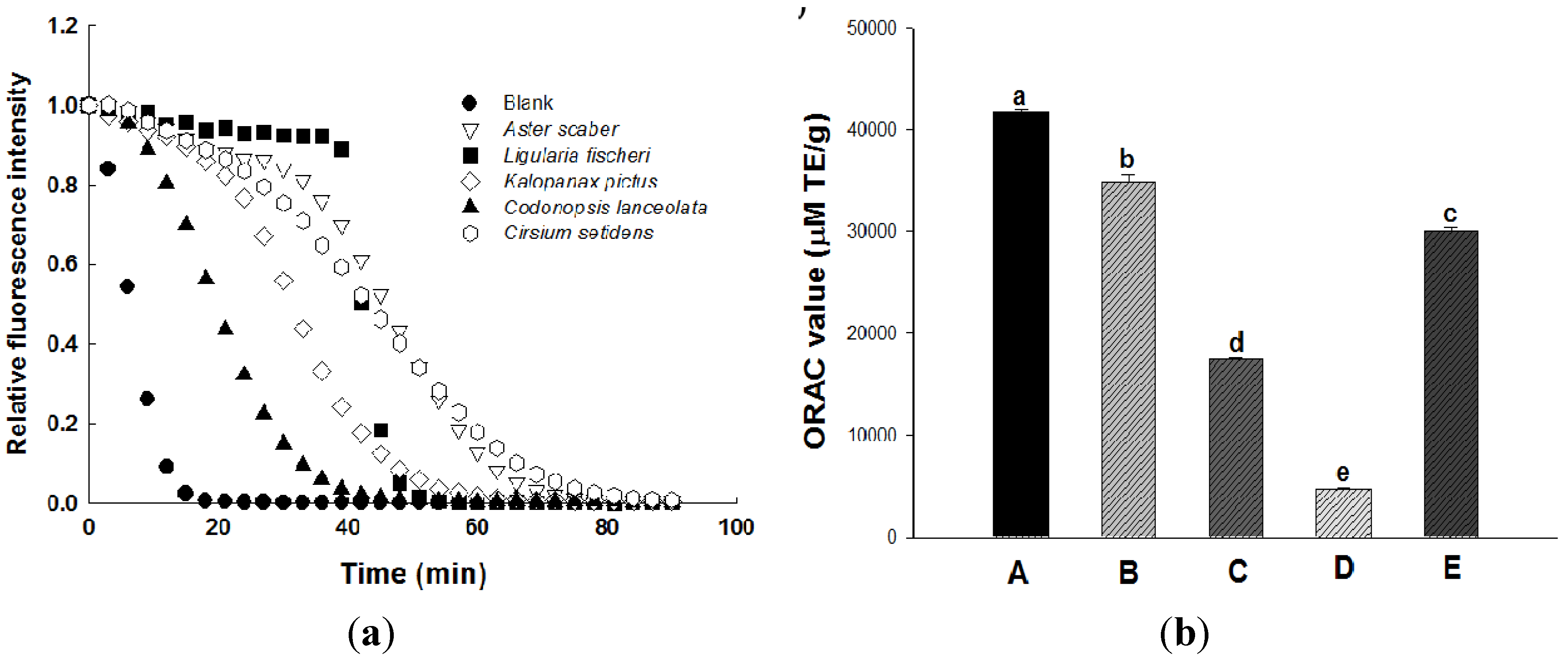
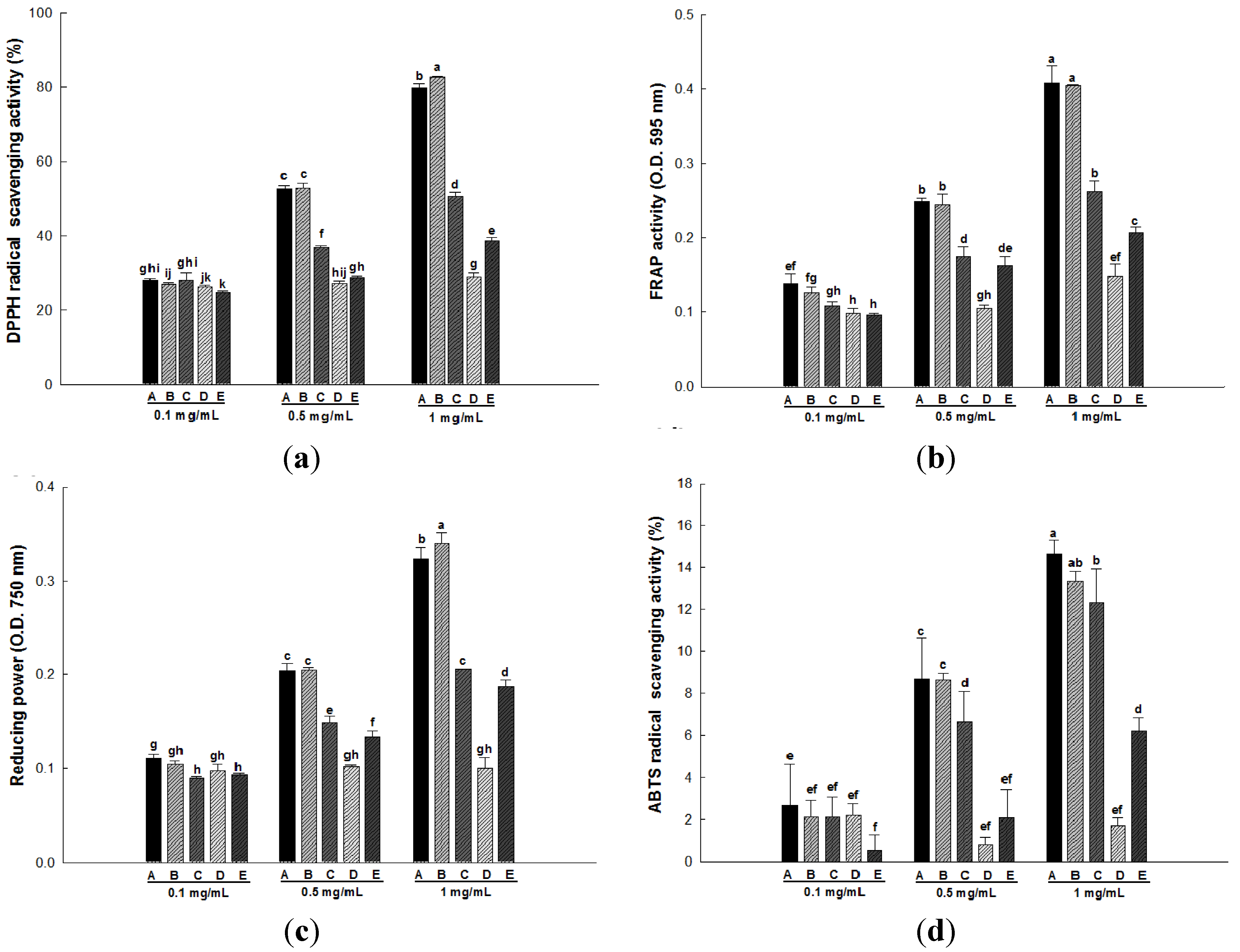
2.2. Antioxidant Activity
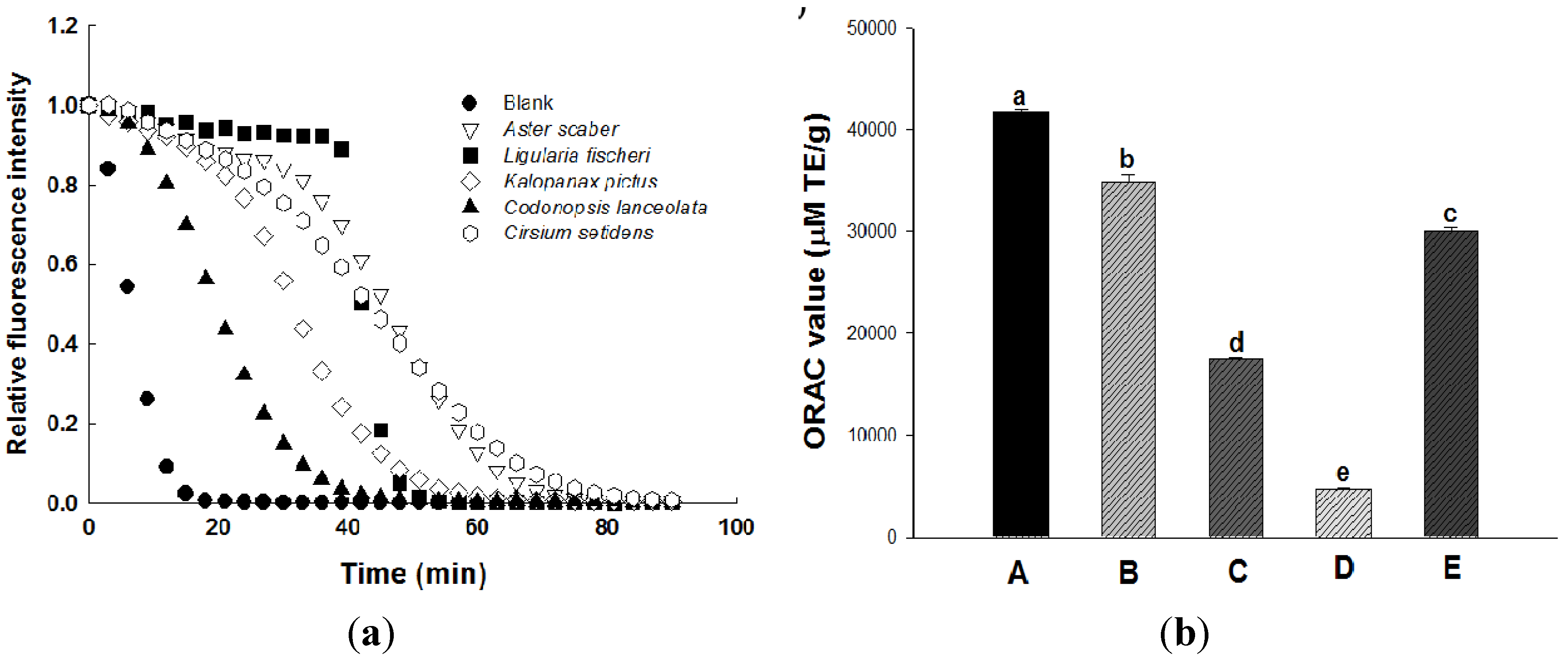
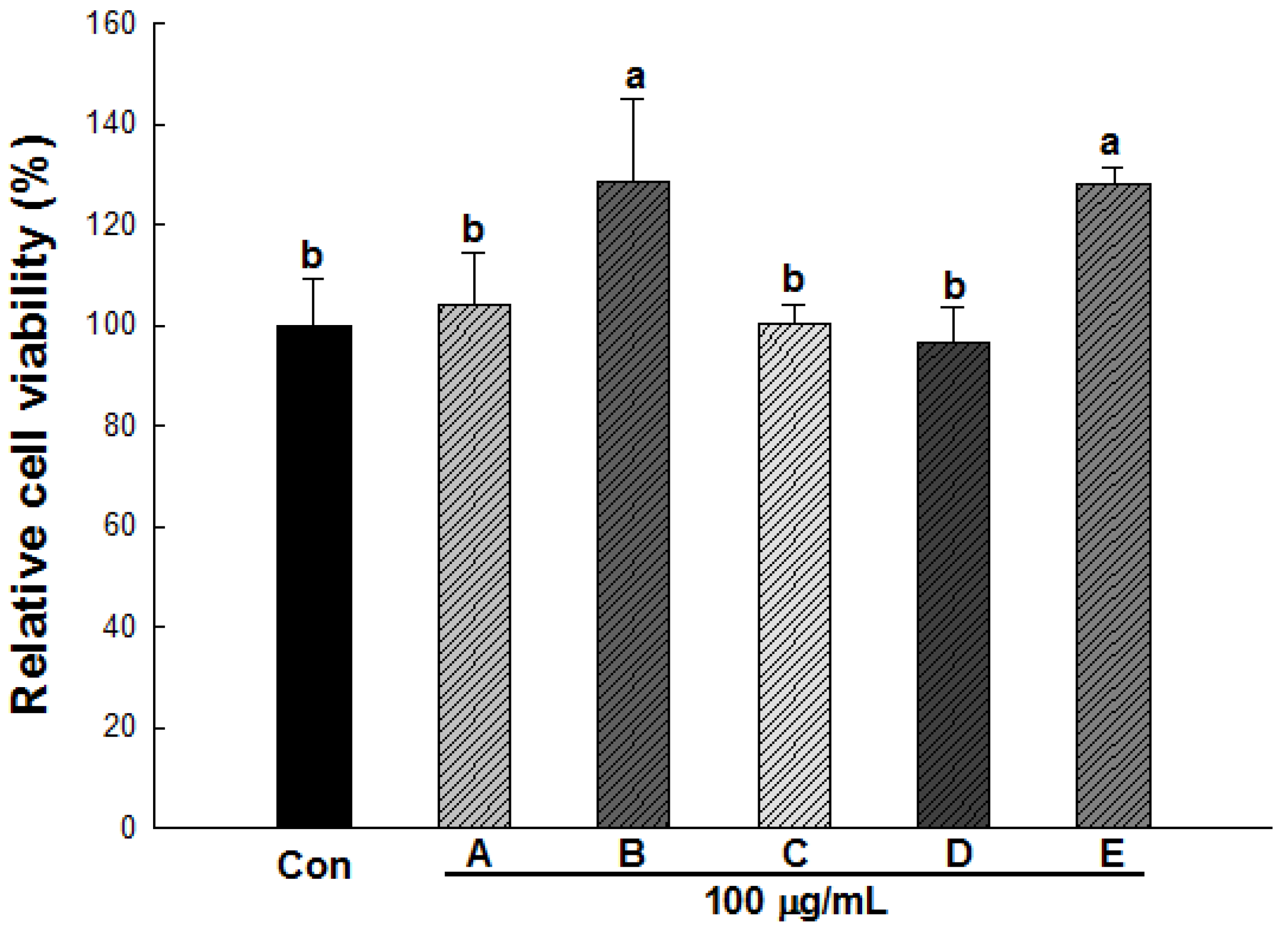
2.3. Effect of Wild Herb Extracts on the Cell Viability of 3T3-L1 Preadipocytes
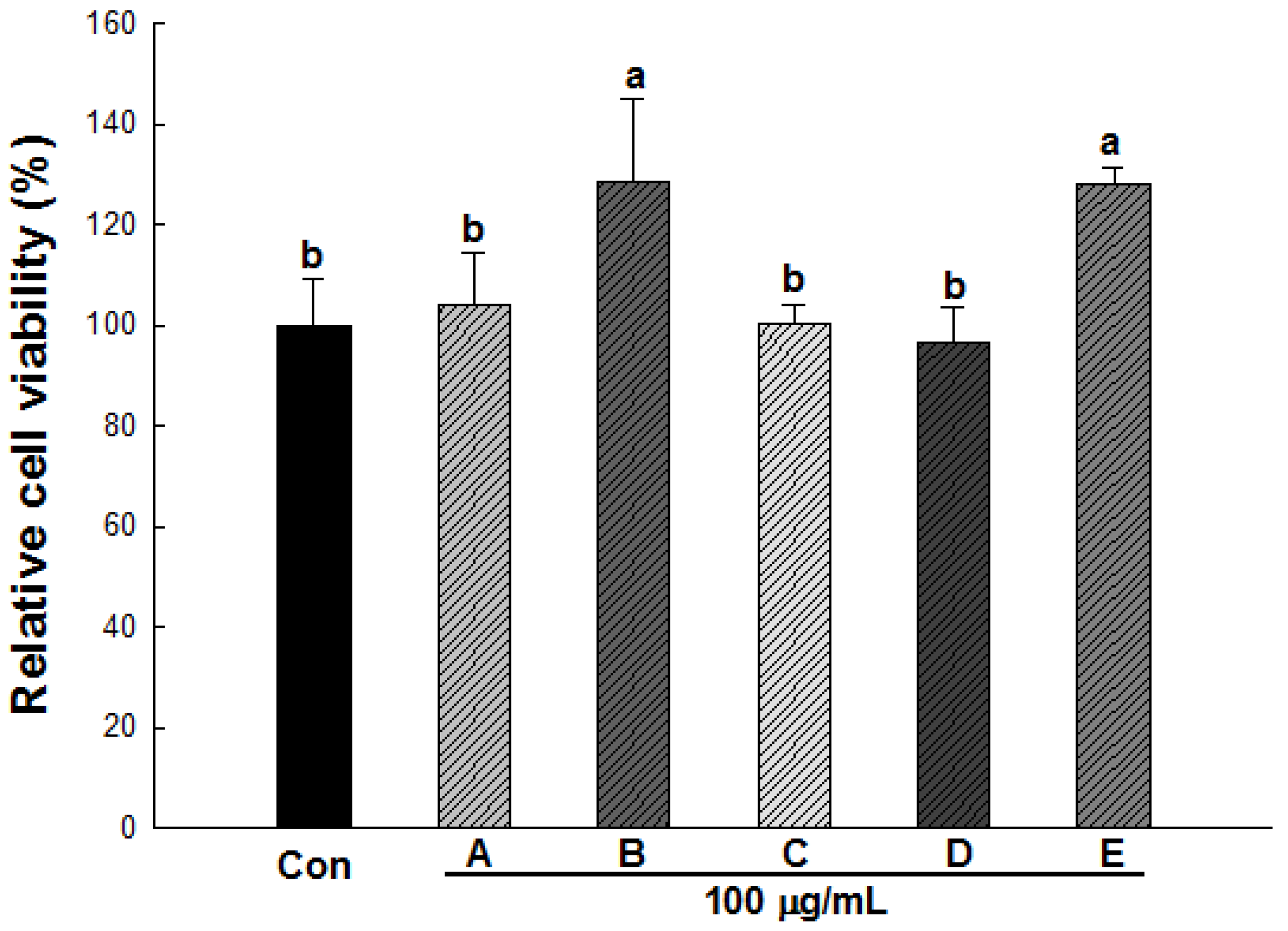
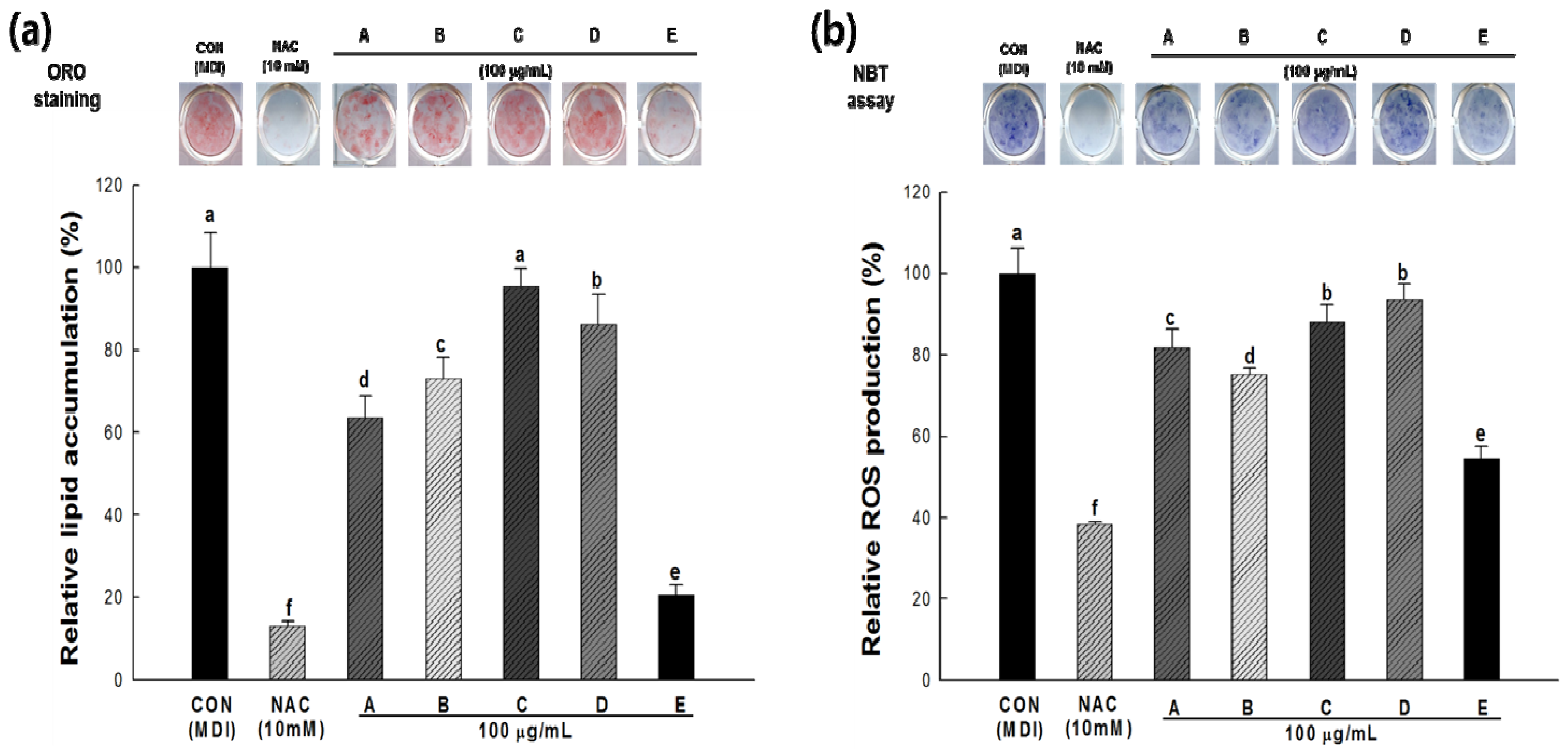
2.4. Lipid Accumulation and ROS Production in 3T3-L1 Cells
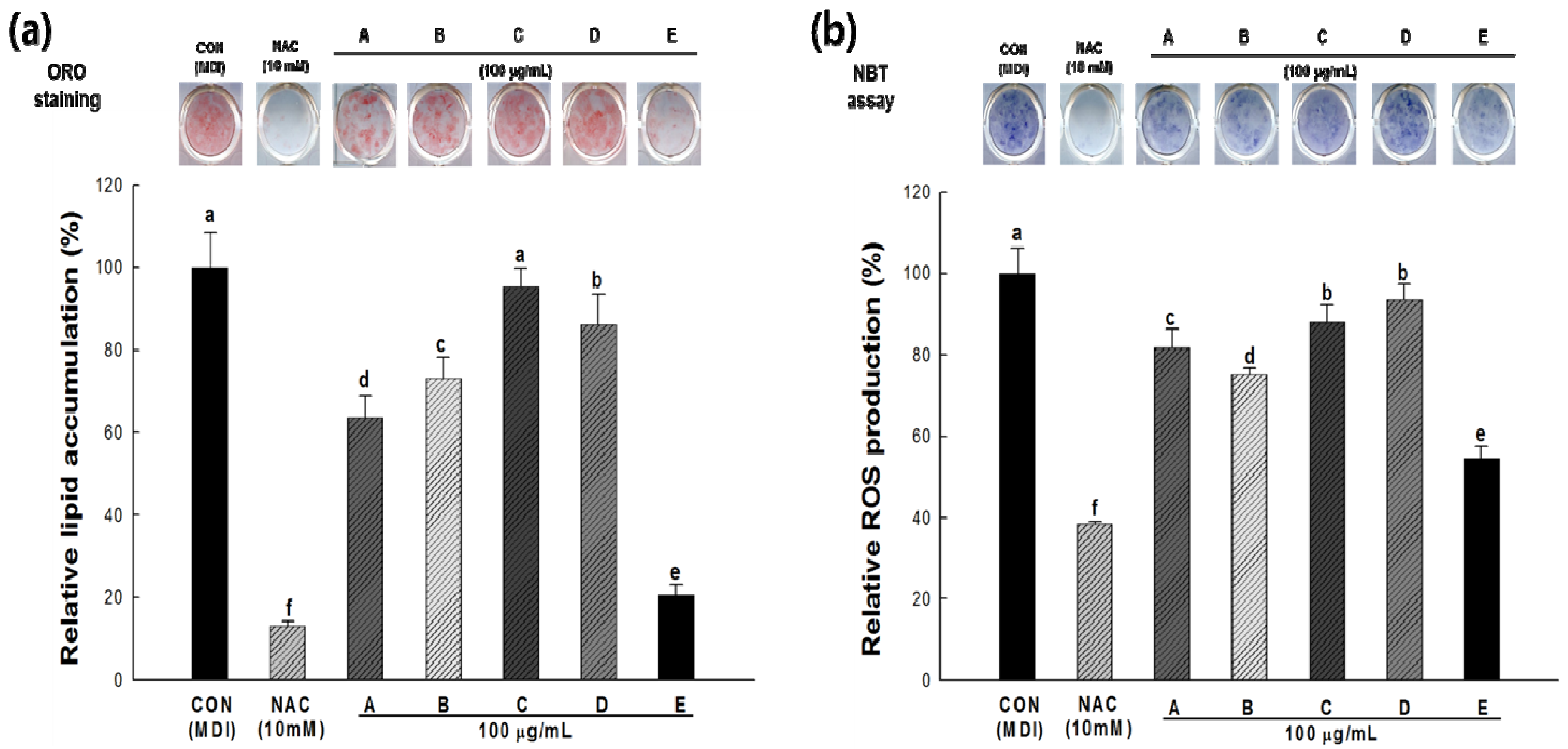
2.5. Correlation Coefficients (R) between Bioactive Compounds and Their Biological Activity
| TPC a | TFC b | DPPH | FRAP | Reducing power | ABTS | ORAC | Lipid accumulation | ROS production | |
|---|---|---|---|---|---|---|---|---|---|
| TPC | 1 | 0.890 *c | 0.991 ** | 0.984 ** | 0.985 ** | 0.847 | 0.825 | 0.017 | 0.108 |
| TFC | 1 | 0.882 * | 0.899 * | 0.930 * | 0.774 | 0.987 ** | 0.379 | 0.419 | |
| DPPH | 1 | 0.998 ** | 0.985 ** | 0.898 * | 0.816 | 0.082 | 0.035 | ||
| FRAP | 1 | 0.988 ** | 0.914 * | 0.840 | 0.053 | 0.056 | |||
| Reducing power | 1 | 0.901 * | 0.890 * | 0.070 | 0.0203 | ||||
| ABTS | 1 | 0.753 | 0.142 | 0.021 | |||||
| ORAC | 1 | 0.485 | 0.536 | ||||||
| Lipid accumulation | 1 | 0.935 * | |||||||
| ROS production | 1 |
3. Experimental
3.1. Materials
3.2. Total Phenolic and Flavonoid Contents
3.3. DPPH Radical Scavenging Assay
3.4. FRAP Assay
3.5. Reducing Power
3.6. ABTS Radical Scavenging Activity
3.7. ORAC Assay
3.8. Cell Culture
3.9. XTT Assay
3.10. NBT Assay
3.11. Oil Red O Staining
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ferreira, I.C.F.R.; Barros, L.; Abreu, R. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef]
- Cheeseman, K.H.; Slater, T.F. An introduction to free radical biochemistry. Br. Med. Bull. 1993, 49, 481–493. [Google Scholar]
- Gülçın, İ.; Oktay, M.; Kıreçcı, E.; Küfrevıoǧlu, Ö.İ. Screening of antioxidant and antimicrobial activities of anise (Pimpinella anisum L.) seed extracts. Food Chem. 2003, 83, 371–382. [Google Scholar] [CrossRef]
- Niki, E. Antioxidant capacity: Which capacity and how to assess it? J. Berry Res. 2011, 1, 169–176. [Google Scholar]
- Branen, A.L. Toxicology and biochemistry of butylated hydroxyanisole and butylated hydroxytoluene. J. Am. Oil Chem. Soc. 1975, 52, 59–63. [Google Scholar] [CrossRef]
- Ito, N.; Fukushima, S.; Hagiwara, A.; Shibata, M.; Ogiso, T. Carcinogenicity of butylated hydroxyanisole in F344 rats. J. Natl. Cancer Inst. 1983, 70, 343–352. [Google Scholar]
- Ajila, C.M.; Naidu, K.A.; Bhat, S.G.; Prasada Rao, U.J.S. Bioactive compounds and antioxidant potential of mango peel extract. Food Chem. 2007, 105, 982–988. [Google Scholar] [CrossRef]
- Ali, S.S.; Kasoju, N.; Luthra, A.; Singh, A.; Sharanabasava, H.; Sahu, A.; Bora, U. Indian medicinal herbs as sources of antioxidants. Food Res. Int. 2008, 41, 1–15. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Larson, R.A. The antioxidants of higher plants. Phytochemistry 1988, 27, 969–978. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Spiegelman, B.M.; Flier, J.S. Adipogenesis and obesity: Rounding out the big picture. Cell 1999, 87, 377–389. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 2004, 114, 1752–1761. [Google Scholar]
- Hsu, C.L.; Yen, G.C. Phenolic compounds: Evidence for inhibitory effects against obesity and their underlying molecular signaling mechanisms. Mol. Nutr. Food Res. 2008, 52, 53–61. [Google Scholar] [CrossRef]
- Lamien-Meda, A.; Lamien, C.E.; Compaoré, M.M.; Meda, R.N.; Kiendrebeogo, M.; Zeba, B.; Millogo, J.F.; Nacoulma, O.G. Polyphenol content and antioxidant activity of fourteen wild edible fruits from Burkina Faso. Molecules 2008, 13, 581–594. [Google Scholar] [CrossRef]
- Hanasaki, Y.; Ogawa, S.; Fukui, S. The correlation between active oxygens scavenging and antioxidative effects of flavonoids. Free Radic. Biol. Med. 1994, 16, 845–850. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free. Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Koleva, I.I.; van Beek, T.A.; Linssen, J.P.; de Groot, A.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Mosquera, O.M.; Correa, Y.M.; Buitrago, D.C.; Niño, J. Antioxidant activity of twenty five plants from Colombian biodiversity. Mem. Inst. Oswaldo Cruz. 2007, 102, 631–634. [Google Scholar] [CrossRef]
- MacDonald-Wicks, L.K.; Wood, L.G.; Garg, M.L. Methodology for the determination of biological antioxidant capacity in vitro: A review. J. Sci. Food Agric. 2006, 86, 2046–2056. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Duh, P.D. Antioxidant activity of burdock (Arctium lappa Linné): Its scavenging effect on free-radical and active oxygen. J. Am. Oil Chem. Soc. 1998, 75, 455–461. [Google Scholar] [CrossRef]
- Oyaizu, M. Antioxidative activities of browning products of glucosamine fractionated by organic solvent and thin-layer chromatography. Nippon Shokuhin Kogyo Gakkaishi 1988, 35, 771–775. [Google Scholar] [CrossRef]
- Tachakittirungrod, S.; Okonogi, S.; Chowwanapoonpohn, S. Study on antioxidant activity of certain plants in Thailand: Mechanism of antioxidant action of guava leaf extract. Food Chem. 2007, 103, 381–388. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Calzadilla, P.; Sapochnik, D.; Cosentino, S.; Diz, V.; Dicelio, L.; Calvo, J.C.; Guerra, L.N. N-acetylcysteine reduces markers of differentiation in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2011, 12, 6936–6951. [Google Scholar] [CrossRef]
- Baehner, R.L.; Boxer, L.A.; Davis, J. The biochemical basis of nitroblue tetrazolium reduction in normal human and chronic granulomatous disease polymorphonuclear leukocytes. Blood 1976, 48, 309–313. [Google Scholar]
- Alañóna, M.E.; Castro-Vázqueza, L.; Díaz-Marotob, M.C.; Gordonc, M.H.; Pérez-Coelloa, M.S. A study of the antioxidant capacity of oak wood used in wine ageing and the correlation with polyphenol composition. Food Chem. 2011, 128, 997–1002. [Google Scholar] [CrossRef]
- Chun, O.K.; Kim, D.O.; Moon, H.Y.; Kang, H.G.; Lee, C.Y. Contribution of individual polyphenolics to total antioxidant capacity of plums. J. Agric. Food Chem. 2003, 51, 7240–7245. [Google Scholar] [CrossRef]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant activity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Roy, M.K.; Koide, M.; Rao, T.P.; Okubo, T.; Ogasawara, Y.; Juneja, L.R. ORAC and DPPH assay comparison to assess antioxidant capacity of tea infusions: Relationship between total polyphenol and individual catechin content. Int. J. Food Sci. Nutr. 2010, 61, 109–124. [Google Scholar] [CrossRef]
- Gutfinger, T. Polyphenols in olive oils. J. Am. Oil Chem. Soc. 1981, 58, 966–968. [Google Scholar] [CrossRef]
- Moreno, M.I.; Isla, M.I.; Sampietro, A.R.; Vattuone, M.A. Comparison of the free radical-scavenging activity of propolis from several regions of Argentina. J. Ethnopharmacol. 2000, 71, 109–114. [Google Scholar] [CrossRef]
- Chu, Y.H.; Chang, C.L.; Hsu, H.F. Flavonoid content of several vegetables and their antioxidant activity. J. Sci. Food Agric. 2000, 80, 561–566. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Lee, O.H.; Kwon, Y.I.; Apostolidis, E.; Shetty, K.; Kim, Y.C. Rhodiola-induced inhibition of adipogenesis involves antioxidant enzyme response associated with pentose phosphate pathway. Phytother. Res. 2011, 25, 106–115. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, Y.-J.; Kim, D.-B.; Lee, J.S.; Cho, J.-H.; Kim, B.K.; Choi, H.-S.; Lee, B.-Y.; Lee, O.-H. Antioxidant Activity and Anti-Adipogenic Effects of Wild Herbs Mainly Cultivated in Korea. Molecules 2013, 18, 12937-12950. https://doi.org/10.3390/molecules181012937
Lee Y-J, Kim D-B, Lee JS, Cho J-H, Kim BK, Choi H-S, Lee B-Y, Lee O-H. Antioxidant Activity and Anti-Adipogenic Effects of Wild Herbs Mainly Cultivated in Korea. Molecules. 2013; 18(10):12937-12950. https://doi.org/10.3390/molecules181012937
Chicago/Turabian StyleLee, Young-Jun, Dan-Bi Kim, Jong Seok Lee, Ju-Hyun Cho, Bong Kyun Kim, Hyeon-Son Choi, Boo-Yong Lee, and Ok-Hwan Lee. 2013. "Antioxidant Activity and Anti-Adipogenic Effects of Wild Herbs Mainly Cultivated in Korea" Molecules 18, no. 10: 12937-12950. https://doi.org/10.3390/molecules181012937