Madecassoside Inhibits Melanin Synthesis by Blocking Ultraviolet-Induced Inflammation

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
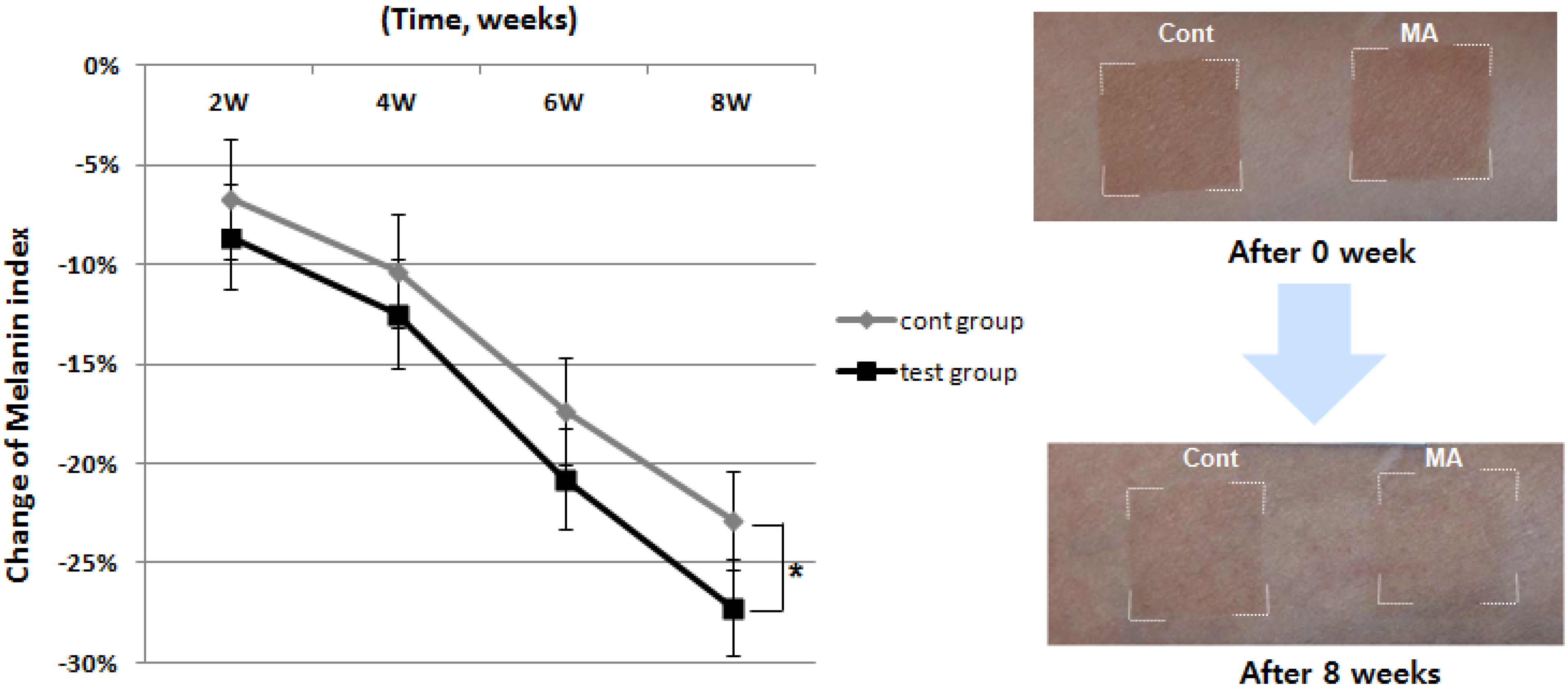{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
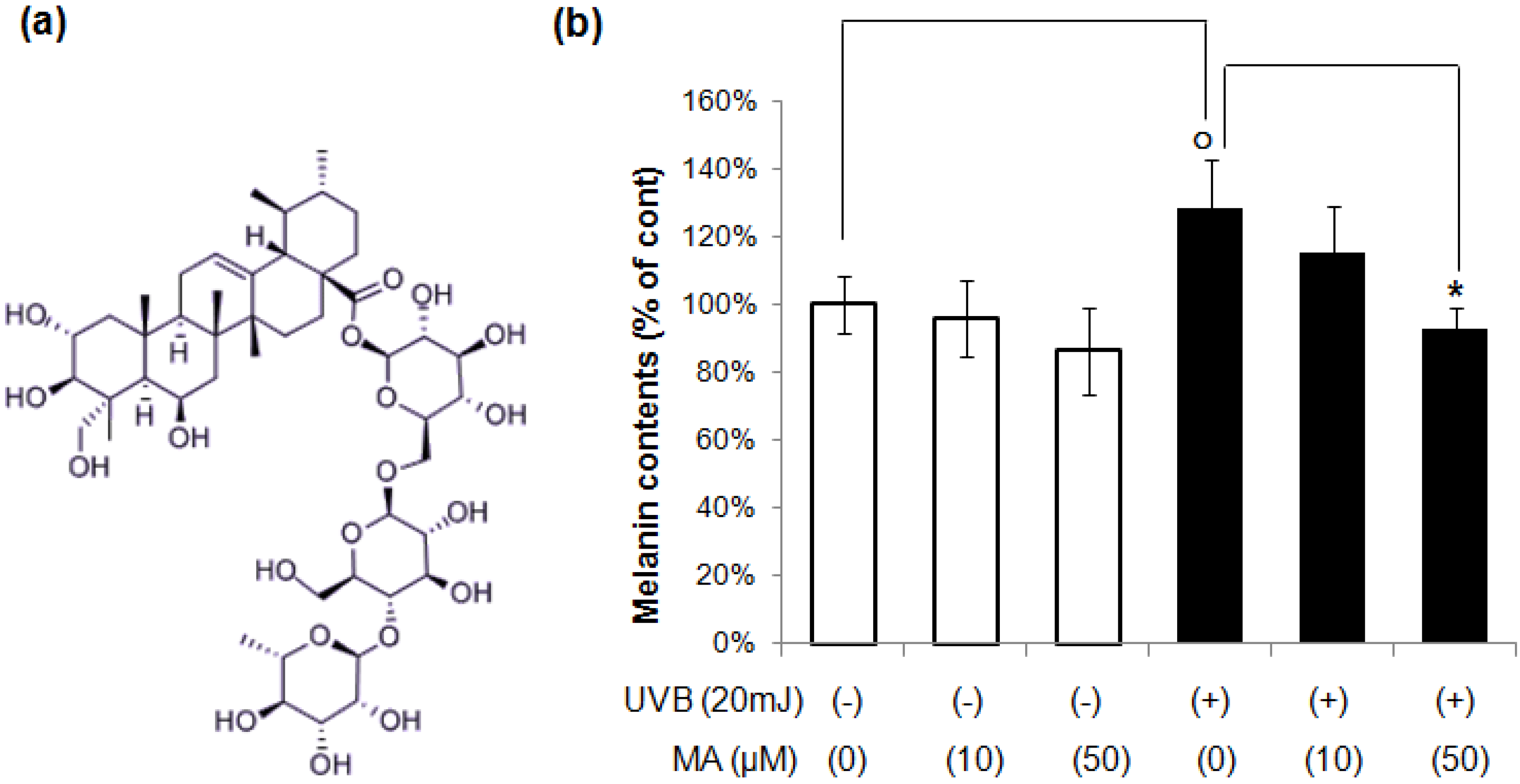
2.1. Effect of MA on UVR-Induced Melanogenesis in Keratinocyte/Melanocytes Co-Cultures

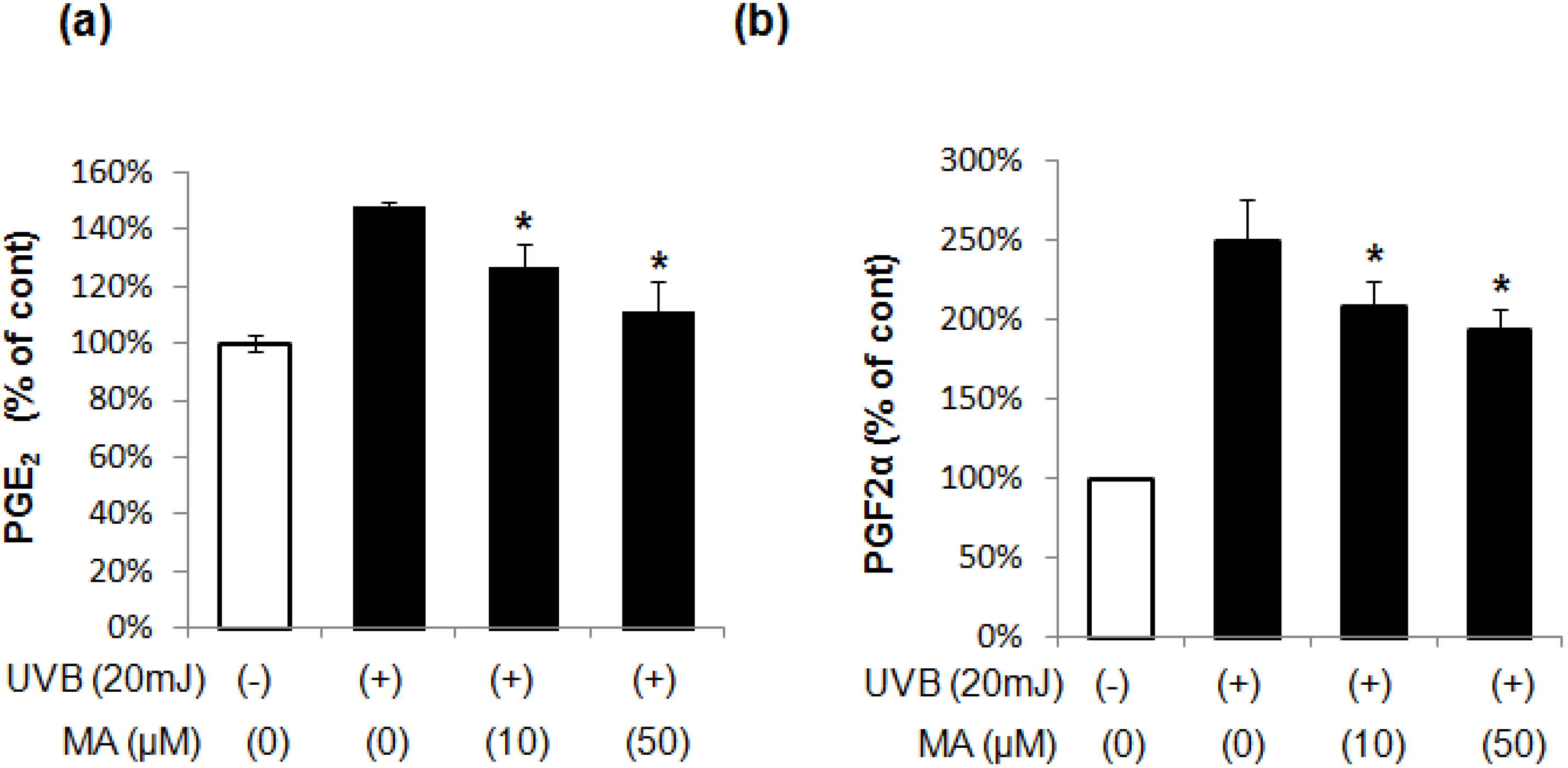
2.2. Effect of MA on PGE2 and PGF2α Production in Keratinocytes
2.3. Effect of MA on COX-2 and PAR-2 Expression in Keratinocytes


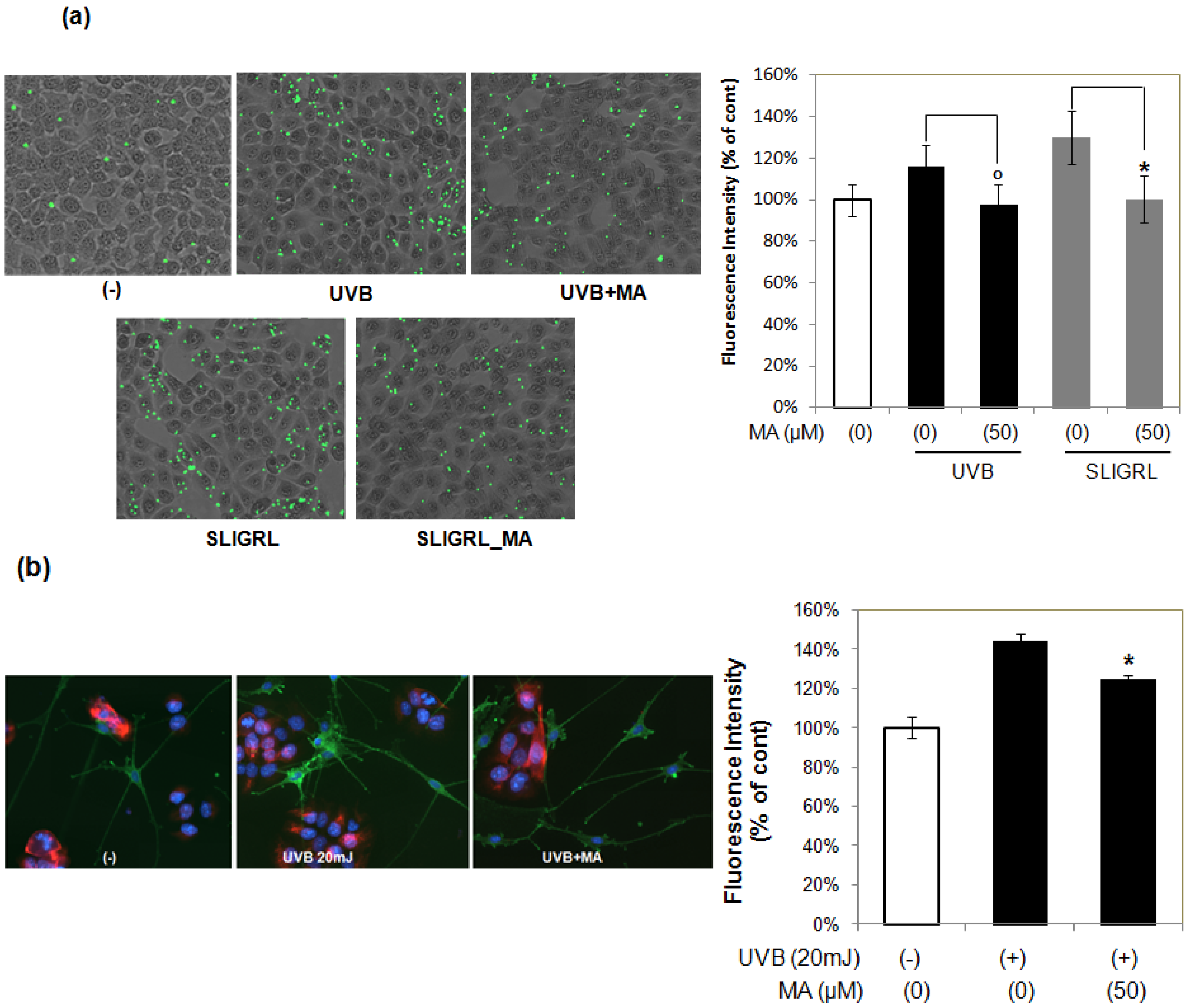
2.4. Effect of MA on Phogocytosis

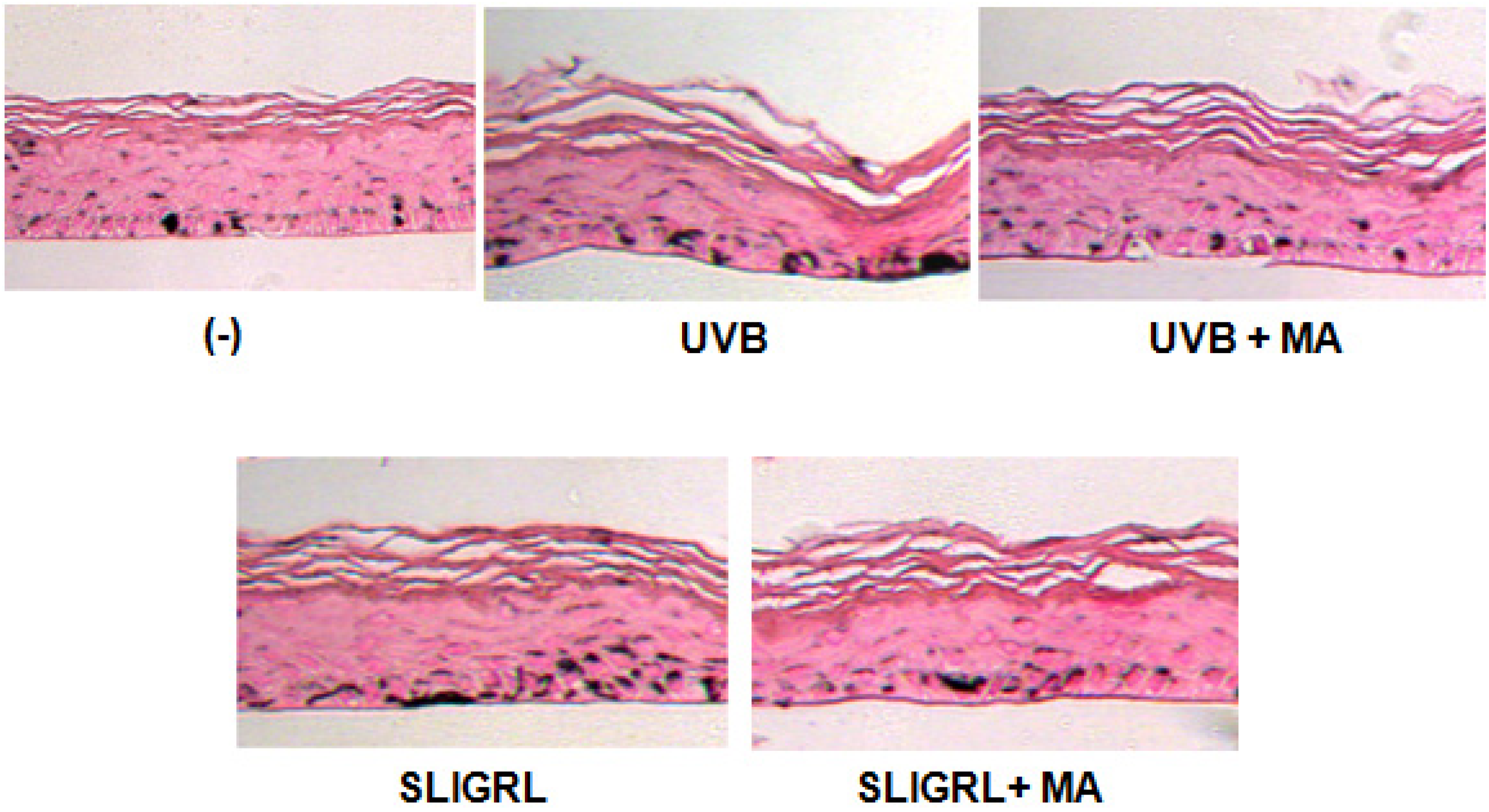
2.5. Effect of MA on Melanogenesis in Reconstructed Epidermis

2.6. Effect of MA on UVB-Induced Tannin

3. Experimental
3.1. Materials
3.2. Cell and Cell Culture
3.3. UV Irradiation
3.4. Co-Culture
3.5. Melanin Assay
3.6. Lipid Mediator of Inflammation Assay
3.7. Western Blot Analysis
3.8. Phagocytosis Assay
3.9. Immunofluorescence Staining
3.10. Histochemistry of Reconstructed Epidermis
3.11. Clinical Study of MA on UV-Induced Tanning Skin
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yamaguchi, Y.; Brenner, M.; Hearing, V.J. The regulation of skin pigmentation. J. Biol. Chem. 2007, 282, 27557–27561. [Google Scholar] [CrossRef]
- The Pigmentary System: Physiology and Pathophysiology; Norlund, J.J.; Boissy, R.E.; Hearing, V.J. (Eds.) Oxford University Press: New York, NY, USA, 1998; pp. 909–911.
- Kang-Rotondo, C.H.; Miller, C.C.; Morrison, A.R.; Pentland, A.P. Enhanced keratinocyte prostaglandin synthesis after UV injury is due to increased phospholipase activity. Am. J. Physiol. 1993, 264, 396–401. [Google Scholar]
- Scott, G.; Jacobs, S.; Leopardi, S.; Anthony, F.A.; Learn, D.; Malaviya, R.; Pentland, A. Effects of PGF2alpha on human melanocytes and regulation of the FP receptor by ultraviolet radiation. Exp. Cell Res. 2005, 304, 407–416. [Google Scholar] [CrossRef]
- Scott, G.; Fricke, A.; Fender, A.; McClelland, L.; Jacobs, S. Prostaglandin E2 regulates melanocyte dendrite formation through activation of PKCzeta. Exp. Cell Res. 2007, 313, 3840–3850. [Google Scholar] [CrossRef]
- Derian, C.K.; Eckardt, A.J.; Andrade-Gordon, P. Differential regulation of human keratinocyte growth and differentiation by a novel family of protease-activated receptors. Cell. Growth Differ. 1997, 8, 743–749. [Google Scholar]
- Seiberg, M.; Paine, C.; Sharlow, E.; Andrade-Gordon, P.; Costanzo, M.; Eisinger, M.; Shapiro, S.S. The protease-activated receptor 2 regulates pigmentation via keratinocyte-melanocyte interactions. Exp. Cell Res. 2000, 254, 25–32. [Google Scholar] [CrossRef]
- Seiberg, M. Keratinocyte-melanocyte interactions during melanosome transfer. Pigment Cell Res. 2001, 14, 236–242. [Google Scholar]
- Scott, G.; Deng, A.; Rodriguez-Burford, C.; Seiberg, M.; Han, R.; Babiarz, L.; Grizzle, W.; Bell, W.; Pentland, A. Protease-activated receptor 2, a receptor involved in melanosome transfer, is upregulated in human skin by ultraviolet irradiation. J. Invest. Dermatol. 2001, 117, 1412–1420. [Google Scholar] [CrossRef]
- Chambers, L.S.; Black, J.L.; Ge, Q.; Carlin, S.M.; Au, W.W.; Poniris, M.; Thompson, J.; Johnson, P.R.; Burgess, J.K. PAR-2 activation, PGE2, and COX-2 in human asthmatic and nonasthmatic airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, 619–627. [Google Scholar]
- Kong, W.; McConalogue, K.; Khitin, L.M.; Hollenberg, M.D.; Payan, D.G.; Böhm, S.K.; Bunnett, N.W. Luminal trypsin may regulate enterocytes through proteinase-activated receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 8884–8889. [Google Scholar] [CrossRef]
- Scott, G. Proteinase-activated receptor-2 stimulates prostaglandin production in keratinocytes: Analysis of prostaglandin receptors on human melanocytes and effects of PGE2 and PGF2α on melanocyte dendricity. J. Invest. Dermatol. 2004, 122, 1214–1224. [Google Scholar] [CrossRef]
- Liu, M.; Dai, Y.; Li, Y.; Luo, Y.; Huang, F.; Gong, Z.; Meng, Q. Madecassoside isolated from Centella asiatica herbs facilitates burn wound healing in mice. Planta Med. 2008, 74, 809–815. [Google Scholar] [CrossRef]
- Cao, W.; Li, X.Q.; Zhang, X.N.; Hou, Y.; Zeng, A.G.; Xie, Y.H.; Wang, S.W. Madecassoside suppresses LPS-induced TNF-alpha production in cardiomyocytes through inhibition of ERK, p38, and NF-kappaB activity. Int. Immunopharmacol. 2010, 10, 723–729. [Google Scholar] [CrossRef]
- Haftek, M.; Mac-Mary, S.; le Bitoux, M.A.; Creidi, P.; Seité, S.; Rougier, A.; Humbert, P. Clinical, biometric and structural evaluation of the long-term effects of a topical treatment with ascorbic acid and madecassoside in photoaged human skin. Exp. Dermatol. 2008, 17, 946–952. [Google Scholar] [CrossRef]
- Maquart, F.X.; Bellon, G.; Gillery, P.; Wegrowski, Y.; Borel, J.P. Stimulation of collagen synthesis in fibroblast cultures by a triterpene extracted from Centella asiatica. Connect. Tissue Res. 1990, 24, 107–120. [Google Scholar] [CrossRef]
- Bian, D.; Liu, M.; Li, Y.; Xia, Y.; Gong, Z.; Dai, Y. Madecassoside, a triterpenoid saponin isolated from Centella asiatica herbs, protects endothelial cells against oxidative stress. J. Biochem. Mol. Toxicol. 2012, 26, 399–406. [Google Scholar] [CrossRef]
- An, I.S.; An, S.; Kang, S.M.; Choe, T.B.; Lee, S.N.; Jang, H.H.; Bae, S. Titrated extract of Centella asiatica provides a UVB protective effect by altering microRNA expression profiles in human dermal fibroblasts. Int. J. Mol. Med. 2012, 30, 1194–1202. [Google Scholar]
- Won, J.H.; Shin, J.S.; Park, H.J.; Jung, H.J.; Koh, D.J.; Jo, B.G.; Lee, J.Y.; Yun, K.; Lee, K.T. Anti-inflammatory effects of madecassic acid via the suppression of NF-kappaB pathway in LPS-induced RAW 264.7 macrophage cells. Planta Med. 2010, 76, 251–257. [Google Scholar] [CrossRef]
- Li, H.; Gong, X.; Zhang, L.; Zhang, Z.; Luo, F.; Zhou, Q.; Chen, J.; Wan, J. Madecassoside attenuates inflammatory response on collagen-induced arthritis in DBA/1 mice. Phytomedicine 2009, 16, 538–546. [Google Scholar] [CrossRef]
- Maldve, R.E.; Kim, Y.; Muga, S.J.; Fischer, S.M. Prostaglandin E2 regulation of cyclooxygenase expression in keratinocytes is mediated via cyclic nucleotide-linked prostaglandin receptors. J. Lipid Res. 2000, 41, 873–881. [Google Scholar]
- Enomoto, A.; Yoshihisa, Y.; Yamakoshi, T.; Ur Rehman, M.; Norisugi, O.; Hara, H.; Matsunaga, K.; Makino, T.; Nishihira, J.; Shimizu, T. UV-B radiation induces macrophage migration inhibitory factor-mediated melanogenesis through activation of protease-activated receptor-2 and stem cell factor in keratinocytes. Am. J. Pathol. 2011, 178, 679–687. [Google Scholar] [CrossRef]
- Springer, M.; Engelhart, K.; Biesalski, H.K. Effects of 3-isobutyl-1-methylxanthine and kojic acid on cocultures and skin equivalents composed of HaCaT cells and human melanocytes. Arch. Dermatol. Res. 2003, 95, 88–91. [Google Scholar]
- Ma, H.J.; Zhao, G.; Zi, S.X.; Li, D.G.; Liu, W.; Yang, Q.Q. Efficacy of quantifying melanosome transfer with flow cytometry in a human melanocyte-HaCaT keratinocyte co-culture system in vitro. Exp. Dermatol. 2010, 19, e282–e285. [Google Scholar]
- Jung, E.S.; Hwang, W.T.; Kim, Y.S.; Kim, S.B.; Kim, Y.S.; Lee, J.S.; Park, D.H. Depigmentation by platycodin D is mediated by cAMP-PKA/Rho-dependent signaling pathway. Exp. Dermatol. 2011, 20, 986–991. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the madecassoside are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jung, E.; Lee, J.-A.; Shin, S.; Roh, K.-B.; Kim, J.-H.; Park, D. Madecassoside Inhibits Melanin Synthesis by Blocking Ultraviolet-Induced Inflammation. Molecules 2013, 18, 15724-15736. https://doi.org/10.3390/molecules181215724
Jung E, Lee J-A, Shin S, Roh K-B, Kim J-H, Park D. Madecassoside Inhibits Melanin Synthesis by Blocking Ultraviolet-Induced Inflammation. Molecules. 2013; 18(12):15724-15736. https://doi.org/10.3390/molecules181215724
Chicago/Turabian StyleJung, Eunsun, Jung-A Lee, Seoungwoo Shin, Kyung-Baeg Roh, Jang-Hyun Kim, and Deokhoon Park. 2013. "Madecassoside Inhibits Melanin Synthesis by Blocking Ultraviolet-Induced Inflammation" Molecules 18, no. 12: 15724-15736. https://doi.org/10.3390/molecules181215724