Targeting C-myc G-Quadruplex: Dual Recognition by Aminosugar-Bisbenzimidazoles with Varying Linker Lengths

Abstract
:1. Introduction


2. Results and Discussion
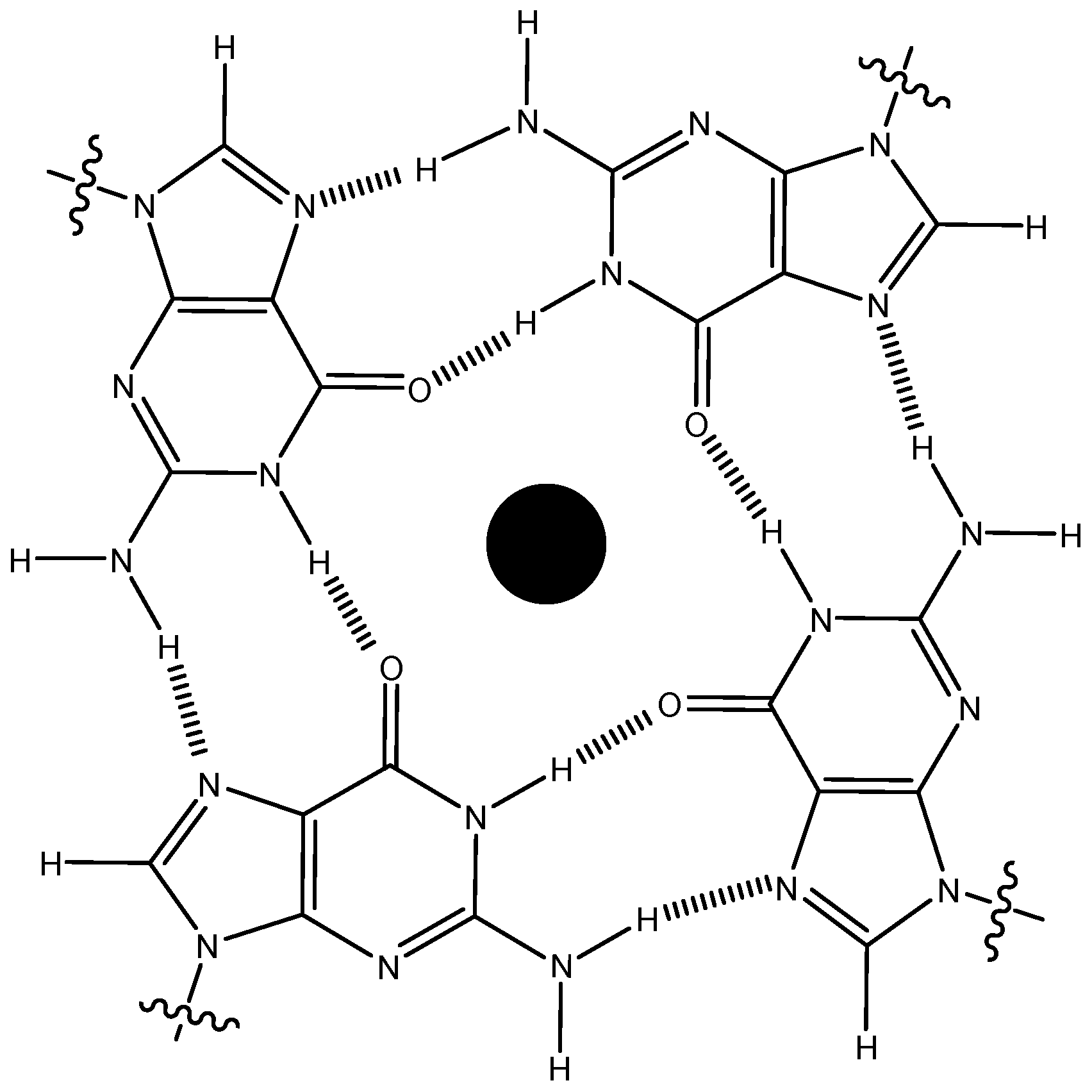
2.1. Selection of the G-Quadruplex

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | DNA sequence (5ʹ-3ʹ) |
|---|---|
| C-myc quadruplex | TGGGGAGGGTGGGGAGGGTGGGGAAGG |
| TFO Active against C-myc duplex a | AGGGAGGGAGGTAAGAAAAAGGG |
| TFO Active against C-myc 1 duplex a | GGAAGGGGTGGGAGGGGTGGGAGGGG |
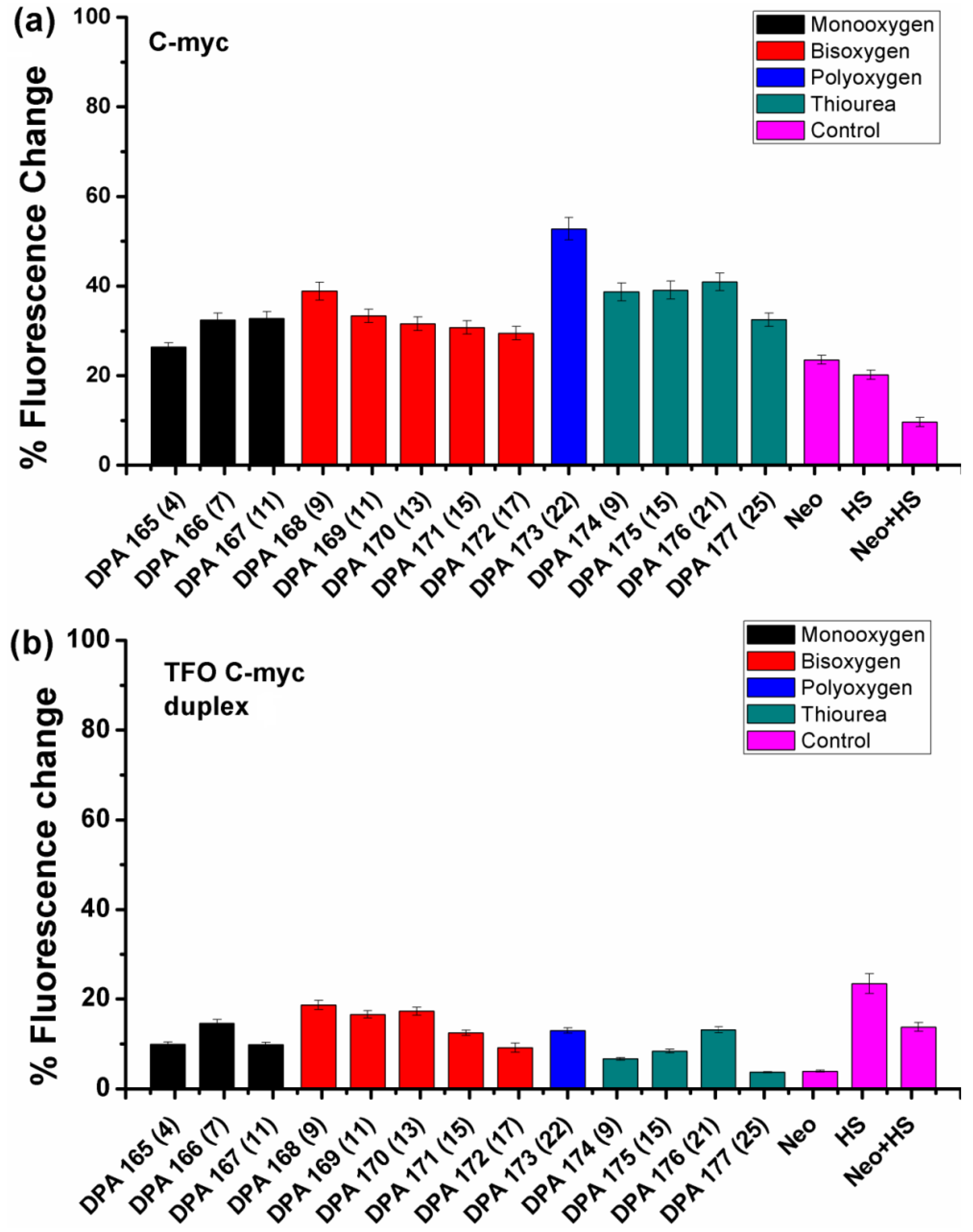
2.2. Fluorescent Intercalator Displacement (FID) Assay

| Nucleic Acid | Ligand | DC50 (µM) |
|---|---|---|
| C-myc | DPA 173 | 0.48 ± 0.04 |
| C-myc | Hoechst 33258 | 1.58 ± 0.01 |
| C-myc | Neomycin | >5.5 |
| C-myc duplex | DPA 173 | 1.07 ± 0.09 |
| C-myc duplex 1 | DPA 173 | 1.22 ± 0.17 |
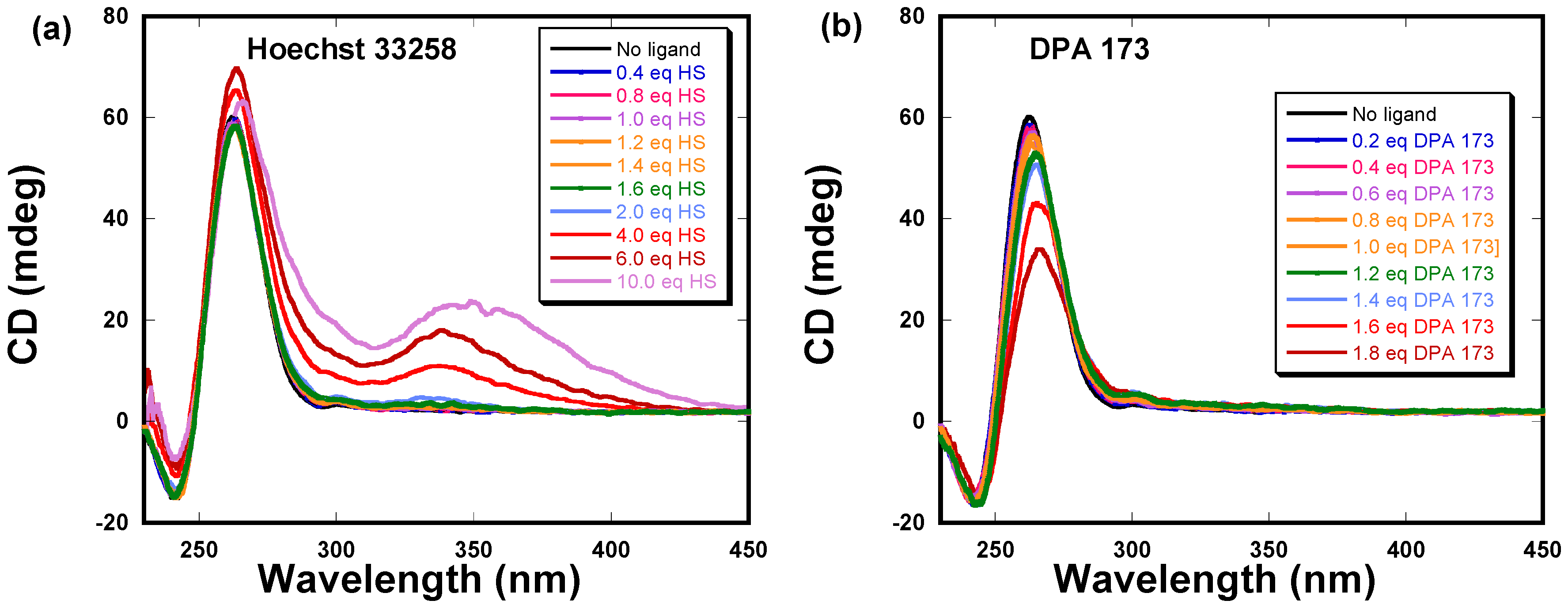
2.3. Circular Dichroism Studies of DPA 173 and Hoechst 33258 Binding to C-myc G-Quadruplex

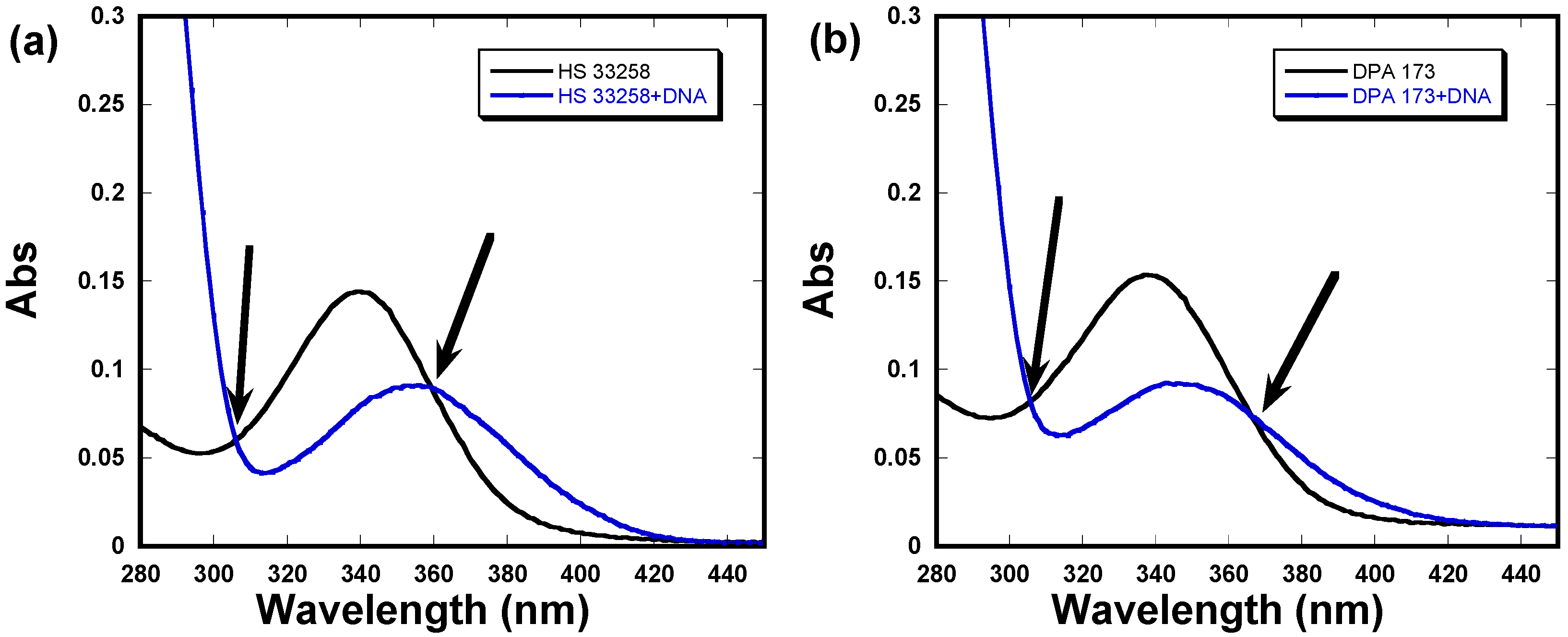
2.4. UV-Vis Absorption Studies

3. Experimental
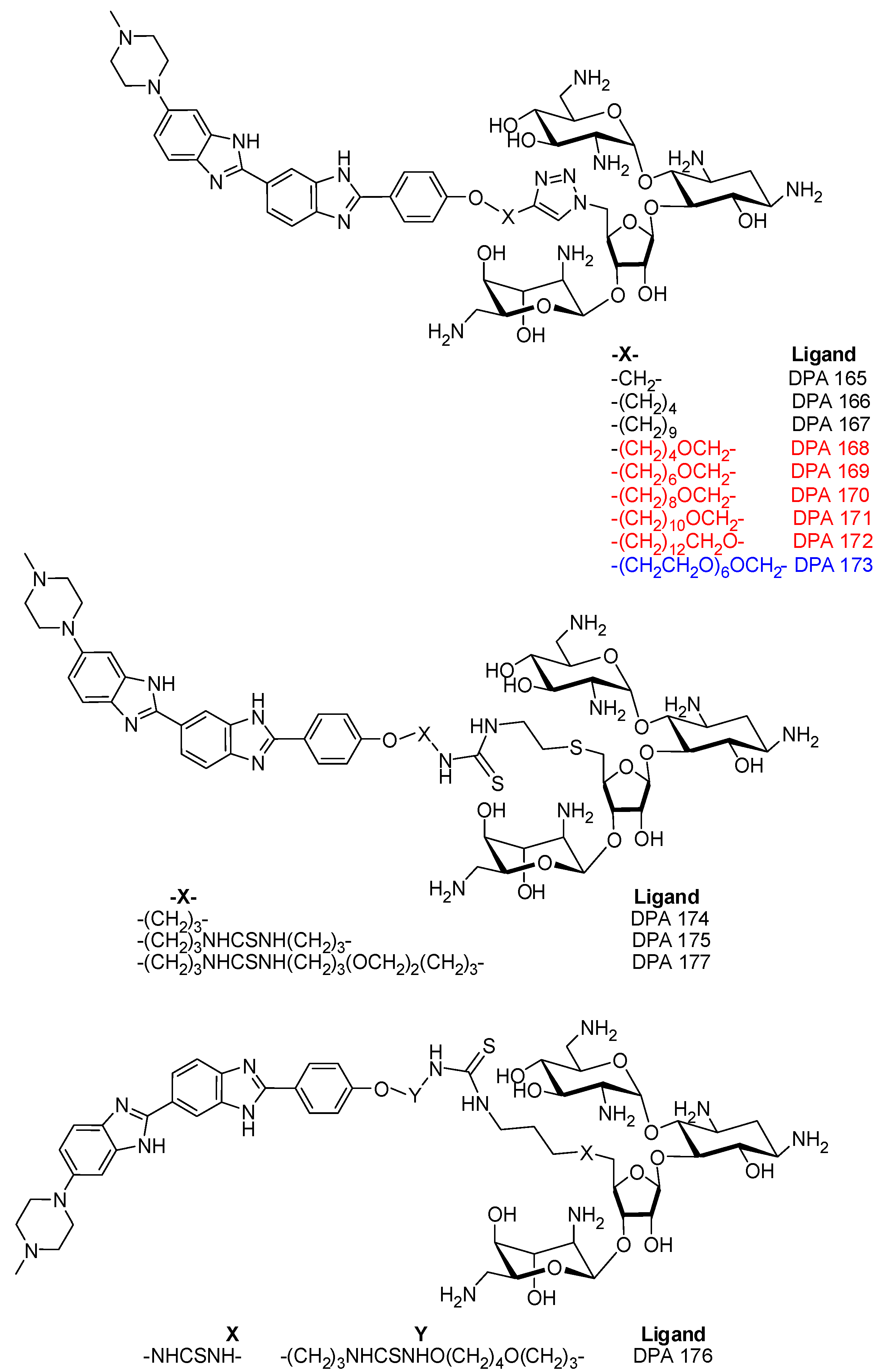
3.1. Synthesis of DPA 165-DPA177
3.2. Nucleic Acids
3.3. Fluorescent Intercalator Displacement (FID) Experiments
3.4. Circular Dichroism (CD) Spectroscopy
3.5. Fluorescence Titration Based FID Experiments
3.6. UV-Vis Experiments
4. Conclusions
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Huppert, J.L.; Balasubramanian, S. G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef]
- Maizels, N. G4 motifs in human genes. Ann. N. Y. Acad. Sci. 2012, 1267, 53–60. [Google Scholar] [CrossRef]
- Kumar, N.; Maiti, S. A thermodynamic overview of naturally occurring intramolecular DNA quadruplexes. Nucleic Acids Res. 2008, 36, 5610–5622. [Google Scholar] [CrossRef]
- Zahler, A.M.; Williamson, J.R.; Cech, T.R.; Prescott, D.M. Inhibition of telomerase by G-quartet DNA structures. Nature 1991, 350, 718–720. [Google Scholar] [CrossRef]
- Ruden, M.; Puri, N. Novel anticancer therapeutics targeting telomerase. Cancer Treat. Rev. 2013, 39, 444–456. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Neidle, S. G-quadruplex nucleic acids as therapeutic targets. Curr. Opin. Chem. Biol. 2009, 13, 345–353. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, topology and structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef]
- Autexier, C.; Lue, N.F. The structure and function of telomerase reverse transcriptase. Annu. Rev. Biochem. 2006, 75, 493–517. [Google Scholar] [CrossRef]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative visualization of DNA G-quadruplex structures in human cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef]
- Lam, E.Y.N.; Beraldi, D.; Tannahill, D.; Balasubramanian, S. G-quadruplex structures are stable and detectable in human genomic DNA. Nat. Commun. 2013. [Google Scholar] [CrossRef]
- Sun, D.; Guo, K.; Shin, Y. Evidence of the formation of G-quadruplex structures in the promoter region of the human vascular endothelial growth factor gene. Nucleic Acids Res. 2011, 39, 1256–1265. [Google Scholar] [CrossRef]
- Telomeric Quadruplex Ligands I: Anthraquinones and acridines. In Therapeutic Applications of Quadruplex Nucleic Acids; Neidle, S. (Ed.) Academic Press: Boston, MA, USA, 2012; pp. 67–91.
- Monchaud, D.; Teulade-Fichou, M.P. A hitchhiker’s guide to G-quadruplex ligands. Org. Biomol. Chem. 2008, 6, 627–636. [Google Scholar] [CrossRef]
- Haq, I.; Trent, J.O.; Chowdhry, B.Z.; Jenkins, T.C. Intercalative G-tetraplex stabilization of telomeric DNA by a cationic porphyrin. J. Am. Chem. Soc. 1999, 121, 1768–1779. [Google Scholar] [CrossRef]
- Jantos, K.; Rodriguez, R.; Ladame, S.; Shirude, P.S.; Balasubramanian, S. Oxazole-based peptide macrocycles: A new class of G-quadruplex binding ligands. J. Am. Chem. Soc. 2006, 128, 13662–13663. [Google Scholar] [CrossRef]
- Kerwin, S.M.; Chen, G.; Kern, J.T.; Thomas, P.W. Perylene diimide G-quadruplex DNA binding selectivity is mediated by ligand aggregation. Bioorg. Med. Chem. Lett. 2002, 12, 447–450. [Google Scholar] [CrossRef]
- Rossetti, L.; Franceschin, M.; Bianco, A.; Ortaggi, G.; Savino, M. Perylene diimides with different side chains are selective in inducing different G-quadruplex DNA structures and in inhibiting telomerase. Bioorg. Med. Chem. Lett. 2002, 12, 2527–2533. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, C.; Yao, P.; Wang, X.; Tan, J.; Li, D.; Ou, T.; Gu, L.; Huang, Z. Disubstituted 1,8-dipyrazolcarbazole derivatives as a new type of C-myc G-quadruplex binding ligands. Bioorg. Med. Chem. 2012, 20, 2829–2836. [Google Scholar] [CrossRef]
- Liao, S.; Zhou, C.; Wu, W.; Ou, T.; Tan, J.; Li, D.; Gu, L.; Huang, Z. 12-n-Methylated 5,6-dihydrobenzo[c]acridine derivatives: A new class of highly selective ligands for C-myc G-quadruplex DNA. Eur. J. Med. Chem. 2012, 53, 52–63. [Google Scholar] [CrossRef]
- Chan, D.S.; Yang, H.; Kwan, M.H.; Cheng, Z.; Lee, P.; Bai, L.; Jiang, Z.; Wong, C.; Fong, W.; Leung, C.; et al. Structure-based optimization of FDA-approved drug methylene blue as a C-myc G-quadruplex DNA stabilizer. Biochimie 2011, 93, 1055–1064. [Google Scholar] [CrossRef]
- Lee, H.; Chan, D.S.; Yang, F.; Lam, H.; Yan, S.; Che, C.; Ma, D.; Leung, C. Identification of natural product fonsecin B as a stabilizing ligand of C-myc G-quadruplex DNA by high-throughput virtual screening. Chem. Commun. 2010, 46, 4680–4682. [Google Scholar] [CrossRef]
- Ma, D.; Ma, V.P.; Chan, D.S.; Leung, K.; Zhong, H.; Leung, C. In silico screening of quadruplex-binding ligands. Methods 2012, 57, 106–114. [Google Scholar] [CrossRef]
- Maiti, S.; Chaudhury, N.K.; Chowdhury, S. Hoechst 33258 binds to G-quadruplex in the promoter region of human C-myc. Biochem. Biophys. Res. Commun. 2003, 310, 505–512. [Google Scholar] [CrossRef]
- Ranjan, N.; Andreasen, K.F.; Kumar, S.; Hyde-Volpe, D.; Arya, D.P. Aminoglycoside binding to oxytricha nova telomeric DNA. Biochemistry 2010, 49, 9891–9903. [Google Scholar] [CrossRef]
- Dominick, P.K.; Keppler, B.R.; Legassie, J.D.; Moon, I.K.; Jarstfer, M.B. Nucleic acid-binding ligands identify new mechanisms to inhibit telomerase. Bioorg. Med. Chem. Lett. 2004, 14, 3467–3471. [Google Scholar] [CrossRef]
- Shaw, N.N.; Xi, H.; Arya, D.P. Molecular recognition of a DNA:RNA hybrid: Sub-nanomolar binding by a neomycin-methidium conjugate. Bioorg. Med. Chem. Lett. 2008, 18, 4142–4145. [Google Scholar] [CrossRef]
- Arya, D.P.; Willis, B. Reaching into the major groove of b-DNA: Synthesis and nucleic acid binding of a neomycin-hoechst 33258 conjugate. J. Am. Chem. Soc. 2003, 125, 12398–12399. [Google Scholar] [CrossRef]
- Kumar, S.; Xue, L.; Arya, D.P. Neomycin-neomycin dimer: An all-carbohydrate scaffold with high affinity for AT-rich DNA duplexes. J. Am. Chem. Soc. 2011, 133, 7361–7375. [Google Scholar] [CrossRef]
- Willis, B.; Arya, D.P. Recognition of b-DNA by neomycin-Hoechst 33258 conjugates. Biochemistry 2006, 45, 10217–10232. [Google Scholar] [CrossRef]
- Xue, L.; Ranjan, N.; Arya, D.P. Synthesis and spectroscopic studies of the aminoglycoside (neomycin)-perylene conjugate binding to human telomeric DNA. Biochemistry 2011, 50, 2838–2849. [Google Scholar] [CrossRef]
- Kumar, S.; Arya, D.P. Recognition of HIV TAR RNA by triazole linked neomycin dimers. Bioorg. Med. Chem. Lett. 2011, 21, 4788–4792. [Google Scholar] [CrossRef]
- Ranjan, N.; Davis, E.; Xue, L.; Arya, D.P. Dual recognition of the human telomeric G-quadruplex by a neomycin-anthraquinone conjugate. Chem. Commun. 2013, 49, 5796–5798. [Google Scholar] [CrossRef]
- Bai, L.; Ho, H.; Ma, D.; Yang, H.; Fu, W.; Jiang, Z. Aminoglycosylation can enhance the G-quadruplex binding activity of epigallocatechin. PLoS One 2013, 8, e53962. [Google Scholar]
- Yuan, L.; Tian, T.; Chen, Y.; Yan, S.; Xing, X.; Zhang, Z.; Zhai, Q.; Xu, L.; Wang, S.; Weng, X.; et al. Existence of G-quadruplex structures in promoter region of oncogenes confirmed by G-quadruplex DNA cross-linking strategy. Sci. Rep. 2013. [Google Scholar] [CrossRef]
- Gonzalez, V.; Hurley, L.H. The C-myc NHE III1: Function and regulation. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 111–129. [Google Scholar] [CrossRef]
- Boger, D.L.; Fink, B.E.; Brunette, S.R.; Tse, W.C.; Hedrick, M.P. A simple, high-resolution method for establishing DNA binding affinity and sequence selectivity. J. Am. Chem. Soc. 2001, 123, 5878–5891. [Google Scholar] [CrossRef]
- Boger, D.L.; Fink, B.E.; Hedrick, M.P. Total synthesis of distamycin A and 2640 analogues: A solution-phase combinatorial approach to the discovery of new, bioactive DNA binding agents and development of a rapid, high-throughput screen for determining relative DNA binding affinity or DNA binding sequence selectivity. J. Am. Chem. Soc. 2000, 122, 6382–6394. [Google Scholar] [CrossRef]
- Monchaud, D.; Allain, C.; Teulade-Fichou, M. Development of a fluorescent intercalator displacement assay (G4-FID) for establishing quadruplex-DNA affinity and selectivity of putative ligands. Bioorg. Med. Chem. Lett. 2006, 16, 4842–4845. [Google Scholar] [CrossRef]
- Xi, H.; Davis, E.; Ranjan, N.; Xue, L.; Hyde-Volpe, D.; Arya, D.P. Thermodynamics of nucleic acid “Shape Readout” by an aminosugar. Biochemistry 2011, 50, 9088–9113. [Google Scholar] [CrossRef]
- Xi, H.; Kumar, S.; Dosen-Micovic, L.; Arya, D.P. Calorimetric and spectroscopic studies of aminoglycoside binding to AT-rich DNA triple helices. Biochimie 2010, 92, 514–529. [Google Scholar] [CrossRef]
- Mergny, J.L.; Li, J.; Lacroix, L.; Amrane, S.; Chaires, J.B. Thermal difference spectra: A specific signature for nucleic acid structures. Nucleic Acids Res. 2005, 33, e138/1–e138/6. [Google Scholar]
- Arya, D.P.; Micovic, L.; Charles, I.; Coffee, R.L., Jr.; Willis, B.; Xue, L. Neomycin binding to watson-hoogsteen (W-H) DNA triplex groove: A model. J. Am. Chem. Soc. 2003, 125, 3733–3744. [Google Scholar] [CrossRef]
- Arya, D.P.; Coffee, R.L., Jr.; Willis, B.; Abramovitch, A.I. Aminoglycoside-nucleic acid interactions: remarkable stabilization of DNA and RNA triple helices by neomycin. J. Am. Chem. Soc. 2001, 123, 5385–5395. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ranjan, N.; Arya, D.P. Targeting C-myc G-Quadruplex: Dual Recognition by Aminosugar-Bisbenzimidazoles with Varying Linker Lengths. Molecules 2013, 18, 14228-14240. https://doi.org/10.3390/molecules181114228
Ranjan N, Arya DP. Targeting C-myc G-Quadruplex: Dual Recognition by Aminosugar-Bisbenzimidazoles with Varying Linker Lengths. Molecules. 2013; 18(11):14228-14240. https://doi.org/10.3390/molecules181114228
Chicago/Turabian StyleRanjan, Nihar, and Dev P. Arya. 2013. "Targeting C-myc G-Quadruplex: Dual Recognition by Aminosugar-Bisbenzimidazoles with Varying Linker Lengths" Molecules 18, no. 11: 14228-14240. https://doi.org/10.3390/molecules181114228