Investigating the Role of Polyunsaturated Fatty Acids in Bone Development Using Animal Models


Abstract
:1. Introduction
1.1. Bone Development and Peak Bone Mass

{kind=link}
{kind=link}
| Study subject and age | PUFA source | Amount of PUFA | Control group | Treatment duration | Skeletal site | Outcome on fatty acid composition | Reference |
|---|---|---|---|---|---|---|---|
| Male quails (1–8 months) | Menhaden oil | n-6/n-3 ratio: 0.66 | Soy bean oil n-6/n-3 ratio: 12.6 | 7 months | Tibia cortical bone |
| [12] |
| Female rats (0–12 weeks) | ALA (flaxseed oil) group (n-3 adq), ALA+DHA (DHASCO oil) group | Total n-3 PUFA: 3.12% in n-3 adq group, 3.83% in n-3 supp group | LA group (n-3 def), high LA group (n-3 def) (LA from safflower oil) | Dams exposed to diet 5 wks before conception. Pups exposed until 12 wks old | Femur cortical bone (results similar in tibia but not shown) |
| [13] |
| Male rats (3–9 weeks) | Mixture of safflower oil and menhaden oil | n-6/n-3 ratio: 1.2, 2.6, 9.8, 23.8 | N/A | 6 weeks | Femur periosteum polar lipids |
| [14] |
| Femur cortical polar lipids |
| [14] | |||||
| Male and female rats (7–15 weeks) | AIN-93G for n-3 adequate group, Flaxseed oil and DHA for repletion group (n-6/n-3 ratio | n-3 adequate group, n-3 repleted group | Safflower and coconut oil for n-3 depleted group n-6/n-3 ratio: 37 | 8 weeks | Femur, tibia cortical bone |
| [15] |
| Male and female Fat-1 mice (0–12 weeks) | Modified AIN-93G, Fat-1 mice produce n-3 from n-6 in safflower oil | 10% w/w safflower oil | Age matched wild-type mice as controls | 12 weeks | Femur |
| [16] |
| Lumbar Vertebrae 5-6 |
| [16] | |||||
| Male rats (7–16 weeks) | Flaxseed oil for n-3 group, safflower oil for n-6 group | 20% w/w flaxseed oil, n-6/n-3: 0.21 | Chow diet: n-6/n-3: 9.46 | 9 weeks | Femur bone marrow, diaphysis, epiphysis (proximal and distal) |
| [17] |
| Male rats (0–15 weeks) | AIN-93G with flax for n-3 adequate group, DHASCO for DHA group | ALA 3.1% total lipids, n-6/n-3:5, DHA group 1% total lipids DHA, n-6/n-3: 14.2 | Safflower oil (n-6/n-3: 383.7) | 15 weeks | Femur and tibia bone marrow and periosteum |
| [18] |
| Male and female rabbits | Soybean oil (LA/ALA), sesame oil (LA), fish oil (EPA &DHA), algae oil (DHA&AA) | 7% w/w oils in diet, soybean oil n-6/n-3: 8.68, fish oil: n-6/n-3: 0.39, algae oil: n-6/n-3: 0.63 | Sesame oil n-6/n-3: 21.75 | 14 weeks | Tibia, femur and humerus bone marrow |
| [19] |
| Study subject and age | PUFA source | Amount of PUFA | Control group | Treatment duration | Bone mineral | Biomechanical strength | Reference |
|---|---|---|---|---|---|---|---|
| Female rats (terminated at 30 weeks of age) | Flaxseed oilSoybean oil | n-6/n-3 ratio: 0.4 for n-3 group, 9 for n-6 + n-3 group | Sunflower seed oil (n-6 group) n-6/n-3 ratio: 216 | 31 days (last 10 days of gestation and first 3 weeks of lactation) |
|
| [20] |
| Male and female mice (4–13 weeks) | Flaxseed oil | 10% flaxseed oil n-6/n-3 ratio: 0.25 | 10% corn oil n-6/n-3 ratio: 57 | 9 weeks |
|
| [21] |
| Male piglets (5–26 days) | Soybean, safflower, coconut, and flaxseed oils | n-6/n-3 ratio: 4.5 | n-6/n-3 ratio: 9.0 | 21 days |
|
| [22] |
| Female rats (4–12 weeks) | Flaxseed oil | Flaxseed oil (12% w/w) n-6/n-3 ratio: 0.33 | Corn oil n-6/n-3: 73 | 8 weeks |
|
| [23] |
| Female chickens (16 weeks to 58 weeks) | Flaxseed oil | Varying amounts of flaxseed oil and corn oil in diet n-6/n-3: 47.8–4.8 | N/A | 42 weeks |
|
| [24] |
| Male rats (7–16 weeks) | Flaxseed oil | 20% w/w flaxseed oil n-6/n-3 ratio: 0.21 | Chow diet, n-6/n-3: 9.46 | 9 weeks |
|
| [17] |
| Male rats (4–12 weeks) | Flaxseed oil | 20% w/w n-6/n-3: 0.4 | 20% w/w Corn oil n-6/n-3 ratio: 9.0 | 8 weeks |
|
| [25] |
| Male rats (conception-7 weeks or conception-19 weeks) | Flaxseed oil | 10% ground flaxseed, approx. 4% w/w flaxseed oil | AIN-93G with safflower oil, n-6/n-3: 6 | Gestation and lactation, or continuation until 7 weeks or 19 weeks |
|
| [26] |
| Study subject and age | PUFA source | Amount of PUFA | Control group | Treatment duration | Bone mineral | Biomechanical strength | Reference |
|---|---|---|---|---|---|---|---|
| Male quails (1–8 months) | Menhaden oil | n-6/n-3 ratio: 0.66 | Soy bean oil n-6/n-3 ratio: 12.6 | 7 months |
|
| [12] |
| Male rats (4–9 weeks) | Menhaden oil | 4% menhaden, 3% corn—7% w/w diet n-6/n-3 ratio: 1.4 | AIN-93G with 7% soybean oil n-6/n-3 ratio: 7.1 | 5 weeks |
|
| [27] |
| Male mice (6–12 weeks) | Tuna oil | 4% tuna oil + 1% corn oil n-6/n-3 ratio: 0.5 26.3% DHA | 5% corn oil n-6/n-3 ratio: 45.3 | 6 weeks |
|
| [28] |
| Fish oil | 4% fish oil + 1% corn oil n-6/n-3 ratio: 0.5 11.7% DHA |
|
| [28] | |||
| Male and female rats (3–8 weeks) | Menhaden oil | 6% menhaden oil + 1% soybean oil | 7% soybean oil | 5 weeks |
|
| [29] |
| Male and female rats (7–15 weeks) | AIN-93G for n-3 adequate group, DHASCO for repletion group | Repletion diet: AIN-93G with DHASCO (3.0 g/kg diet) ALA: 2.6% of total fatty acids DHA: 1.3% of total fatty acids n-6/n-3 ratio: 4 | Safflower and coconut oil for n-3 depleted group n-6/n-3 ratio: 378 | 8 weeks |
|
| [15] |
| Male rats (0–15 weeks) | DHA (DHASCO, ALA from Flaxseed oil | Flaxseed ALA 3.1% total lipids, n-6/n-3:5, DHA group 1% total lipids DHA, n-6/n-3: 14.2 | Safflower oil (n-6/n-3: 383.7) | 15 weeks |
|
| [18] |
| Female rats (4–12 weeks) | Krill oil | 12% w/w diet, n-6/n-3: 0.03 | Corn oil, 12% w/w diet | 8 weeks |
|
| [23] |
| Salmon Oil | 12%w/w diet, n-6/n-3: 0.04 |
|
| [23] | |||
| Tuna Oil | 12%w/w diet, n-6/n-3: 0.04 |
|
| [23] | |||
| Menhaden oil | 12%w/w diet, n-6/n-3: 0.04 |
|
| [23] | |||
| Female rats (0–21 weeks) | Menhaden oil | 6.5% w/w menhaden oil diet, 1% safflower oil, n-6/n-3: 0.5 | Safflower oil 7.5% w/w diet, n-6/n-3: 250 | 21 weeks |
|
| [30] |
| Male rats (8–10 weeks) | Fish oil | 15% w/w diet as fish oil, n-6/n-3: 0.52 | 15% w/w diet soybean oil, n-6/n-3: 11.7 | 15 days |
|
| [31] |
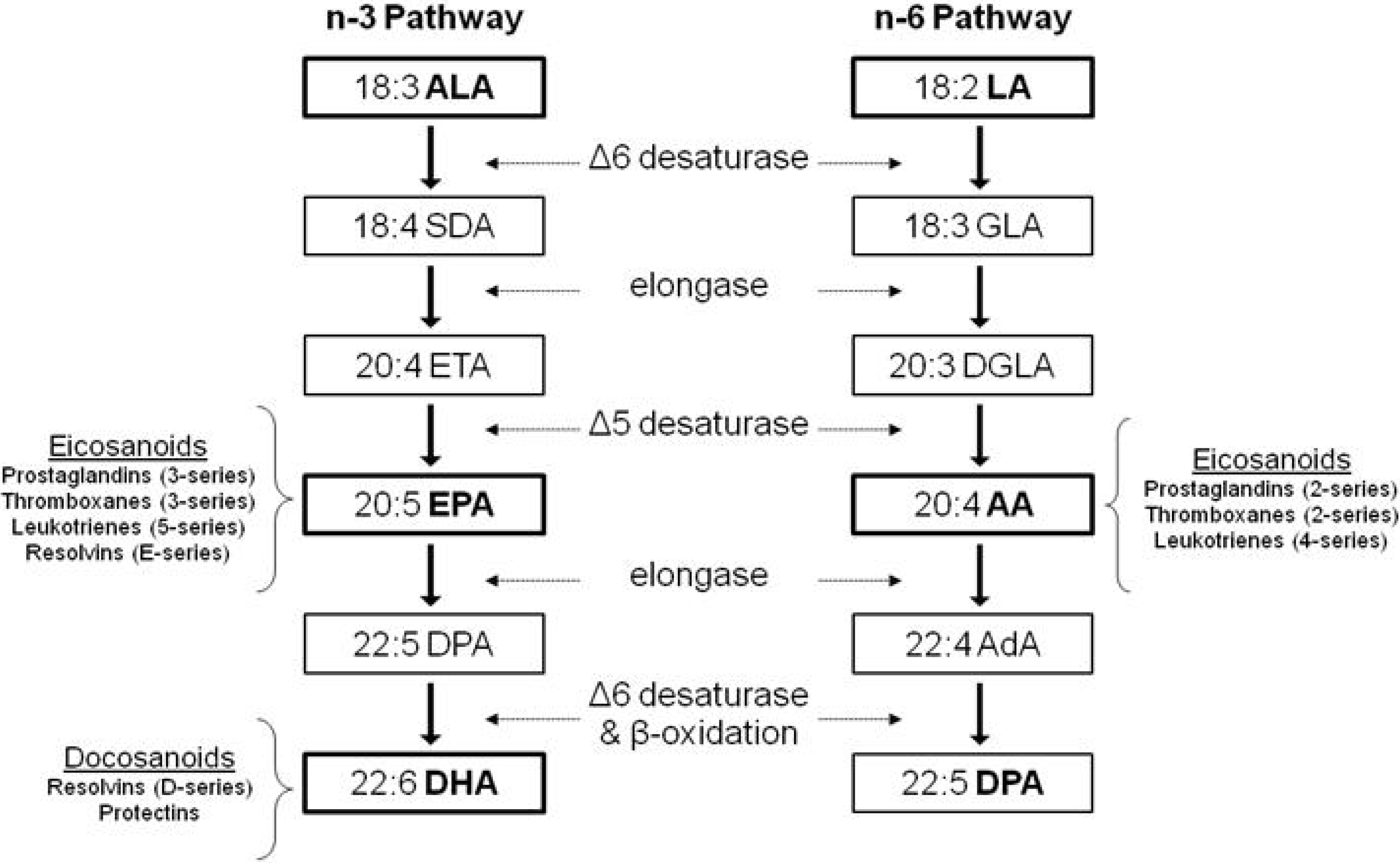
1.2. Dietary Polyunsaturated Fatty Acids (PUFA)

2. n-3 PUFA and Its Effect on Bone Fatty Acid Composition
2.1. ALA Studies
2.2. EPA and DHA Studies
2.3. Effect of ALA vs. EPA and DHA
3. PUFA and Bone Mineral Content and Strength
3.1. ALA Studies
3.2. EPA and DHA Studies
3.3. Comparison of ALA vs. EPA/DHA Studies
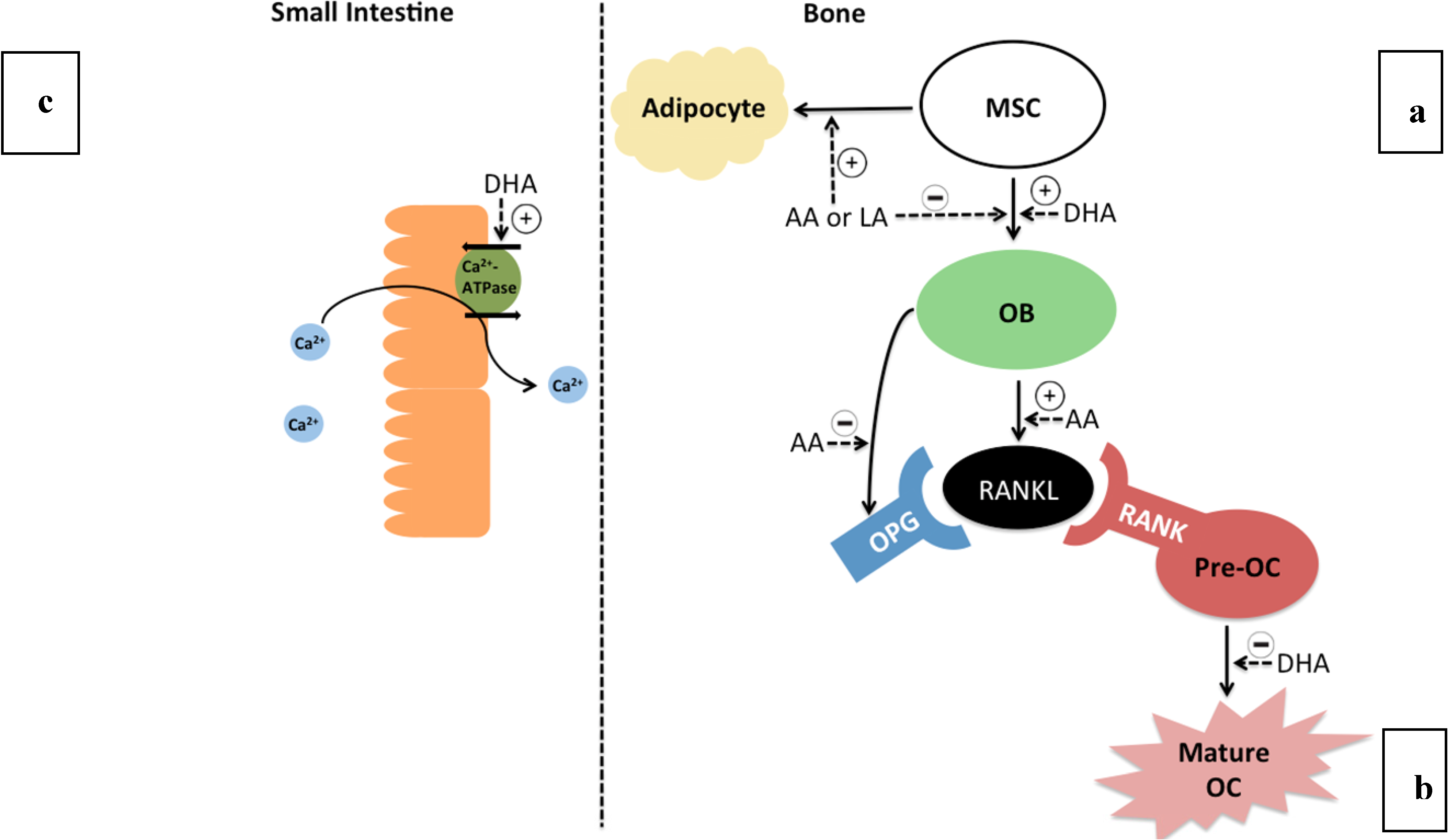
4. Mechanisms of PUFA in Bone Metabolism

5. Knowledge Gaps and Future Directions
| Research Area | Current State | Knowledge Gap | Future Directions |
|---|---|---|---|
| Dose and Type of PUFA |
|
|
|
| Timing of Exposure |
|
|
|
| Gender Differences |
|
|
|
5.1. Dose and Type of PUFA
5.2. Timing of Exposure
5.3. Gender Differences
6. Summary and Conclusions
- There is a probable role for n-3 PUFA in the development of stronger bones, with EPA and DHA intervention being more efficacious than ALA. Whether this translates to reduced fracture risk in humans remains to be determined.
- Both n-3 and n-6 PUFA play a role in bone development. Future research should be aimed at determining the dose, duration, and timing of exposure to individual n-3 and n-6 PUFA throughout the lifecycle.
- n-3 PUFA may improve bone health by increasing calcium absorption in the gut, and increasing osteoblast differentiation and activity, reducing osteoclast activity and promoting deposition of mineral in developing bones. These mechanisms require further study.
Acknowledgments
Conflicts of Interest
References
- Chen, B.; McClements, D.J.; Decker, E.A. Design of foods with bioactive lipids for improved health. Annu. Rev. Food Sci. Technol. 2013, 4, 35–56. [Google Scholar] [CrossRef]
- Lim, S.Y.; Hoshiba, J.; Moriguchi, T.; Salem, N., Jr. N-3 fatty acid deficiency induced by a modified artificial rearing method leads to poorer performance in spatial learning tasks. Pediatr. Res. 2005, 58, 741–748. [Google Scholar] [CrossRef]
- Llanos, A.; Lin, Y.; Mena, P.; Salem, N., Jr.; Uauy, R. Infants with intrauterine growth restriction have impaired formation of docosahexaenoic acid in early neonatal life: A stable isotope study. Pediatr. Res. 2005, 58, 735–740. [Google Scholar] [CrossRef]
- Whalley, L.J.; Fox, H.C.; Wahle, K.W.; Starr, J.M.; Deary, I.J. Cognitive aging, childhood intelligence, and the use of food supplements: Possible involvement of n-3 fatty acids. Am. J. Clin. Nutr. 2004, 80, 1650–1657. [Google Scholar]
- Anderson, B.M.; Ma, D.W. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009, 8, 33. [Google Scholar] [CrossRef]
- Rauch, F.; Schoenau, E. The developing bone: Slave or master of its cells and molecules? Pediatr. Res. 2001, 50, 309–314. [Google Scholar] [CrossRef]
- Seeman, E. Periosteal bone formation--a neglected determinant of bone strength. N. Engl. J Med. 2003, 349, 320–323. [Google Scholar] [CrossRef]
- Heaney, R.P.; Abrams, S.; wson-Hughes, B.; Looker, A.; Marcus, R.; Matkovic, V.; Weaver, C. Peak bone mass. Osteoporos. Int. 2000, 11, 985–1009. [Google Scholar]
- McGowan, J.A. Bone health and osteoporosis: A report of the surgeon general. US Dep. Health Hum. Serv. 2004.
- Warden, S.J.; Fuchs, R.K.; Castillo, A.B.; Nelson, I.R.; Turner, C.H. Exercise when young provides lifelong benefits to bone structure and strength. J. Bone Miner. Res. 2007, 22, 251–259. [Google Scholar]
- Ammann, P.; Rizzoli, R. Bone strength and its determinants. Osteoporos. Int. 2003, 14, S13–S18. [Google Scholar]
- Liu, D.; Veit, H.P.; Wilson, J.H.; Denbow, D.M. Long-term supplementation of various dietary lipids alters bone mineral content, mechanical properties and histological characteristics of Japanese quail. Poult. Sci. 2003, 82, 831–839. [Google Scholar] [CrossRef]
- Li, Y.; Greiner, R.S.; Salem, N., Jr.; Watkins, B.A. Impact of dietary n-3 FA deficiency on rat bone tissue FA composition. Lipids 2003, 38, 683–686. [Google Scholar] [CrossRef]
- Watkins, B.A.; Li, Y.; Allen, K.G.; Hoffmann, W.E.; Seifert, M.F. Dietary ratio of (n-6)/(n-3) polyunsaturated fatty acids alters the fatty acid composition of bone compartments and biomarkers of bone formation in rats. J. Nutr. 2000, 130, 2274–2284. [Google Scholar]
- Reinwald, S.; Li, Y.; Moriguchi, T.; Salem, N., Jr.; Watkins, B.A. Repletion with (n-3) fatty acids reverses bone structural deficits in (n-3)-deficient rats. J. Nutr. 2004, 134, 388–394. [Google Scholar]
- Lau, B.Y.; Ward, W.E.; Kang, J.X.; Ma, D.W. Fat-1 gene modulates the fatty acid composition of femoral and vertebral phospholipids. Appl. Physiol. Nutr. Metab. 2010, 35, 447–455. [Google Scholar] [CrossRef]
- Lau, B.Y.; Fajardo, V.A.; McMeekin, L.; Sacco, S.M.; Ward, W.E.; Roy, B.D.; Peters, S.J.; Leblanc, P.J. Influence of high-fat diet from differential dietary sources on bone mineral density, bone strength, and bone fatty acid composition in rats. Appl. Physiol. Nutr. Metab. 2010, 35, 598–606. [Google Scholar] [CrossRef]
- Li, Y.; Seifert, M.F.; Lim, S.Y.; Salem, N., Jr.; Watkins, B.A. Bone mineral content is positively correlated to n-3 fatty acids in the femur of growing rats. Br. J. Nutr. 2010, 104, 674–685. [Google Scholar] [CrossRef]
- Al-Nouri, D.M.; Al-Khalifa, A.S. Long-term supplementation of dietary omega-6/omega-3 ratios alters bone marrow fatty acid and biomarkers of bone metabolism in growing rabbits. J. Funct. Foods 2013, 4, 584–593. [Google Scholar] [CrossRef]
- Korotkova, M.; Ohlsson, C.; Hanson, L.A.; Strandvik, B. Dietary n-6:n-3 fatty acid ratio in the perinatal period affects bone parameters in adult female rats. Br. J. Nutr. 2004, 92, 643–648. [Google Scholar] [CrossRef]
- Cohen, S.L.; Ward, W.E. Flaxseed oil and bone development in growing male and female mice. J. Toxicol. Environ. Health A 2005, 68, 1861–1870. [Google Scholar] [CrossRef]
- Weiler, H.A.; Fitzpatrick-Wong, S.C. Modulation of essential (n-6):(n-3) fatty acid ratios alters fatty acid status but not bone mass in piglets. J. Nutr. 2002, 132, 2667–2672. [Google Scholar]
- Lukas, R.; Gigliotti, J.C.; Smith, B.J.; Altman, S.; Tou, J.C. Consumption of different sources of omega-3 polyunsaturated fatty acids by growing female rats affects long bone mass and microarchitecture. Bone 2011, 49, 455–462. [Google Scholar] [CrossRef]
- Baird, H.T.; Eggett, D.L.; Fullmer, S. Varying ratios of omega-6:omega-3 fatty acids on the pre- and postmortem bone mineral density, bone ash, and bone breaking strength of laying chickens. Poult. Sci. 2008, 87, 323–328. [Google Scholar] [CrossRef]
- Macri, E.V.; Gonzales Chaves, M.M.; Rodriguez, P.N.; Mandalunis, P.; Zeni, S.; Lifshitz, F.; Friedman, S.M. High-fat diets affect energy and bone metabolism in growing rats. Eur. J. Nutr. 2012, 51, 399–406. [Google Scholar] [CrossRef]
- Ward, W.E.; Yuan, Y.V.; Cheung, A.M.; Thompson, L.U. Exposure to flaxseed and its purified lignan reduces bone strength in young but not older male rats. J. Toxicol. Environ. Health A 2001, 63, 53–65. [Google Scholar] [CrossRef]
- Green, K.H.; Wong, S.C.; Weiler, H.A. The effect of dietary n-3 long-chain polyunsaturated fatty acids on femur mineral density and biomarkers of bone metabolism in healthy, diabetic and dietary-restricted growing rats. Prostaglandins Leukot. Essent. Fat. Acids 2004, 71, 121–130. [Google Scholar] [CrossRef]
- Kruger, M.C.; Schollum, L.M. Is docosahexaenoic acid more effective than eicosapentaenoic acid for increasing calcium bioavailability? Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 327–334. [Google Scholar] [CrossRef]
- Sirois, I.; Cheung, A.M.; Ward, W.E. Biomechanical bone strength and bone mass in young male and female rats fed a fish oil diet. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 415–421. [Google Scholar] [CrossRef]
- Nielsen, F.H.; Stoecker, B.J. Boron and fish oil have different beneficial effects on strength and trabecular microarchitecture of bone. J. Trace Elem. Med. Biol. 2009, 23, 195–203. [Google Scholar] [CrossRef]
- Lobo, A.R.; Filho, J.M.; Alvares, E.P.; Cocato, M.L.; Colli, C. Effects of dietary lipid composition and inulin-type fructans on mineral bioavailability in growing rats. Nutrition 2009, 25, 216–225. [Google Scholar] [CrossRef]
- Burdge, G.C. Metabolism of alpha-linolenic acid in humans. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 161–168. [Google Scholar] [CrossRef]
- Van, M.G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential effects of prostaglandin derived from omega-6 and omega-3 polyunsaturated fatty acids on COX-2 expression and IL-6 secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef]
- Broughton, K.S.; Wade, J.W. Total fat and (n-3):(n-6) fat ratios influence eicosanoid production in mice. J. Nutr. 2002, 132, 88–94. [Google Scholar]
- Serhan, C.N.; Jain, A.; Marleau, S.; Clish, C.; Kantarci, A.; Behbehani, B.; Colgan, S.P.; Stahl, G.L.; Merched, A.; Petasis, N.A.; et al. Reduced inflammation and tissue damage in transgenic rabbits overexpressing 15-lipoxygenase and endogenous anti-inflammatory lipid mediators. J. Immunol. 2003, 171, 6856–6865. [Google Scholar]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef]
- Kang, J.X. Fat-1 transgenic mice: A new model for omega-3 research. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 263–267. [Google Scholar] [CrossRef]
- Lau, B.Y.; Ward, W.E.; Kang, J.X.; Ma, D.W. Vertebrae of developing fat-1 mice have greater strength and lower n-6/n-3 fatty acid ratio. Exp. Biol. Med. (Maywood) 2009, 234, 632–638. [Google Scholar] [CrossRef]
- Orr, S.K.; Tong, J.Y.; Kang, J.X.; Ma, D.W.; Bazinet, R.P. The fat-1 mouse has brain docosahexaenoic acid levels achievable through fish oil feeding. Neurochem. Res. 2010, 35, 811–819. [Google Scholar] [CrossRef]
- Weiler, H.A.; Kovacs, H.; Nitschmann, E.; Bankovic-Calic, N.; Aukema, H.; Ogborn, M. Feeding flaxseed oil but not secoisolariciresinol diglucoside results in higher bone mass in healthy rats and rats with kidney disease. Prostaglandins Leukot. Essent. Fat. Acids 2007, 76, 269–275. [Google Scholar] [CrossRef]
- Gao, F.; Kiesewetter, D.; Chang, L.; Ma, K.; Bell, J.M.; Rapoport, S.I.; Igarashi, M. Whole-body synthesis-secretion rates of long-chain n-3 PUFAs from circulating unesterified alpha-linolenic acid in unanesthetized rats. J. Lipid Res. 2009, 50, 749–758. [Google Scholar]
- Claassen, N.; Coetzer, H.; van Papendorp, D.H.; Kruger, M.C. Direct determination of saturable and non-saturable calcium uptake in the developing rat duodenum. Lab. Anim. 1995, 29, 438–441. [Google Scholar] [CrossRef]
- Haag, M.; Magada, O.N.; Claassen, N.; Bohmer, L.H.; Kruger, M.C. Omega-3 fatty acids modulate ATPases involved in duodenal Ca absorption. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 423–429. [Google Scholar] [CrossRef]
- Atkinson, T.G.; Barker, H.J.; Meckling-Gill, K.A. Incorporation of long-chain n-3 fatty acids in tissues and enhanced bone marrow cellularity with docosahexaenoic acid feeding in post-weanling Fischer 344 rats. Lipids 1997, 32, 293–302. [Google Scholar] [CrossRef]
- Casado-Diaz, A.; Santiago-Mora, R.; Dorado, G.; Quesada-Gomez, J.M. The omega-6 arachidonic fatty acid, but not the omega-3 fatty acids, inhibits osteoblastogenesis and induces adipogenesis of human mesenchymal stem cells: Potential implication in osteoporosis. Osteoporos. Int. 2013, 24, 1647–1661. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, M.; Alander, C.B.; Choudhary, S.; Pilbeam, C.C.; Raisz, L.G. Effects of prostaglandin E2 on bone in mice in vivo. Prostaglandins Other Lipid Mediat. 2009, 89, 20–25. [Google Scholar] [CrossRef]
- Raisz, L.G.; Alander, C.B.; Simmons, H.A. Effects of prostaglandin E3 and eicosapentaenoic acid on rat bone in organ culture. Prostaglandins 1989, 37, 615–625. [Google Scholar]
- Coetzee, M.; Haag, M.; Kruger, M.C. Effects of arachidonic acid, docosahexaenoic acid, prostaglandin E(2) and parathyroid hormone on osteoprotegerin and RANKL secretion by MC3T3-E1 osteoblast-like cells. J. Nutr. Biochem. 2007, 18, 54–63. [Google Scholar] [CrossRef]
- Fong, L.; Muhlhausler, B.S.; Gibson, R.A.; Xian, C.J. Perinatal maternal dietary supplementation of omega3-fatty acids transiently affects bone marrow microenvironment, osteoblast and osteoclast formation, and bone mass in male offspring. Endocrinology 2012, 153, 2455–2465. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Rahman, M.; Sun, D.; Fernandes, G. Effect of fish oil on bone mineral density in aging C57BL/6 female mice. J. Nutr. Biochem. 2007, 18, 372–379. [Google Scholar] [CrossRef]
- Roqueta-Rivera, M.; Stroud, C.K.; Haschek, W.M.; Akare, S.J.; Segre, M.; Brush, R.S.; Agbaga, M.P.; Anderson, R.E.; Hess, R.A.; Nakamura, M.T. Docosahexaenoic acid supplementation fully restores fertility and spermatogenesis in male delta-6 desaturase-null mice. J. Lipid Res. 2010, 51, 360–367. [Google Scholar] [CrossRef]
- Jee, W.S.; Yao, W. Overview: Animal models of osteopenia and osteoporosis. J. Musculoskelet. Neuronal. Interact. 2001, 1, 193–207. [Google Scholar]
- Ward, W.E.; Piekarz, A.V.; Fonseca, D. Bone mass, bone strength, and their relationship in developing CD-1 mice. Can. J. Physiol. Pharmacol. 2007, 85, 274–279. [Google Scholar] [CrossRef]
- Harvey, N.; Dhanwal, D.; Robinson, S.; Kim, M.; Inskip, H.; Godfrey, K.; Dennison, E.; Calder, P.; Cooper, C. Does maternal long chain polyunsaturated fatty acid status in pregnancy influence the bone health of children? The Southampton Women’s Survey. Osteoporos. Int. 2012, 23, 2359–2367. [Google Scholar] [CrossRef]
- Hogstrom, M.; Nordstrom, P.; Nordstrom, A. n-3 Fatty acids are positively associated with peak bone mineral density and bone accrual in healthy men: The NO2 Study. Am. J. Clin. Nutr. 2007, 85, 803–807. [Google Scholar]
- Damsgaard, C.T.; Molgaard, C.; Matthiessen, J.; Gyldenlove, S.N.; Lauritzen, L. The effects of n-3 long-chain polyunsaturated fatty acids on bone formation and growth factors in adolescent boys. Pediatr. Res. 2012, 71, 713–719. [Google Scholar] [CrossRef]
- Geusens, P.; Dinant, G. Integrating a gender dimension into osteoporosis and fracture risk research. Gend. Med. 2007, 4, S147–S161. [Google Scholar] [CrossRef]
- Decsi, T.; Kennedy, K. Sex-specific differences in essential fatty acid metabolism. Am. J. Clin. Nutr. 2011, 94, 1914S–1919S. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lau, B.Y.Y.; Cohen, D.J.A.; Ward, W.E.; Ma, D.W.L. Investigating the Role of Polyunsaturated Fatty Acids in Bone Development Using Animal Models. Molecules 2013, 18, 14203-14227. https://doi.org/10.3390/molecules181114203
Lau BYY, Cohen DJA, Ward WE, Ma DWL. Investigating the Role of Polyunsaturated Fatty Acids in Bone Development Using Animal Models. Molecules. 2013; 18(11):14203-14227. https://doi.org/10.3390/molecules181114203
Chicago/Turabian StyleLau, Beatrice Y.Y., Daniel J.A. Cohen, Wendy E. Ward, and David W.L. Ma. 2013. "Investigating the Role of Polyunsaturated Fatty Acids in Bone Development Using Animal Models" Molecules 18, no. 11: 14203-14227. https://doi.org/10.3390/molecules181114203