Helping Eve Overcome ADAM: G-Quadruplexes in the ADAM-15 Promoter as New Molecular Targets for Breast Cancer Therapeutics

Abstract
:1. Introduction
2. Results and Discussion
2.1. G-Quadruplex (G4) Formation in the ADAM-15 Promoter
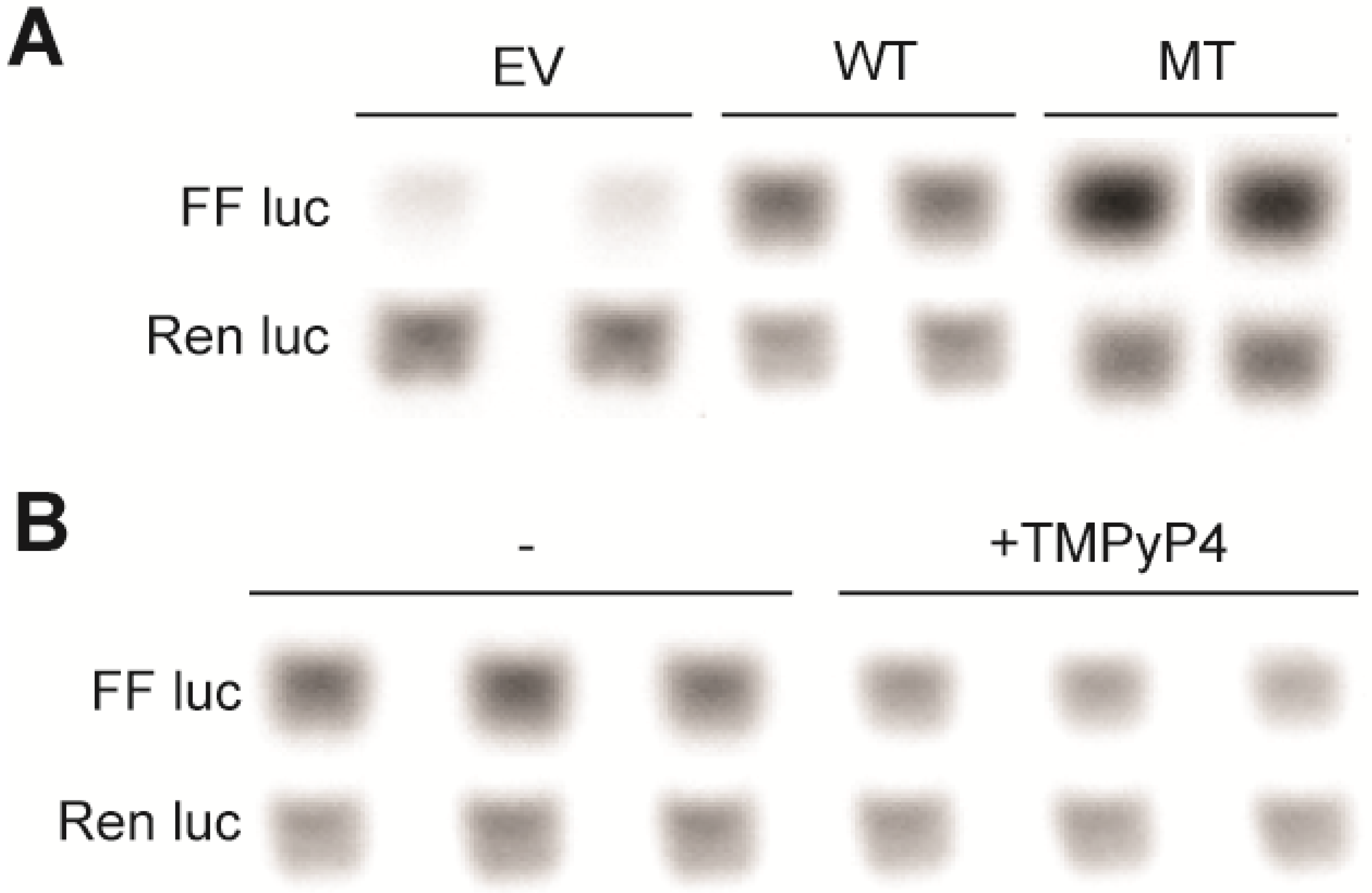
2.2. G4-Mediated Transcriptional Silencing of ADAM-15



2.3. Multiple G4-Isoforms Forming within the ADAM-15 Promoter

2.4. Resolution of Putative G4 Isoforms
3. Experimental
3.1. Chemicals and Oligonucleotides



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | 5'−3' Oligomer Sequence |
|---|---|
| FL | TTGGGGCCGGGTGGGAGGGGGCGGGCCGGGGCGGGGCC |
| 5'-end | TTGGGGCCGGGTGGGAGGGGGCG |
| 5'-mid | CCGGGTGGGAGGGGGCGGGCC |
| 3'-mid | GTGGGAGGGGGCGGGCCGGGGCG |
| 3'-end | GAGGGGGCGGGCCGGGGCGGGGCC |
| a23d3g23 | TTGTTGCCGGGTGGGAGGTGGCGGGCCGGGGCGTTGCC |
| a23 | TTGTTGCCGGGTGGGAGGGGGCGGGCCGGGGCGGGGCC |
| b2 | TTGGGGCCGTGTGGGAGGGGGCGGGCCGGGGCGGGGCC |
| c2 | TTGGGGCCGGGTGTGAGGGGGCGGGCCGGGGCGGGGCC |
| e2 | TTGGGGCCGGGTGGGAGGGGGCGTGCCGGGGCGGGGCC |
| f23 | TTGGGGCCGGGTGGGAGGGGGCGGGCCGGTGCGGGGCC |
| g23 | TTGGGGCCGGGTGGGAGGGGGCGGGCCGGGGCGTTGCC |
3.2. Circular Dichroism and Dimethyl Sulfate Footprinting
3.3. Plasmid Construction
3.4. Cell Culture, Transfection, Luciferase and PCR Assays
4. Conclusions
Conflicts of Interest
References
- Edwards, D.R.; Handsley, M.M.; Pennington, C.J. The adam metalloproteinases. Mol. Asp. Med. 2008, 29, 258–289. [Google Scholar] [CrossRef]
- Wolfsberg, T.G.; Straight, P.D.; Gerena, R.L.; Huovila, A.P.; Primakoff, P.; Myles, D.G.; White, J.M. Adam, a widely distributed and developmentally regulated gene family encoding membrane proteins with a disintegrin and metalloprotease domain. Dev. Biol. 1995, 169, 378–383. [Google Scholar] [CrossRef]
- Borrell-Pages, M.; Rojo, F.; Albanell, J.; Baselga, J.; Arribas, J. Tace is required for the activation of the egfr by tgf-alpha in tumors. EMBO J. 2003, 22, 1114–1124. [Google Scholar] [CrossRef]
- Kenny, P.A.; Bissell, M.J. Targeting tace-dependent egfr ligand shedding in breast cancer. J. Clin. Investig. 2007, 117, 337–345. [Google Scholar] [CrossRef]
- Moss, M.L.; Stoeck, A.; Yan, W.; Dempsey, P.J. Adam10 as a target for anti-cancer therapy. Curr. Pharm. Biotechnol. 2008, 9, 2–8. [Google Scholar] [CrossRef]
- Eto, K.; Huet, C.; Tarui, T.; Kupriyanov, S.; Liu, H.Z.; Puzon-McLaughlin, W.; Zhang, X.P.; Sheppard, D.; Engvall, E.; Takada, Y. Functional classification of adams based on a conserved motif for binding to integrin alpha 9beta 1: Implications for sperm-egg binding and other cell interactions. J. Biol. Chem. 2002, 277, 17804–17810. [Google Scholar] [CrossRef]
- Evans, J.P. Fertilin beta and other adams as integrin ligands: Insights into cell adhesion and fertilization. Bioessays 2001, 23, 628–639. [Google Scholar] [CrossRef]
- Bohm, B.B.; Aigner, T.; Roy, B.; Brodie, T.A.; Blobel, C.P.; Burkhardt, H. Homeostatic effects of the metalloproteinase disintegrin adam15 in degenerative cartilage remodeling. Arthritis Rheum. 2005, 52, 1100–1109. [Google Scholar] [CrossRef]
- Horiuchi, K.; Weskamp, G.; Lum, L.; Hammes, H.P.; Cai, H.; Brodie, T.A.; Ludwig, T.; Chiusaroli, R.; Baron, R.; Preissner, K.T.; et al. Potential role for adam15 in pathological neovascularization in mice. Mol. Cell. Biol. 2003, 23, 5614–5624. [Google Scholar] [CrossRef]
- Kuefer, R.; Day, K.C.; Kleer, C.G.; Sabel, M.S.; Hofer, M.D.; Varambally, S.; Zorn, C.S.; Chinnaiyan, A.M.; Rubin, M.A.; Day, M.L. Adam15 disintegrin is associated with aggressive prostate and breast cancer disease. Neoplasia 2006, 8, 319–329. [Google Scholar] [CrossRef]
- Najy, A.J.; Day, K.C.; Day, M.L. The ectodomain shedding of e-cadherin by adam15 supports erbb receptor activation. J. Biol. Chem. 2008, 283, 18393–18401. [Google Scholar] [CrossRef]
- Schutz, A.; Hartig, W.; Wobus, M.; Grosche, J.; Wittekind, C.; Aust, G. Expression of adam15 in lung carcinomas. Virchows Arch. 2005, 446, 421–429. [Google Scholar] [CrossRef]
- Lucas, N.; Day, M.L. The role of the disintegrin metalloproteinase adam15 in prostate cancer progression. J. Cell. Biochem. 2009, 106, 967–974. [Google Scholar] [CrossRef]
- Witters, L.; Scherle, P.; Friedman, S.; Fridman, J.; Caulder, E.; Newton, R.; Lipton, A. Synergistic inhibition with a dual epidermal growth factor receptor/her-2/neu tyrosine kinase inhibitor and a disintegrin and metalloprotease inhibitor. Cancer Res. 2008, 68, 7083–7089. [Google Scholar]
- Khammas, H.; Bowen, T.; Williams, J.D.; Phillips, A.O.; Steadman, R.; Martin, J. Characterisation of the human adam15 promoter. Nephron Exp. Nephrol. 2011, 118, e27–e38. [Google Scholar]
- Verma, A.; Halder, K.; Halder, R.; Yadav, V.K.; Rawal, P.; Thakur, R.K.; Mohd, F.; Sharma, A.; Chowdhury, S. Genome-wide computational and expression analyses reveal g-quadruplex DNA motifs as conserved cis-regulatory elements in human and related species. J. Med. Chem. 2008, 51, 5641–5649. [Google Scholar] [CrossRef]
- Eddy, J.; Maizels, N. Selection for the g4 DNA motif at the 5' end of human genes. Mol. Carcinog. 2009, 48, 319–325. [Google Scholar] [CrossRef]
- Brooks, T.A.; Hurley, L.H. The role of supercoiling in transcriptional control of myc and its importance in molecular therapeutics. Nat. Rev. 2009, 9, 849–861. [Google Scholar]
- Agarwala, P.; Pandey, S.; Mapa, K.; Maiti, S. The g-quadruplex augments translation in the 5’ untranslated region of transforming growth factor beta2. Biochemistry 2013, 52, 1528–1538. [Google Scholar] [CrossRef]
- Bochman, M.L.; Paeschke, K.; Zakian, V.A. DNA secondary structures: Stability and function of g-quadruplex structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar] [CrossRef]
- Marcel, V.; Tran, P.L.; Sagne, C.; Martel-Planche, G.; Vaslin, L.; Teulade-Fichou, M.P.; Hall, J.; Mergny, J.L.; Hainaut, P.; van Dyck, E. G-quadruplex structures in tp53 intron 3: Role in alternative splicing and in production of p53 mrna isoforms. Carcinogenesis 2011, 32, 271–278. [Google Scholar] [CrossRef]
- Millevoi, S.; Moine, H.; Vagner, S. G-quadruplexes in rna biology. Wiley Interdiscip. Rev. RNA 2012, 3, 495–507. [Google Scholar] [CrossRef]
- Qin, Y.; Hurley, L.H. Structures, folding patterns, and functions of intramolecular DNA g-quadruplexes found in eukaryotic promoter regions. Biochimie 2008, 90, 1149–1171. [Google Scholar] [CrossRef]
- IDT Biophysics: UV Spectrum of DNA. Available online: http://biophysics.idtdna.com/UVSpectrum.html/ (accessed on 1 October 2013).
- Qin, Y.; Fortin, J.S.; Tye, D.; Gleason-Guzman, M.; Brooks, T.A.; Hurley, L.H. Molecular cloning of the human platelet-derived growth factor receptor β (pdgfr-β) promoter and drug targeting of the g-quadruplex-forming region to repress pdgfr-β expression. Biochemistry 2010, 49, 4208–4219. [Google Scholar] [CrossRef]
- Maekawa, T.; Sano, Y.; Shinagawa, T.; Rahman, Z.; Sakuma, T.; Nomura, S.; Licht, J.D.; Ishii, S. Atf-2 controls transcription of maspin and gadd45 alpha genes independently from p53 to suppress mammary tumors. Oncogene 2008, 27, 1045–1054. [Google Scholar]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting g-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef]
- Sjostrom, J.; Blomqvist, C.; von Boguslawski, K.; Bengtsson, N.O.; Mjaaland, I.; Malmstrom, P.; Ostenstadt, B.; Wist, E.; Valvere, V.; Takayama, S.; et al. The predictive value of bcl-2, bax, bcl-xl, bag-1, fas, and fasl for chemotherapy response in advanced breast cancer. Clin. Cancer Res. 2002, 8, 811–816. [Google Scholar]
- Eddy, J.; Maizels, N. Gene function correlates with potential for g4 DNA formation in the human genome. Nucleic Acids Res. 2006, 34, 3887–3896. [Google Scholar] [CrossRef]
- MacLeod, M.C. Identification of a DNA structural motif that includes the binding sites for sp1, p53 and ga-binding protein. Nucleic Acids Res. 1993, 21, 1439–1447. [Google Scholar] [CrossRef]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative visualization of DNA g-quadruplex structures in human cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef]
- Lam, E.Y.; Beraldi, D.; Tannahill, D.; Balasubramanian, S. G-quadruplex structures are stable and detectable in human genomic DNA. Nat. Commun. 2013. [Google Scholar] [CrossRef]
- Muller, S.; Kumari, S.; Rodriguez, R.; Balasubramanian, S. Small-molecule-mediated g-quadruplex isolation from human cells. Nat. Chem. 2010, 2, 1095–1098. [Google Scholar] [CrossRef]
- Brooks, T.A.; Kendrick, S.; Hurley, L. Making sense of g-quadruplex and i-motif functions in oncogene promoters. FEBS J. 2010, 277, 3459–3469. [Google Scholar] [CrossRef]
- Eto, K.; Puzon-McLaughlin, W.; Sheppard, D.; Sehara-Fujisawa, A.; Zhang, X.P.; Takada, Y. Rgd-independent binding of integrin alpha9beta1 to the adam-12 and -15 disintegrin domains mediates cell-cell interaction. J. Biol. Chem. 2000, 275, 34922–34930. [Google Scholar]
- Nath, D.; Slocombe, P.M.; Stephens, P.E.; Warn, A.; Hutchinson, G.R.; Yamada, K.M.; Docherty, A.J.; Murphy, G. Interaction of metargidin (adam-15) with alphavbeta3 and alpha5beta1 integrins on different haemopoietic cells. J. Cell Sci. 1999, 112, 579–587. [Google Scholar]
- Trochon-Joseph, V.; Martel-Renoir, D.; Mir, L.M.; Thomaidis, A.; Opolon, P.; Connault, E.; Li, H.; Grenet, C.; Fauvel-Lafeve, F.; Soria, J.; et al. Evidence of antiangiogenic and antimetastatic activities of the recombinant disintegrin domain of metargidin. Cancer Res. 2004, 64, 2062–2069. [Google Scholar] [CrossRef]
- Zhang, X.P.; Kamata, T.; Yokoyama, K.; Puzon-McLaughlin, W.; Takada, Y. Specific interaction of the recombinant disintegrin-like domain of mdc-15 (metargidin, adam-15) with integrin alphavbeta3. J. Biol. Chem. 1998, 273, 7345–7350. [Google Scholar]
- Ham, C.; Levkau, B.; Raines, E.W.; Herren, B. Adam15 is an adherens junction molecule whose surface expression can be driven by ve-cadherin. Exp. Cell Res. 2002, 279, 239–247. [Google Scholar] [CrossRef]
- Najy, A.J.; Day, K.C.; Day, M.L. Adam15 supports prostate cancer metastasis by modulating tumor cell-endothelial cell interaction. Cancer Res. 2008, 68, 1092–1099. [Google Scholar] [CrossRef]
- Nieman, M.T.; Prudoff, R.S.; Johnson, K.R.; Wheelock, M.J. N-cadherin promotes motility in human breast cancer cells regardless of their e-cadherin expression. J. Cell Biol. 1999, 147, 631–644. [Google Scholar] [CrossRef]
- Gofuku, J.; Shiozaki, H.; Doki, Y.; Inoue, M.; Hirao, M.; Fukuchi, N.; Monden, M. Characterization of soluble e-cadherin as a disease marker in gastric cancer patients. Br. J. Cancer 1998, 78, 1095–1101. [Google Scholar] [CrossRef]
- Kuefer, R.; Hofer, M.D.; Gschwend, J.E.; Pienta, K.J.; Sanda, M.G.; Chinnaiyan, A.M.; Rubin, M.A.; Day, M.L. The role of an 80 kda fragment of e-cadherin in the metastatic progression of prostate cancer. Clin. Cancer Res. 2003, 9, 6447–6452. [Google Scholar]
- Bedard, P.L.; Piccart-Gebhart, M.J. Current paradigms for the use of her2-targeted therapy in early-stage breast cancer. Clin. Breast Cancer 2008, 8, S157–S165. [Google Scholar] [CrossRef]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of her2-overexpressing metastatic breast cancer. J. Clin. Oncol. 2002, 20, 719–726. [Google Scholar] [CrossRef]
- Dieras, V.; Vincent-Salomon, A.; Degeorges, A.; Beuzeboc, P.; Mignot, L.; de Cremoux, P. trastuzumab (herceptin) and breast cancer: Mechanisms of resistance. Bull. Cancer 2007, 94, 259–266. [Google Scholar]
- Mariani, G.; Fasolo, A.; de Benedictis, E.; Gianni, L. Trastuzumab as adjuvant systemic therapy for her2-positive breast cancer. Nat. Clin. Pract. 2009, 6, 93–104. [Google Scholar]
- Sample Availability: Samples of the compounds TMPyP4 are available from CalBioChem/Millipore.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brown, R.V.; Gaerig, V.C.; Simmons, T.; Brooks, T.A. Helping Eve Overcome ADAM: G-Quadruplexes in the ADAM-15 Promoter as New Molecular Targets for Breast Cancer Therapeutics. Molecules 2013, 18, 15019-15034. https://doi.org/10.3390/molecules181215019
Brown RV, Gaerig VC, Simmons T, Brooks TA. Helping Eve Overcome ADAM: G-Quadruplexes in the ADAM-15 Promoter as New Molecular Targets for Breast Cancer Therapeutics. Molecules. 2013; 18(12):15019-15034. https://doi.org/10.3390/molecules181215019
Chicago/Turabian StyleBrown, Robert V., Vanessa C. Gaerig, Taesha Simmons, and Tracy A. Brooks. 2013. "Helping Eve Overcome ADAM: G-Quadruplexes in the ADAM-15 Promoter as New Molecular Targets for Breast Cancer Therapeutics" Molecules 18, no. 12: 15019-15034. https://doi.org/10.3390/molecules181215019