Sodium Valproate Induces Cell Senescence in Human Hepatocarcinoma Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
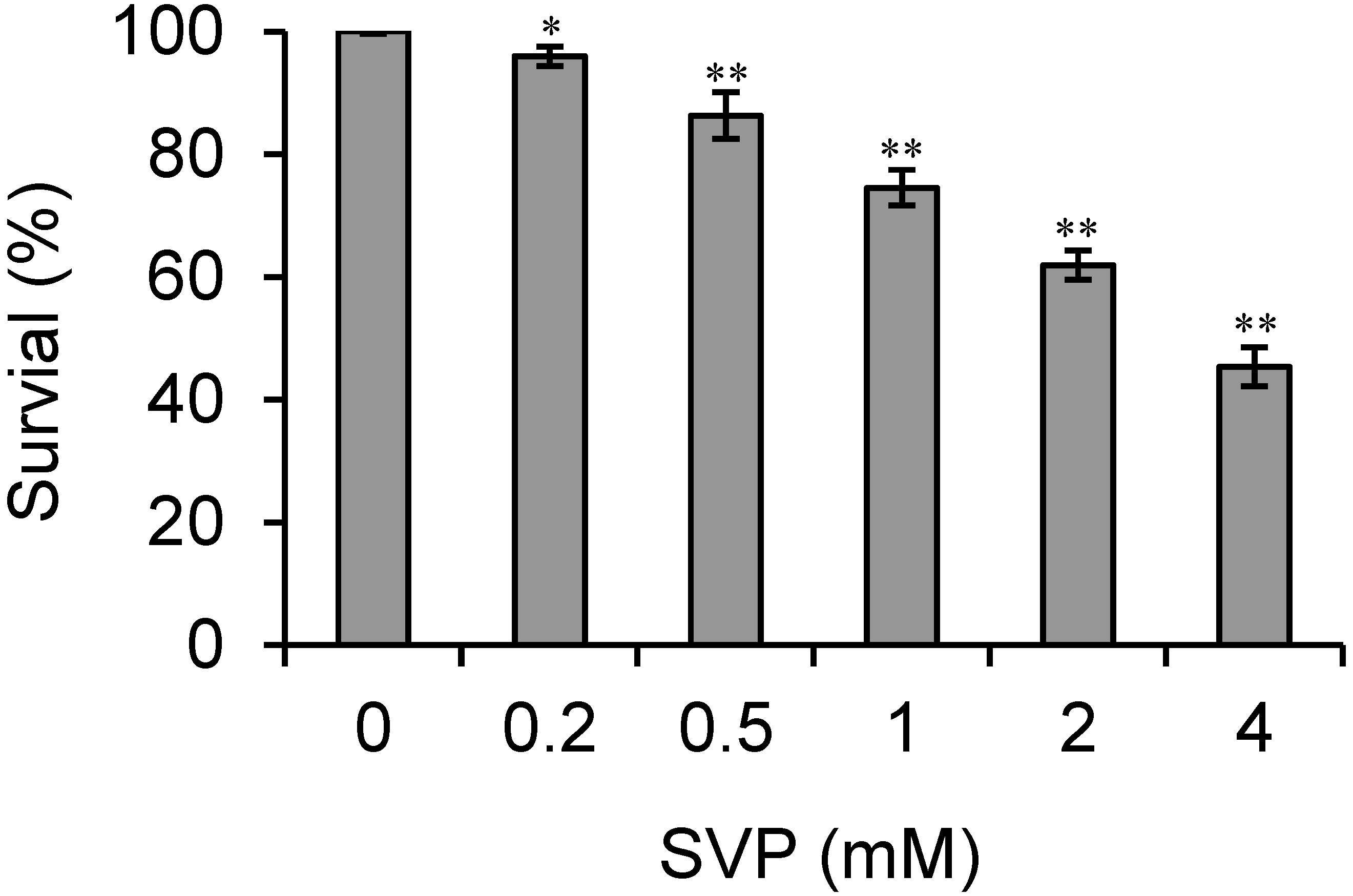
2.1. SVP Inhibits the Proliferation of Bel-7402 Cells

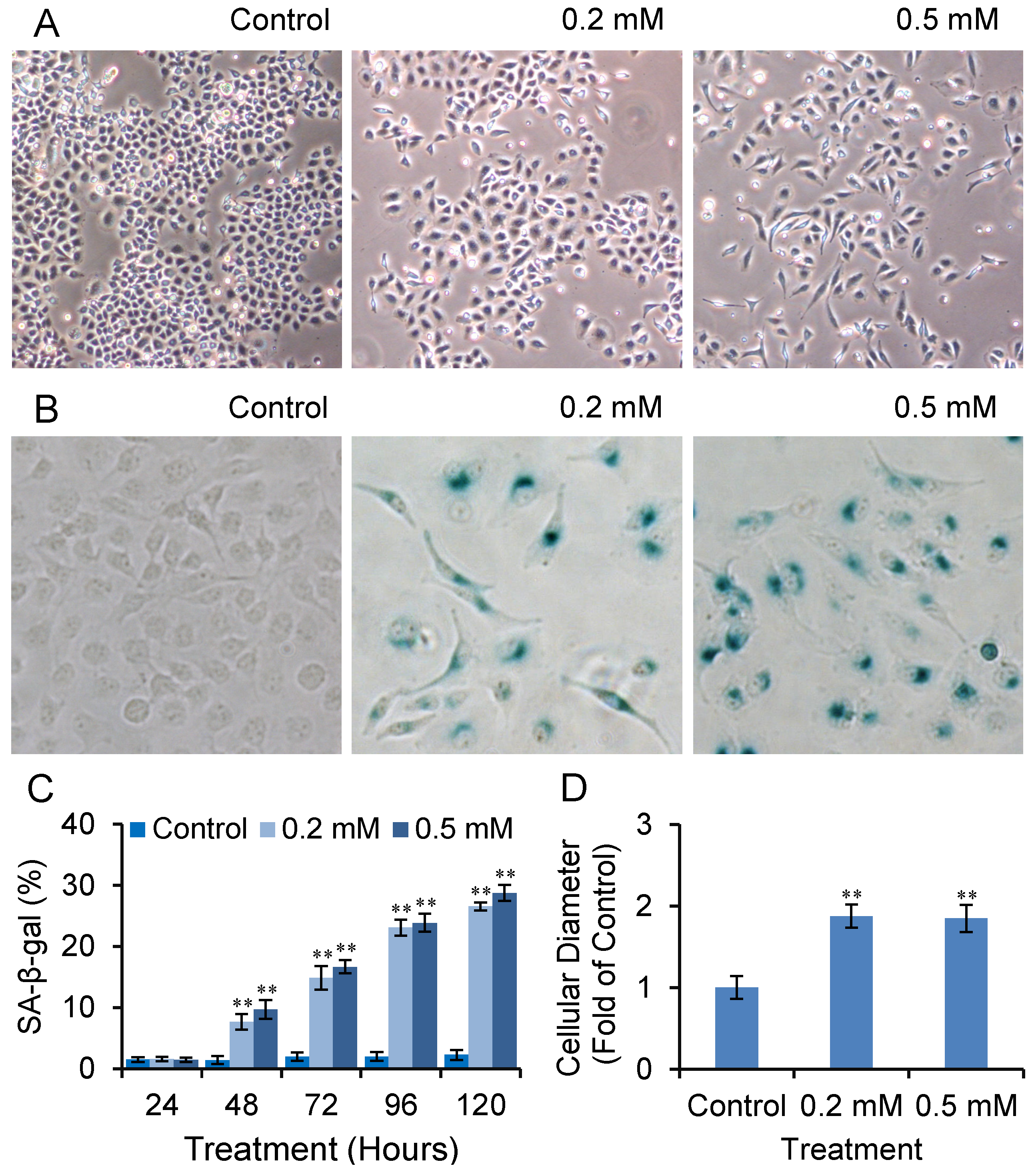
2.2. SVP Activates SA-β-gal in Bel-7402 Cells
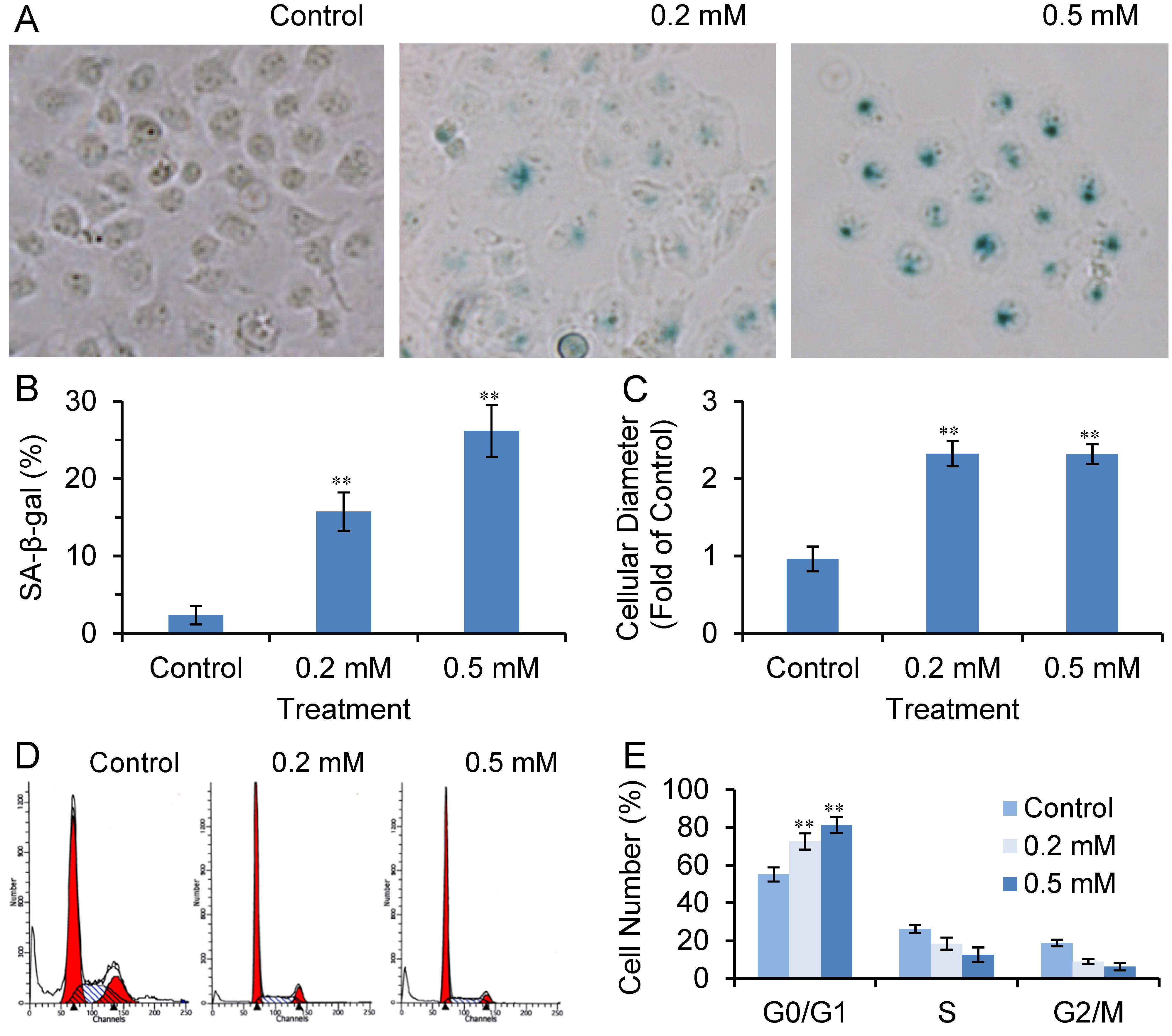
2.3. SVP Arrests Bel-7402 Cells in G0/G1 Phase
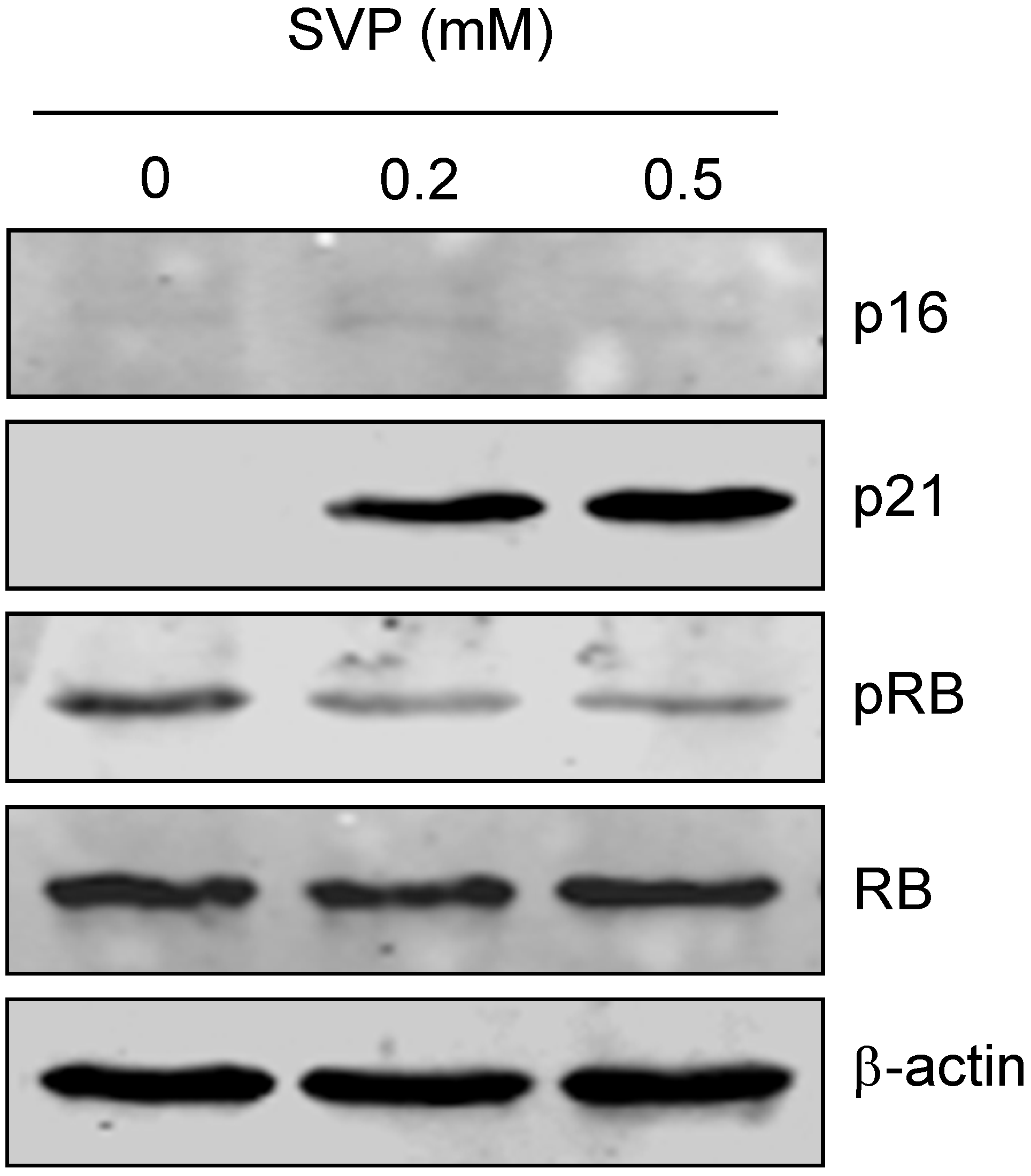
2.4. Effects of SVP on the Expression of Senescence Associated Genes



2.5. SVP Induces Hyperacetylation of Histone H3 and H4
2.6. SVP Induces Cell Senescence in Bel-7404 Cells
2.7. SVP Induces p21 Expression and Inhibits RB Phosphorylation in Bel-7404 Cells



2.8. Discussion
3. Experimental
3.1. Chemicals and Reagents
3.2. Cell Culture
3.3. Cell Proliferation Assay
3.4. Senescence-Associated β-Galactosidase Staining
3.5. Flow Cytometric Analysis
3.6. Western Blot
3.7. Chromatin Immunoprecipitation Assay
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Hagymási, K.; Tulassay, Z. Epidemiology risk factors and molecular pathogenesis of primary liver cancer. Orv. Hetil. 2008, 149, 541–548. [Google Scholar] [CrossRef]
- Rampone, B.; Schiavone, B.; Confuorto, G. Current management of hepatocellular cancer. Curr. Oncol. Rep. 2010, 12, 186–192. [Google Scholar]
- Livraghi, T.; Mäkisalo, H.; Line, P.D. Treatment options in hepatocellular carcinoma today. Scand. J. Surg. 2011, 100, 22–29. [Google Scholar]
- El-Serag, H.B. Current concepts: Hepatocellular carcinoma. N. Engl. J. Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef]
- Herceg, Z.; Paliwal, A. Epigenetic mechanisms in hepatocellular carcinoma: How environmental factors influence the epigenome. Mutat. Res. 2011, 727, 55–61. [Google Scholar] [CrossRef]
- Duong, F.H.; Christen, V.; Lin, S.; Heim, M.H. Hepatitis C virus-induced up-regulation of protein phosphatase 2A inhibits histone modification and DNA damage repair. Hepatology 2010, 51, 741–751. [Google Scholar]
- Yoo, Y.G.; Na, T.Y.; Seo, H.W.; Seong, J.K.; Park, C.K.; Shin, Y.K.; Lee, M.O. Hepatitis B virus X protein induces the expression of MTA1 and HDAC1, which enhances hypoxia signaling in hepatocellular carcinoma cells. Oncogene 2008, 27, 3405–3413. [Google Scholar] [CrossRef]
- Rikimaru, T.; Taketomi, A.; Yamashita, Y.; Shirabe, K.; Hamatsu, T.; Shimada, M.; Maehara, Y. Clinical significance of histone deacetylase 1 expression in patients with hepatocellular carcinoma. Oncology 2007, 72, 69–74. [Google Scholar] [CrossRef]
- Quint, K.; Agaimy, A.; Di Fazio, P.; Montalbano, R.; Steindorf, C.; Jung, R.; Hellerbrand, C.; Hartmann, A.; Sitter, H.; Neureiter, D.; et al. Clinical significance of histone deacetylases 1, 2, 3, and 7: HDAC2 is an independent predictor of survival in HCC. Virchows Arch. 2011, 459, 129–139. [Google Scholar] [CrossRef]
- Göttlicher, M.; Minucci, S.; Zhu, P.; Krämer, O.H.; Schimpf, A.; Giavara, S.; Sleeman, J.P.; Lo Coco, F.; Nervi, C.; Pelicci, P.G.; et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. EMBO J. 2001, 20, 6969–6978. [Google Scholar] [CrossRef]
- Thelen, P.; Schweyer, S.; Hemmerlein, B.; Wuttke, W.; Seseke, F.; Ringert, R.H. Expressional changes after histone deacetylase inhibition by valproic acid in LNCaP human prostate cancer cells. Int. J. Oncol. 2004, 24, 25–31. [Google Scholar]
- Takai, N.; Desmond, J.C.; Kumagai, T.; Gui, D.; Said, J.W.; Whittaker, S.; Miyakawa, I.; Koeffler, H.P. Histone deacetylase inhibitors have a profound antigrowth activity in endometrial cancer cells. Clin. Cancer Res. 2004, 10, 1141–1149. [Google Scholar] [CrossRef]
- Catalano, M.G.; Fortunati, N.; Pugliese, M.; Costantino, L.; Poli, R.; Bosco, O.; Boccuzzi, G. Valproic acid induces apoptosis and cell cycle arrest in poorly differentiated thyroid cancer cells. J. Clin. Endocrinol. Metab. 2005, 90, 1383–1389. [Google Scholar]
- Schwartz, C.; Palissot, V.; Aouali, N.; Wack, S.; Brons, N.H.; Leners, B.; Bosseler, M.; Berchem, G. Valproic acid induces non-apoptotic cell death mechanisms in multiple myeloma cell lines. Int. J. Oncol. 2007, 30, 573–582. [Google Scholar]
- Yagi, Y.; Fushida, S.; Harada, S.; Kinoshita, J.; Makino, I.; Oyama, K.; Tajima, H.; Fujita, H.; Takamura, H.; Ninomiya, I.; et al. Effects of valproic acid on the cell cycle and apoptosis through acetylation of histone and tubulin in a scirrhous gastric cancer cell line. J. Exp. Clin. Cancer Res. 2010, 29, 149. [Google Scholar] [CrossRef]
- Li, X.N.; Shu, Q.; Su, J.M.; Perlaky, L.; Blaney, S.M.; Lau, C.C. Valproic acid induces growth arrest, apoptosis, and senescence in medulloblastomas by increasing histone hyperacetylation and regulating expression of p21Cip1, CDK4, and CMYC. Mol. Cancer Ther. 2005, 4, 1912–1922. [Google Scholar] [CrossRef]
- Gan, C.P.; Hamid, S.; Hor, S.Y.; Zain, R.B.; Ismail, S.M.; Wan Mustafa, W.M.; Teo, S.H.; Saunders, N.; Cheong, S.C. Valproic acid: Growth inhibition of head and neck cancer by induction of terminal differentiation and senescence. Head Neck 2012, 34, 344–353. [Google Scholar] [CrossRef]
- Elknerova, K.; Myslivcova, D.; Lacinova, Z.; Marinov, I.; Uherkova, L.; Stöckbauer, P. Epigenetic modulation of gene expression of human leukemia cell lines—Induction of cell death and senescence. Neoplasma 2011, 58, 35–44. [Google Scholar] [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O.; et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef]
- Chiantore, M.V.; Vannucchi, S.; Mangino, G.; Percario, Z.A.; Affabris, E.; Fiorucci, G.; Romeo, G. Senescence and cell death pathways and their role in cancer therapeutic outcome. Curr. Med. Chem. 2009, 16, 287–300. [Google Scholar] [CrossRef]
- Schmitt, C.A. Cellular senescence and cancer treatment. Biochim. Biophys. Acta 2007, 1775, 5–20. [Google Scholar]
- Gewirtz, D.A.; Holt, S.E.; Elmore, L.W. Accelerated senescence: An emerging role in tumor cell response to chemotherapy and radiation. Biochem. Pharmacol. 2008, 76, 947–957. [Google Scholar] [CrossRef]
- Di, X.; Shiu, R.P.; Newsham, I.F.; Gewirtz, D.A. Apoptosis, autophagy, accelerated senescence and reactive oxygen in the response of human breast tumor cells to adriamycin. Biochem. Pharmacol. 2009, 77, 1139–1150. [Google Scholar] [CrossRef]
- Te Poele, R.H.; Okorokov, A.L.; Jardine, L.; Cummings, J.; Joel, S.P. DNA damage is able to induce senescence in tumor cells in vitro and in vivo. Cancer Res. 2002, 62, 1876–1883. [Google Scholar]
- Han, Z.; Wei, W.; Dunaway, S.; Darnowski, J.W.; Calabresi, P.; Sedivy, J.; Hendrickson, E.A.; Balan, K.V.; Pantazis, P.; Wyche, J.H. Role of p21 in apoptosis and senescence of human colon cancer cells treated with camptothecin. J. Biol. Chem. 2002, 277, 17154–17160. [Google Scholar] [CrossRef]
- Roninson, I.B. Tumor cell senescence in cancer treatment. Cancer Res. 2003, 63, 2705–2715. [Google Scholar]
- Dimri, G.P. What has senescence got to do with cancer. Cancer Cell 2005, 7, 505–512. [Google Scholar] [CrossRef]
- El-Deiry, W.S.; Tokino, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar] [CrossRef]
- Ocker, M.; Schneider-Stock, R. Histone deacetylase inhibitors: Signalling towards p21cip1/waf1. Int. J. Biochem. Cell Biol. 2007, 39, 1367–1374. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, J.H.; Chou, C.W.; Chang, Y.F.; Yeh, S.H.; Chen, C.C. Statins increase p21 through inhibition of histone deacetylase activity and release of promoter-associated HDAC1/2. Cancer Res. 2008, 68, 2375–2383. [Google Scholar] [CrossRef]
- Noh, J.H.; Jung, K.H.; Kim, J.K.; Eun, J.W.; Bae, H.J.; Xie, H.J.; Chang, Y.G.; Kim, M.G.; Park, W.S.; Lee, J.Y.; et al. Aberrant regulation of HDAC2 mediates proliferation of hepatocellular carcinoma cells by deregulating expression of G1/S cell cycle proteins. PLoS One 2011, 6, e28103. [Google Scholar] [CrossRef]
- Gui, C.Y.; Ngo, L.; Xu, W.S.; Richon, V.M.; Marks, P.A. Histone deacetylase (HDAC) inhibitor activation of p21WAF1 involves changes in promoter-associated proteins, including HDAC1. Proc. Natl. Acad. Sci. USA 2004, 101, 1241–1246. [Google Scholar]
- Hu, B.; An, H.M.; Shen, K.P.; Xu, L.; Du, Q.; Deng, S.; Wu, Y. Modified Yi Guan Jian, a Chinese herbal formula, induces anoikis in Bel-7402 human hepatocarcinoma cells in vitro. Oncol. Rep. 2011, 26, 1465–1470. [Google Scholar]
- Hu, B.; An, H.M.; Shen, K.P.; Xu, L.; Du, Q.; Deng, S.; Wu, Y. Liver Yin deficiency tonifying herbal extract induces apoptosis and cell senescence in Bel-7402 human hepatocarcinoma cells. Exp. Ther. Med. 2012, 3, 80–86. [Google Scholar]
- Sample Availability: Not avalable.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
An, H.-M.; Xue, Y.-F.; Shen, Y.-L.; Du, Q.; Hu, B. Sodium Valproate Induces Cell Senescence in Human Hepatocarcinoma Cells. Molecules 2013, 18, 14935-14947. https://doi.org/10.3390/molecules181214935
An H-M, Xue Y-F, Shen Y-L, Du Q, Hu B. Sodium Valproate Induces Cell Senescence in Human Hepatocarcinoma Cells. Molecules. 2013; 18(12):14935-14947. https://doi.org/10.3390/molecules181214935
Chicago/Turabian StyleAn, Hong-Mei, Yong-Fei Xue, Yan-Li Shen, Qin Du, and Bing Hu. 2013. "Sodium Valproate Induces Cell Senescence in Human Hepatocarcinoma Cells" Molecules 18, no. 12: 14935-14947. https://doi.org/10.3390/molecules181214935