NMR Study on Small Proteins from Helicobacter pylori for Antibiotic Target Discovery: A Review

Abstract
:1. Development of Antibiotics
2. Helicobacter pylori as Antibiotics Target Discovery

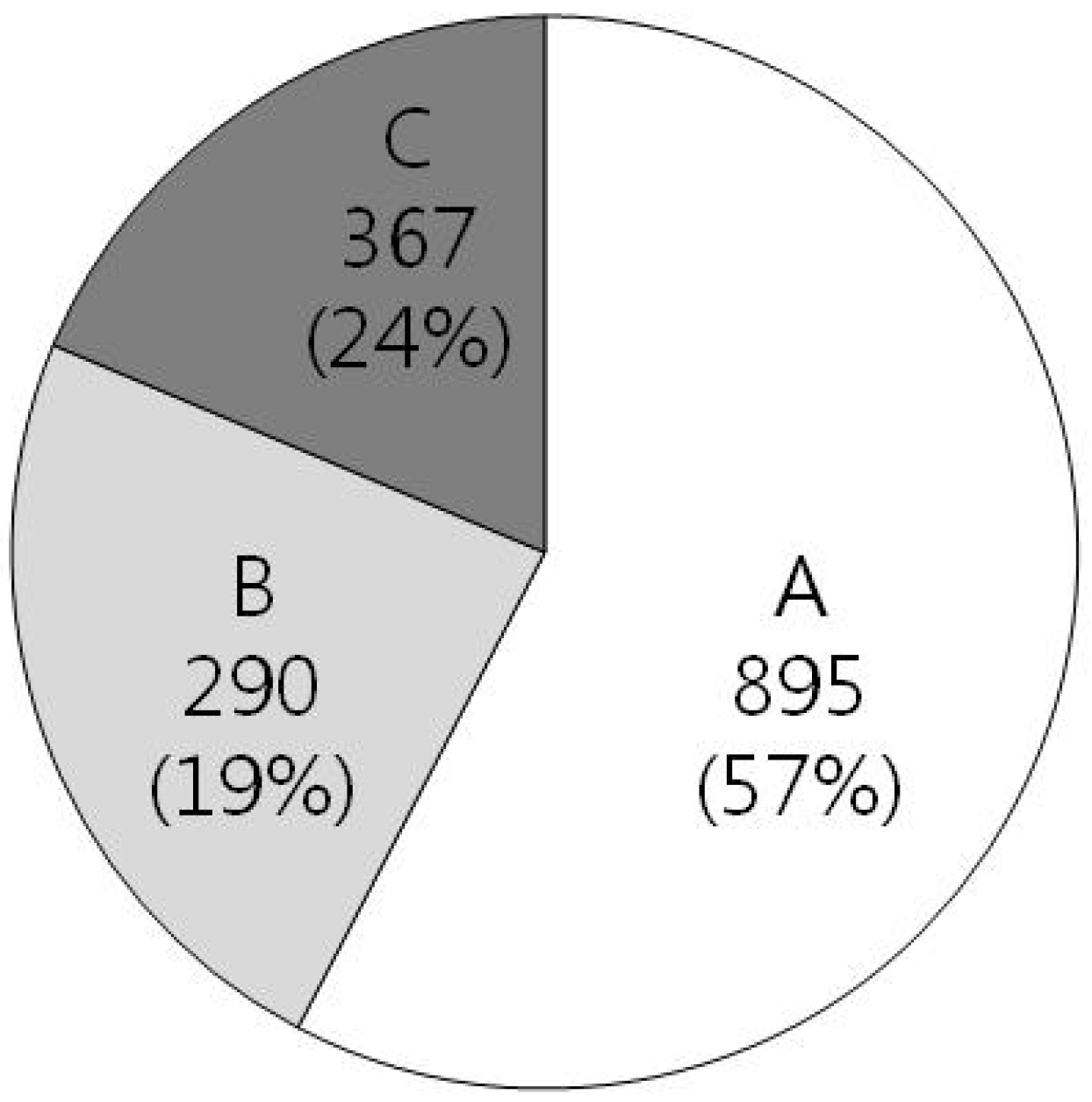
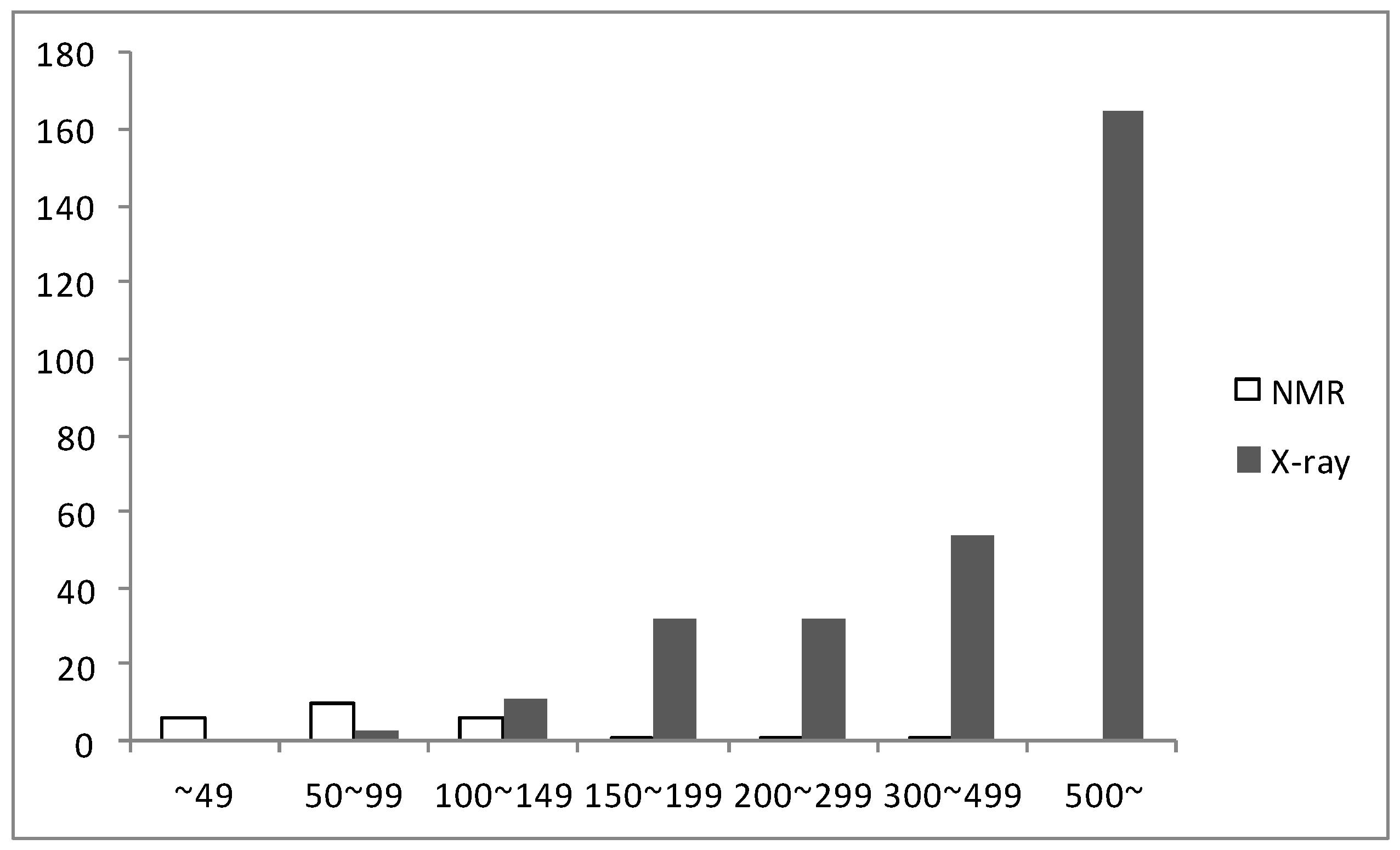
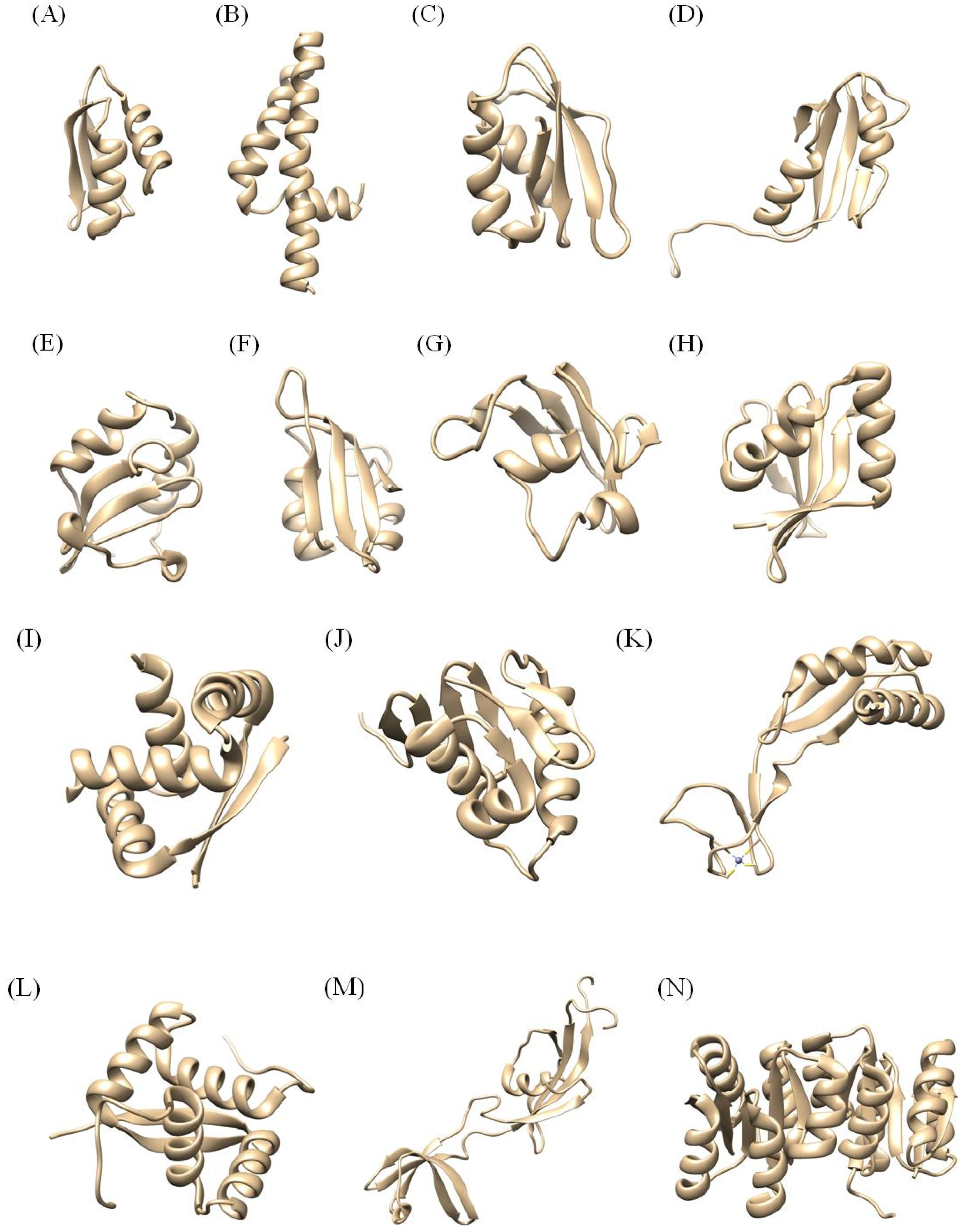
3. Structures of 25 NMR Proteins from H. pylori



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB ID | Macromolecule Name | No. of AA | Function | Year | |
|---|---|---|---|---|---|
| 1 | 1P5K | - | 19 | ribosome | 2003 |
| 2 | 1P5L | - | 19 | ribosome | 2003 |
| 3 | 1P0G | - | 19 | ribosome | 2003 |
| 4 | 1P0J | - | 19 | ribosome | 2003 |
| 5 | 1P0L | - | 19 | ribosome | 2003 |
| 6 | 1P0O | - | 19 | ribosome | 2003 |
| 7 | 1OT0 | - | 19 | antibiotic | 2004 |
| 8 | 1YG0 | HP1073 | 66 | metal transport | 2006 |
| 9 | 1ZHC | HP1242 | 76 | hypothetical protein | 2005 |
| 10 | 2LXR | HP1264 | 76 | oxidoreductase | 2012 |
| 11 | 2H9Z | HP0495 | 86 | hypothetical protein | 2007 |
| 12 | 1Z8M | HP0894 | 88 | hypothetical protein | 2005 |
| 13 | 2KI2 | HP0827 | 90 | RNA binding protein/ ssDNA binding protein | 2009 |
| 14 | 2JOQ | - | 91 | hypothetical protein | 2007 |
| 15 | 2K6P | HP1423 | 92 | unknown function | 2009 |
| 16 | 2OTR | HP0892 | 98 | hypothetical protein | 2007 |
| 17 | 2HQN | - | 109 | singnaling protein | 2007 |
| 18 | 1X93 | HP0222 | 110 | transcription | 2005 |
| 19 | 2K0Z | HP1203 | 110 | unknown function | 2008 |
| 20 | 2K4J | - | 115 | transcription | 2008 |
| 21 | 2KDX | HP0869 | 119 | metal binding protein | 2009 |
| 22 | 2K1O | HP0564 | 132 | gene regulation | 2008 |
| 23 | 2KR7 | HP1123 | 151 | isomerase | 2010 |
| 24 | 2HQO | - | 246 | singnaling protein | 2007 |
| 25 | 2HQR | HP1043 | 446 | singnaling protein | 2007 |
3.1. Proteins under 20 Amino Acid Residues: 1P0O, 1P0L, 1P0J, 1P0G, 1P5L, 1P5K, 1OT0
3.2. Proteins between 20 and 99 Amino Acid Residues: 1YG0, 1ZHC, 2LXR, 2H9Z, 1Z8M, 2KI2, 2JOQ, 2K6P, 2OTR
3.3. Proteins between 100 and 200 Amino Acid residues: 2HQN, 1X93, 2K0Z, 2K4J, 2KDX, 2K1O, 2KR7
3.4. Proteins over 200 Amino Acid Residues: 2HQO, 2HQR
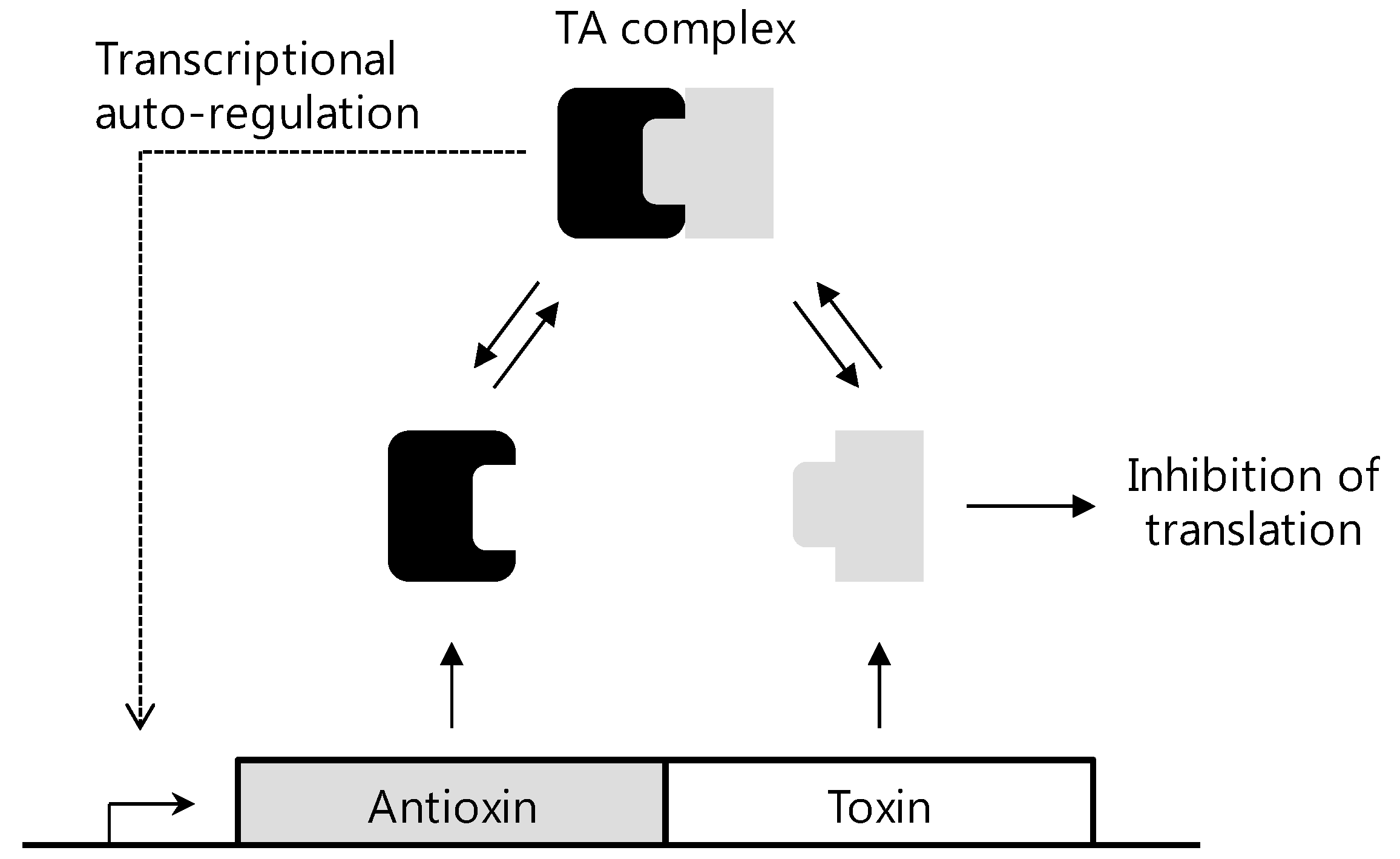
4. Structure and Function Studies of HP0894 and HP0892

5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Waksman, S.A. What is an antibiotic or an antibiotic substance? Mycologia 1947, 39, 565–569. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: from targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef]
- Walsh, C. Where will new antibiotics come from? Nat. Rev. Microbiol. 2003, 1, 65–70. [Google Scholar] [CrossRef]
- Fernandes, P. Antibacterial discovery and development-the failure of success? Nat. Biotechnol. 2006, 24, 1497–1503. [Google Scholar] [CrossRef]
- Fernandes, P.B. The ATCG of drug discovery. Curr. Opin. Mol. Ther. 2000, 2, 624–632. [Google Scholar]
- Schmid, M.B. Crystallizing new approaches for antimicrobial drug discovery. Biochem. Pharmacol. 2006, 71, 1048–1056. [Google Scholar] [CrossRef]
- Waldor, M.K. Disarming pathogens-a new approach for antibiotic development. N. Engl. J. Med. 2006, 354, 296–297. [Google Scholar] [CrossRef]
- Casey, G. Antibiotics and the rise of superbugs. Nurs. N. Z. 2012, 18, 20–24. [Google Scholar]
- Quinlivan, E.P.; Gregory, J.F. Homocysteine, B vitamins, and cardiovascular disease. N. Engl. J. Med. 2006, 355, 209–211. [Google Scholar]
- Tomb, J.F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A.; et al. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef]
- Hessey, S.J.; Spencer, J.; Wyatt, J.I.; Sobala, G.; Rathbone, B.J.; Axon, A.T.; Dixon, M.F. Bacterial adhesion and disease activity in Helicobacter associated chronic gastritis. Gut 1990, 31, 134–138. [Google Scholar] [CrossRef]
- Kazi, J.L.; Sinniah, R.; Zaman, V.; Ng, M.L.; Jafarey, N.A.; Alam, S.M.; Zuberi, S.J.; Kazi, A.M. Ultrastructural study of Helicobacter pylori-associated gastritis. J. Pathol. 1990, 161, 65–70. [Google Scholar] [CrossRef]
- Krienitz, U. Über das Auftreten von Spirochäten verschiedener Form im Mageninhalt bei Carcinoma ventriculi. Dtsch. Med. Wochenschr. 1906, 32, 872. [Google Scholar] [CrossRef]
- Warren, J.R.; Marshall, B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1983, 1, 1273–1275. [Google Scholar]
- Infection with Helicobacter pylori. In Schistosomes, Liver Flukes and Helicobacter pylori. International Agency for Research on Cancer; WHO: Lyon, France, 1994; Volume 61, pp. 177–240.
- Eaton, K.A.; Gilbert, J.V.; Joyce, E.A.; Wanken, A.E.; Thevenot, T.; Baker, P.; Plaut, A.; Wright, A. In vivo complementation of ureB restores the ability of Helicobacter pylori to colonize. Infect. Immun. 2002, 70, 771–778. [Google Scholar] [CrossRef]
- Rothenbacher, D.; Brenner, H. Burden of Helicobacter pylori and H. pylori-related diseases in developed countries: Recent developments and future implications. Microbes Infect. 5, 693–703.
- Wotherspoon, A.C.; Doglioni, C.; Diss, T.C.; Pan, L.; Moschini, A.; de Boni, M.; Isaacson, P.G. Regression of primary low-grade B-cell gastric lymphoma of mucosa-associated lymphoid tissue type after eradication of Helicobacter pylori. Lancet 1993, 342, 575–577. [Google Scholar] [CrossRef]
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef]
- Suerbaum, S.; Achtman, M. Helicobacter pylori: recombination, population structure and human migrations. Int J. Med. Microbiol. 2004, 294, 133–139. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O'Morain, C.; Bazzoli, F.; El-Omar, E.; Graham, D.; Hunt, R.; Rokkas, T.; Vakil, N.; Kuipers, E.J. Current concepts in the management of Helicobacter pylori infection: The Maastricht III Consensus Report. Gut 2007, 56, 772–781. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Calvet, X.; Gomollon, F.; Mones, J. Eradication treatment of Helicobacter pylori. Recommendations of the II Spanish Consensus Conference. Med. Clin. 2005, 125, 301–316. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Pajares, J.M. Treatment of Helicobacter pylori infection: the past and the future. Eur. J. Intern. Med. 2010, 21, 357–359. [Google Scholar] [CrossRef]
- Tominaga, K.; Higuchi, K.; Hamasaki, N.; Hamaguchi, M.; Takashima, T.; Tanigawa, T.; Watanabe, T.; Fujiwara, Y.; Tezuka, Y.; Nagaoka, T.; et al. In vivo action of novel alkyl methyl quinolone alkaloids against Helicobacter pylori. J. Antimicrob. Chemother. 2002, 50, 547–552. [Google Scholar] [CrossRef]
- Koga, T.; Inoue, H.; Ishii, C.; Okazaki, Y.; Domon, H.; Utsui, Y. Effect of plaunotol in combination with clarithromycin or amoxicillin on Helicobacter pylori in vitro in vitro and in vivo. J. Antimicrob. Chemother. 2002, 50, 133–136. [Google Scholar] [CrossRef]
- Alm, R.A.; Ling, L.S.; Moir, D.T.; King, B.L.; Brown, E.D.; Doig, P.C.; Smith, D.R.; Noonan, B.; Guild, B.C.; de Jonge, B.L.; et al. Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature 1999, 397, 176–180. [Google Scholar] [CrossRef]
- Salgado, F.; Garcia, A.; Onate, A.; Gonzalez, C.; Kawaguchi, F. Increased in vitro and in vivo biological activity of lipopolysaccharide extracted from clinical low virulence vacA genotype Helicobacter pylori strains. J. Med. Microbiol. 2002, 51, 771–776. [Google Scholar]
- Selbach, M.; Moese, S.; Hauck, C.R.; Meyer, T.F.; Backert, S. Src is the kinase of the Helicobacter pylori CagA protein in vitro in vitro and in vivo. J. Biol. Chem. 2002, 277, 6775–6778. [Google Scholar]
- Park, S.J.; Son, W.S.; Lee, B.J. Structural Analysis of Hypothetical Proteins from Helicobacter pylori: An Approach to Estimate Functions of Unknown or Hypothetical Proteins. Int J. Mol. Sci 2012, 13, 7109–7137. [Google Scholar] [CrossRef]
- Yeo, H.J.; Savvides, S.N.; Herr, A.B.; Lanka, E.; Waksman, G. Crystal structure of the hexameric traffic ATPase of the Helicobacter pylori type IV secretion system. Mol. Cell. 2000, 6, 1461–1472. [Google Scholar] [CrossRef]
- Lee, K.H.; Lee, D.G.; Park, Y.; Kang, D.I.; Shin, S.Y.; Hahm, K.S.; Kim, Y. Interactions between the plasma membrane and the antimicrobial peptide HP (2–20) and its analogues derived from Helicobacter pylori. Biochem. J. 2006, 394, 105–114. [Google Scholar] [CrossRef]
- Park, S.J.; Jung, Y.S.; Kim, J.S.; Seo, M.D.; Lee, B.J. Structural insight into the distinct properties of copper transport by the Helicobacter pylori CopP protein. Proteins 2008, 71, 1007–1019. [Google Scholar] [CrossRef]
- Kang, S.J.; Park, S.J.; Jung, S.J.; Lee, B.J. Solution structure of HP1242 from Helicobacter pylori. Proteins 2005, 61, 1111–1113. [Google Scholar] [CrossRef]
- Kang, S.J.; Park, S.J.; Jung, S.J.; Lee, B.J. Backbone 1H, 15N, and 13C resonance assignment of HP1242 from Helicobacter pylori. J. Biochem. Mol. Biol. 2005, 38, 591–594. [Google Scholar] [CrossRef]
- Lee, K.Y.; Kim, J.H.; Lee, J.; Lee, I.; Bae, Y.J.; Lee, B.J. Structural Characterization of HP1264 Reveals a Novel Fold for the Flavin Mononucleotide Binding Protein. Biochemistry 2013, 52, 1583–1593. [Google Scholar]
- Seo, M.D.; Park, S.J.; Kim, H.J.; Lee, B.J. Solution structure of hypothetical protein, HP0495 (Y495_HELPY) from Helicobacter pylori. Proteins 2007, 67, 1189–1192. [Google Scholar] [CrossRef]
- Seo, M.D.; Park, S.J.; Kim, H.J.; Seok, S.H.; Lee, B.J. Backbone 1H, 15N, and 13C resonance assignment and secondary structure prediction of HP0495 from Helicobacter pylori. J. Biochem. Mol. Biol. 2007, 40, 839–843. [Google Scholar]
- Han, K.D.; Park, S.J.; Jang, S.B.; Son, W.S.; Lee, B.J. Solution structure of conserved hypothetical protein HP0894 from Helicobacter pylori. Proteins 2005, 61, 1114–1116. [Google Scholar] [CrossRef]
- Han, K.D.; Park, S.J.; Lee, B.J. Backbone 1H, 15N, and 13C resonance assignments and secondary-structure of conserved hypothetical protein HP0894 from Helicobacter pylori. Mol. Cells 2005, 20, 442–445. [Google Scholar]
- Jang, S.B.; Ma, C.; Lee, J.Y.; Kim, J.H.; Park, S.J.; Kwon, A.R.; Lee, B.J. NMR solution structure of HP0827 (O25501_HELPY) from Helicobacter pylori: model of the possible RNA-binding site. J. Biochem. 2009, 146, 667–674. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.J.; Lee, K.Y.; Son, W.S.; Sohn, N.Y.; Kwon, A.R.; Lee, B.J. Solution structure of hypothetical protein HP1423 (Y1423_HELPY) reveals the presence of alphaL motif related to RNA binding. Proteins 2009, 75, 252–257. [Google Scholar] [CrossRef]
- Han, K.D.; Park, S.J.; Jang, S.B.; Lee, B.J. Backbone 1H, 15N, and 13C resonance assignments and secondary-structure of the conserved hypothetical protein HP0892 of Helicobacter pylori. Mol. Cells 2008, 25, 138–141. [Google Scholar]
- Han, K.D.; Park, S.J.; Jang, S.B.; Lee, B.J. Solution structure of conserved hypothetical protein HP0892 from Helicobacter pylori. Proteins 2008, 70, 599–602. [Google Scholar]
- Hong, E.; Lee, H.M.; Ko, H.; Kim, D.U.; Jeon, B.Y.; Jung, J.; Shin, J.; Lee, S.A.; Kim, Y.; Jeon, Y.H.; et al. Structure of an atypical orphan response regulator protein supports a new phosphorylation-independent regulatory mechanism. J. Biol. Chem. 2007, 282, 20667–20675. [Google Scholar] [CrossRef]
- Popescu, A.; Karpay, A.; Israel, D.A.; Peek, R.M., Jr.; Krezel, A.M. Helicobacter pylori protein HP0222 belongs to Arc/MetJ family of transcriptional regulators. Proteins 2005, 59, 303–311. [Google Scholar] [CrossRef]
- Gupta, S.S.; Borin, B.N.; Cover, T.L.; Krezel, A.M. Structural analysis of the DNA-binding domain of the Helicobacter pylori response regulator ArsR. J. Biol. Chem. 2009, 284, 6536–6545. [Google Scholar]
- Xia, W.; Li, H.; Sze, K.H.; Sun, H. Structure of a nickel chaperone, HypA, from Helicobacter pylori reveals two distinct metal binding sites. J. Am. Chem. Soc. 2009, 131, 10031–10040. [Google Scholar] [CrossRef]
- Borin, B.N.; Krezel, A.M. Structure of HP0564 from Helicobacter pylori identifies it as a new transcriptional regulator. Proteins 2008, 73, 265–268. [Google Scholar] [CrossRef]
- Han, K.D.; Matsuura, A.; Ahn, H.C.; Kwon, A.R.; Min, Y.H.; Park, H.J.; Won, H.S.; Park, S.J.; Kim, D.Y.; Lee, B.J. Functional identification of toxin-antitoxin molecules from Helicobacter pylori 26695 and structural elucidation of the molecular interactions. J. Biol. Chem. 2011, 286, 4842–4853. [Google Scholar] [CrossRef]
- Han, K.D.; Ahn, D.H.; Lee, S.A.; Min, Y.H.; Kwon, A.R.; Ahn, H.C.; Lee, B.J. Identification of chromosomal HP0892-HP0893 toxin-antitoxin proteins in Helicobacter pylori and structural elucidation of their protein-protein interaction. J. Biol. Chem. 2013, 288, 6004–6013. [Google Scholar]
- Engelberg-Kulka, H.; Amitai, S.; Kolodkin-Gal, I.; Hazan, R. Bacterial programmed cell death and multicellular behavior in bacteria. PLoS Genet. 2006, 2, e135. [Google Scholar] [CrossRef]
- Van Melderen, L.; Saavedra De Bast, M. Bacterial toxin-antitoxin systems: More than selfish entities? PLoS Genet. 2009, 45, e1000437. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Park, J.H.; Inouye, M. Toxin-antitoxin systems in bacteria and archaea. Annu. Rev. Genet. 2011, 45, 61–79. [Google Scholar] [CrossRef]
- Park, S.J.; Son, W.S.; Lee, B.J. Structural overview of toxin-antitoxin systems in infectious bacteria: A target for developing anti microbial agents. Biochim. Biophys. Acta 2013, 1834, 1155–1167. [Google Scholar]
- Gerdes, K.; Christensen, S.K.; Lobner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kang, S.-J.; Kim, D.-H.; Lee, B.-J. NMR Study on Small Proteins from Helicobacter pylori for Antibiotic Target Discovery: A Review. Molecules 2013, 18, 13410-13424. https://doi.org/10.3390/molecules181113410
Kang S-J, Kim D-H, Lee B-J. NMR Study on Small Proteins from Helicobacter pylori for Antibiotic Target Discovery: A Review. Molecules. 2013; 18(11):13410-13424. https://doi.org/10.3390/molecules181113410
Chicago/Turabian StyleKang, Su-Jin, Do-Hee Kim, and Bong-Jin Lee. 2013. "NMR Study on Small Proteins from Helicobacter pylori for Antibiotic Target Discovery: A Review" Molecules 18, no. 11: 13410-13424. https://doi.org/10.3390/molecules181113410