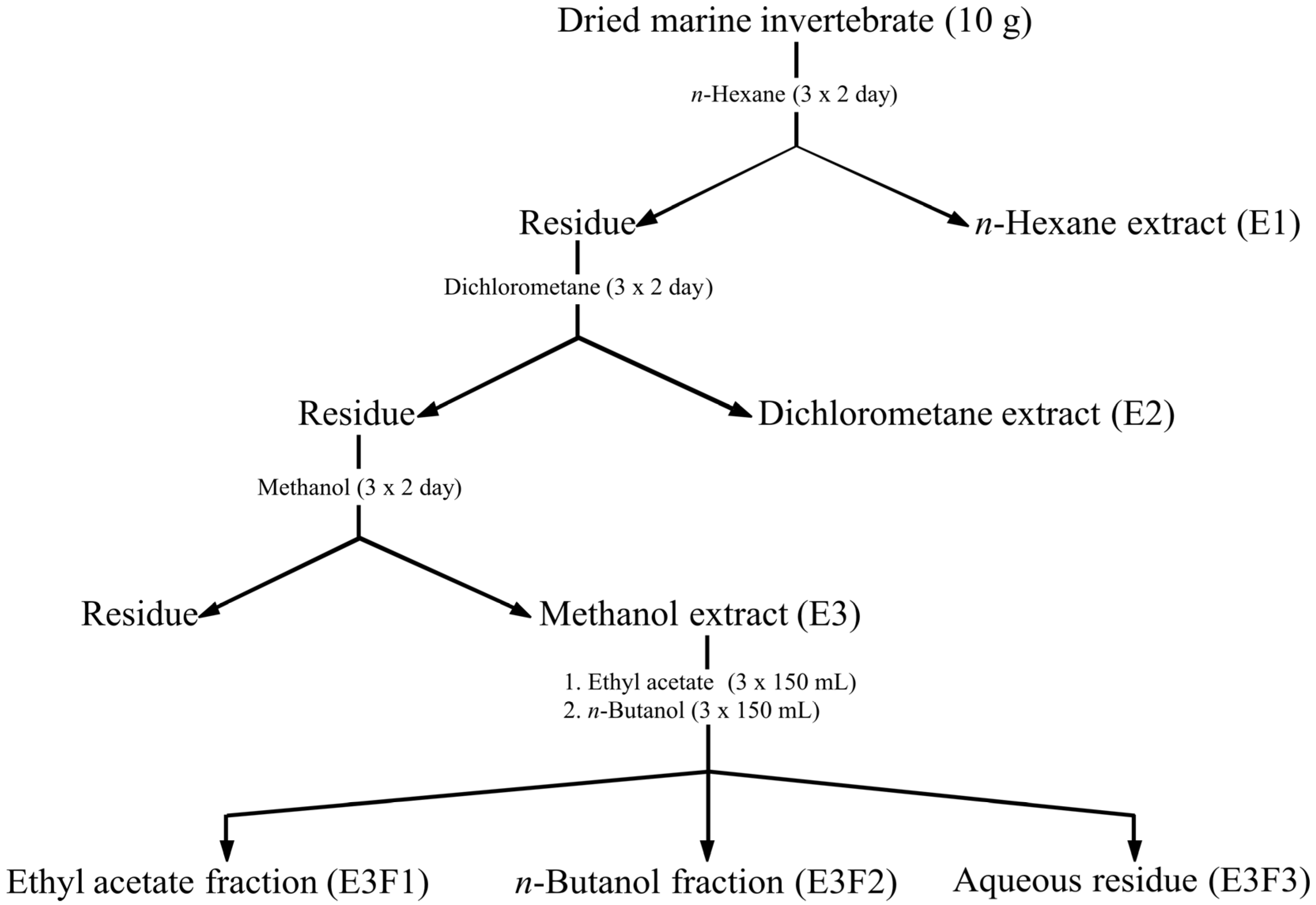
This paper describes the
in vitro antimicrobial, antiprotozoal and antiviral evaluation of organic extracts and fractions from 13 marine invertebrate species (nine sponges, one ascidian, two octocorals, and one bryozoans (
Table 1), and 27 seaweeds species [sixteen Rhodophyta (59.2%), seven Phaeophyceae (26%), and five Chlorophyta (14.8%) (
Table 2). A total of 95 extracts and fractions (65 from marine invertebrates and 30 from seaweeds) were assayed. The results showed that 53 samples (56%) exhibited some anti-infective activity against
Staphylococcus aureus,
Enterococcus faecalis,
Pseudomonas aeruginosa,
Escherichia coli and
Candida albicans (antimicrobial),
Leishmania braziliensis and
Trypanosoma cruzi (anti-protozoal), as well as against HSV-1 replication (antiviral).
2.1. Marine Invertebrates
Thirteen species of marine invertebrates were assayed against bacteria and fungus. Of these, seven [
Dragmacidon reticulatum,
Dragmaxia anomala,
Haliclona (
Halichoclona) sp.,
Leptogorgia punicea,
Petromica citrina,
Tedania ignis, and
Trachycladus sp.], showed some activity (
Table 3).
The most interesting antimicrobial results were obtained with the sponge
D. reticulatum thatshowed significant growth inhibition (13–16 mm) against
S. aureus and
C. albicans. A similar result was proved in another screening carried out with marine organisms from the southeastern Brazilian coast where the sponge
D. reticulatum showed a weak antimicrobial activity against
S. aureus and
C. albicans [
24]
. As far as we are aware, this is the first report of this biological activity for
D. reticulatum,
D. anomala, and
Trachycladus sp.
Furthermore, a weak antimicrobial activity against S. aureus, E. faecalis, and E. coli was also detected in the present study for Haliclona (Halichoclona) sp. and Petromica citrina (9–12 mm inhibition zone), and T. ignis (6–8 mm inhibition zone). In the same way, we verified a weak antimicrobial activity against S. aureus and C. albicans of the n-hexane extract from the octocoral Leptogorgia punicea.
Table 3.
Antibacterial and antifungal screening of marine invertebrates by disc diffusion method.
Table 3.
Antibacterial and antifungal screening of marine invertebrates by disc diffusion method.
| Species | Extracts | Bacterial and fungal strains |
|---|
| S. aureus | E. faecalis | E. coli | P. aeruginosa | C. albicans |
|---|
| Dragmaxia anomala | E1 | + | + | − | − | − |
| Dragmacidon reticulatum | E3F1 | +++ | − | − | − | +++ |
| Haliclona (Halichoclona) sp. | E3F2 | ++ | ++ | + | - | + |
| Leptogorgia punicea | E1 | ++ | − | − | − | ++ |
| Petromica citrina | E3F2 | ++ | ++ | - | - | ++ |
| Tedania ignis | E2 | − | + | − | − | − |
| Trachycladus sp. | E3F2 | ++ | ++ | + | − | + |
| E3F3 | − | + | − | − | − |
As far as we aware, this is the first report for antimicrobial activity for this gorgonian species. On the other hand, the extracts from B. neritina, C. riseii, C. celata, D. granulatum, G. sepia and P. janeirensis did not show antimicrobial activity against the assayed microorganism strains.
Recently it was reported that aqueous extract of
P. citrina (collected in Rio de Janeiro State, southeast of Brazil) showed a large spectrum of activity against clinical strains and resistant-bacteria including
S. aureus,
S. epidermidis,
E. coli,
E. faecalis,
M. fortuitum and
N. gonorrhoeae. All these activities were related to the presence of halistanol trisulphate A in this marine sponge [
29,
30]. Moreover, the antifungal activity for this substance, isolated from
Petromica ciocalyptoides was also reported [
31].
Another study led by Monks
et al. [
19] concerning the antimicrobial activity against
E. coli,
S. aureus,
S. epidermis,
B. subtilis, and
M. luteus strains of southern Brazilian sponges, including
Guitarra sp.,
T. ignis,
Haliclona aff.
tubifera, demonstrated that
H. aff.
tubifera showed moderate activity against
E. coli and weak activity against
S. aureus,
S. epidermis,and
M. luteus. Our results are in agreement with those obtained by these authors, although we used different libraries of microbial strains and extracts.
Concerning antiprotozoal activity, few studies reporting antileishmanial and tripanocidal activities have been described for marine invertebrates. In this work, 13 marine invertebrate species were evaluated against
L. brasiliensis and
T. cruzi (
Table 4).
Table 4.
Antiprotozoal activity expressed as growth inhibition (%) of extracts and fractions obtained from marine invertebrates.
Table 4.
Antiprotozoal activity expressed as growth inhibition (%) of extracts and fractions obtained from marine invertebrates.
| Species | Samples | Leishmania braziliensis (promastigotes) | Trypanosoma cruzi (epimastigotes) |
|---|
| Bugula neritina | E1 | 66 | − |
| E3F2 | 47 | − |
| E3F3 | 30.7 | − |
| Carijoa riisei | E1 | 35.9 | 43.4 |
| E2 | − | 29 |
| E3F1 | − | 26.1 |
| E3F2 | 14.6 | 2.6 |
| E3F3 | − | 5.5 |
| Didemnun granulatum | E1 | − | 21.5 |
| E3F1 | 15.7 | 13.2 |
| E3F2 | 17.9 | − |
| Dragmacidon reticulatum | E1 | 24.1 | 11.4 |
| E2 | − | 20 |
| E3F1 | − | 21.2 |
| E3F2 | 19.8 | − |
| E3F3 | 13.8 | 15.3 |
| Dragmaxia anomala | E1 | 97.2 | 71.7 |
| Guitarra sepia | E3F2 | 12.5 | − |
| E3F3 | 14.9 | − |
| Haliclona (Halichoclona) sp. | E1 | − | 14.6 |
| E3F1 | − | 28.3 |
| E3F2 | 43.6 | 33 |
| E3F3 | 16.9 | − |
| Leptogorgia punicea | E1 | − | 19.2 |
| E2 | − | 38.8 |
| E3F2 | − | 11.1 |
| E3F3 | 15 | − |
| Tedania ignis | E2 | 16.1 | − |
| E3F1 | 12.2 | − |
| E3F2 | 18.4 | − |
| E3F3 | 19.1 | − |
Out of these 13 species tested, Bugula neritina (E1 extract), Carijoa riisei (E1 extract), Dragmaxia anomala (extract E1), and Haliclona (Halichoclona) sp. (fraction E3F2) showed the best results, particularly against extracellular promastigote forms of L. braziliensis (66, 35.9, 97.2 and 43.6% grown inhibition, respectively). However, only two species, C. riisei and D. anomala showed some tripanocidal effects (43.4 and 71.7% growth inhibition, respectively).
Additionally, these extracts and fractions were assayed on
L. brasiliensis amastigotes in bone marrow macrophages from mice, and only the sponge
Haliclona (
Halichoclona) sp. and the octocoral
C. riisei were active (
Table 5). Based on these preliminary results, the E1 extract from
C. riisei was fractionated by chromatographic techniques leading to the isolation of an active pregnane steroid [
32]. Finally, the extracts from
C. celata,
P. citrina,
P. janeirensis and
Trachycladus sp. were not active against
L. braziliensis or
T. cruzi.
Table 5.
Effects of marine invertebrates extracts and fractions on Leishmania brasiliensis amastigotes in bone marrow macrophages from mice, and cytotoxicity on J774.G8 macrophage cell line.
Table 5.
Effects of marine invertebrates extracts and fractions on Leishmania brasiliensis amastigotes in bone marrow macrophages from mice, and cytotoxicity on J774.G8 macrophage cell line.
| Species | Samples | CC50 ± SD (µg/mL) | IC50 ± SD (µg/mL) | Selective index (CC50/IC50) |
|---|
| Bugula neritina | E1 | ND | >50 | ND |
| Carijoa riisei | E1 | 48.6 ± 4.8 | 43.3 ± 8.5 | 1.1 |
| Dragmaxia anomala | E1 | 54.3 ± 1.9 | >15 | <3.6 |
| Haliclona (Halichoclona) sp. | E3F2 | 279.7 ± 21.2 | 43.9 ± 3.4 | 6.8 |
In this work, the antiviral activity against Herpes Simplex Virus type 1 (HSV-1, KOS strain) was also evaluated. Before the evaluation of the antiviral activity, the cytotoxic effects of the selected samples were investigated on VERO cells by MTT assay, and for each tested sample, a CC50 value was calculated. Of the 95 extracts and fractions tested, only the E3F2 fractions from the sponges Haliclona (Halichoclona) sp. and P. citrina showed antiviral activity (SI = 11.92 and, SI > 5, respectively).
In 2006, Silva and co-workers [
20] performed an
in vitro study on the antiherpes, anti-adenovirus and anti-rotavirus activities of marine sponges collected from the Brazilian coast, including
Haliclona sp.,
Polymastia janeirensis and
T. ignis. Of these, only the organic extract (methanol/toluene, 3:1 v/v) from
P. janeirensis showed antiherpetic activity [
20].
2.2. Marine Seaweeds
Only five out of 27 species from seaweeds assayed (Rhodophyta:
Digenea simplex,
Laurencia dendroidea,
Ochtodes secundiramea,
Osmundaria obtusiloba, andPhaeophyta:
Dictyota sp.) showed weak growth inhibition zone (6 to 8 mm, (
Table 6). Otherwise,
A. specifera,
A. saldanhae,
A. stellata,
B. occidentalis,
B. seaforthii,
B. triquetrum,
C. cervicornis,
C. sertularioides,
C. cupressoidesa,
C. cupressoidesb,
C. seminervis,
D. delicatula,
D. jolyana,
G. caudata,
G. cervicornis,
G. cuneifólia,
H. cenomyce,
H. musciformis,
P. gymnospora,
Padina sp
.,
P. flagellifera,
P. papillosa and
Sargassum sp. did not show any antimicrobial activity.
Antibacterial activity may vary according to the species division [
33]. In this study species from the phylum Rhodophyta exhibited better results than species from Chlorophyta and Phaeophyceae. In this context, our results are in agreement with the findings of Padmakumar and Ayyakkannu [
34]. Members of the family Rhodophyceae are prolific producers of acetogenins as well as mono-, sesqui-, di- and triterpenes, many of them halogenated [
35]. Many articles have reported antimicrobial activity of halogenated sesquiterpenes and acetogenins derived from
Laurencia species, especially for (−)-elatol, obtusol, (+)-obtusane, cartilagineol, and triquinane derivatives [
36,
37]. Studies performed with
Osmundaria species are scarce, but some compounds so far described have potential antimicrobial activity, particularly the halogenated phenol derivatives, such as lanosol and sulfated oligobromophenols [
38,
39].
Table 6.
Antibacterial and antifungal screening of marine seaweeds by disc diffusion method.
Table 6.
Antibacterial and antifungal screening of marine seaweeds by disc diffusion method.
| Species | Extracts | Bacterial and fungal strains |
|---|
| S. aureus | E. faecalis | E. coli | P. aeruginosa | C. albicans |
|---|
| Dictyota sp. | DS | + | + | − | + | − |
| Digenea simplex | DS | + | + | − | + | − |
| Laurencia dendroidea | FS | + | + | − | − | − |
| Ochtodes secundiramea | FS | + | − | − | − | − |
| Osmundaria obtusiloba | DS | − | − | − | ++ | − |
Regarding the antiprotozoal activity, of the 27 species assayed, six showed interesting activity against
L. braziliensis and
T. cruzi. Extracts from
Anadyomene saldanhae (FS extract),
Caulerpa cupressoides(a) (FS extract),
Canistrocarpus cervicornis (FS extract),
Dictyota sp. (FS extract),
Ochtodes secundiramea (FS extract), and
Padina sp. (FS extract) showed promising results against
L. braziliensis (87.9, 51.7, 85.9, 93.3, 99.7, and 80.9% growth inhibition, respectively). Only
Dictyota sp. was effective against
T. cruzi (60.4% growth inhibition) (
Table 7). Otherwise,
B. triquetrum,
C. sertularioides,
C. cupressoidesb,
D. delicatula,
G. caudata,
H. cenomyce,
H. musciformis,
P. papillosa and
Sargassum sp., none antiprotozoal activity was detected.
As far as we are aware, there are no reports in the literature on the antiprotozoal activity for four of these seaweeds species (
A. saldanhae,
C. cupressoidesa,
Padina sp., and
O. secundiramea). Concerning the activity of FS extract from the red seaweed
L. dendroidea (= formerly
Laurencia obtusa), only a weak (14.6%) antileishmanial activity against the promastigote forms of
L. braziliensis was observed (
Table 7). Furthermore, two species of seaweeds,
A. saldanhae (SI = 12.3) and
Padina sp. (SI = 7.5), were effective against
L. brasiliensis amastigotes. Additionally,
C. cervicornis,
C. cupressoidesa,
Dictyota sp., and
O. secundiramea were strongly cytotoxic for bone marrow macrophages (
Table 8).
Table 7.
Antiprotozoal activity expressed as growth inhibition (%) of extracts and fractions obtained from marine seaweeds.
Table 7.
Antiprotozoal activity expressed as growth inhibition (%) of extracts and fractions obtained from marine seaweeds.
| Species | Extracts | Leishmania braziliensis (promastigotes) | Trypanosoma cruzi (epimastigotes) |
|---|
| Anadyomene saldanhae | FS | 87.9 | − |
| Botryocladia occidentalis | DS | 20.7 | − |
| Bryothamnion seaforthii | DS | 33.5 | − |
| Canistrocarpus cervicornis | FS | 85.8 | − |
| Caulerpa cupressoides | FS | 51.7 | − |
| Dictyota sp. | DS | 93.3 | 60.4 |
| Digenea simplex | DS | 26 | − |
| Gracilaria caudata | DS | 9.3 | − |
| Grateloupia cuneifolia | FS | 35.2 | 15.9 |
| DS | 37 | 23.98 |
| Laurencia dendroidea | FS | 14.6 | − |
| Ochtodes secundiramea | FS | 99.7 | − |
| Padina sp. | FS | 80.9 | − |
| Palisada flagellifera | DS | 21 | − |
Table 8.
Effects of marine seaweeds extracts and fractions on Leishmania brasiliensis amastigotes in bone marrow macrophages from mice, and cytotoxicity on J774.G8 macrophage cell line.
Table 8.
Effects of marine seaweeds extracts and fractions on Leishmania brasiliensis amastigotes in bone marrow macrophages from mice, and cytotoxicity on J774.G8 macrophage cell line.
| Species | Samples | CC50 ± SD (µg/mL) | IC50 ± SD (µg/mL) | Selective index (CC50/IC50) |
|---|
| Anadyomene saldanhae | FS | 294.2 ± 28.2 | 23.9±2.3 | 12.3 |
| Canistrocarpus cervicornis | FS | >50 | ND | ND |
| Caulerpa cupressoides(a) | FS | >50 | ND | ND |
| Dictyota sp. | DS | >50 | ND | ND |
| Ochtodes secundiramea | FS | >50 | ND | ND |
| Padina sp. | FS | 300.4 ± 28.5 | 40.2 ± 4.3 | 7.5 |
Previous studies performed by Veiga-Santos
et al. [
5] and Machado
et al. [
37] showed that lipophilic extracts from
L. dendroidea collected from the southeastern coast of Brazil strongly inhibited the growth of
T. cruzi and
L. amazonensis. These results are not completely in agreement with our findings for
L. dendroidea and this discrepancy may be due to the different geographic regions where this species was collected, as well as the seawater conditions.
Another study led by Santos and colleagues [
4] found that lipophilic extracts from the brown seaweed
C. cervicornis collected from the northeastern coast of Brazil also strongly inhibited the growth of
L. amazonensis. From this species, a 4-acetoxydolastane diterpene was isolated, which demonstrated dose-dependent activity during 72 h of treatment, exhibiting IC
50 values of 2.0, 12.0 and 4.0 μg/mL for promastigotes, axenic amastigotes and intracellular amastigotes of
L. amazonensis, respectively.
Concerning antiviral activity, none of the species tested displayed any anti-HSV-1 activity. Although Soares and colleagues [
25] reported the anti-HSV-1 activity for the red alga
L. dendroidea collected from the coast of Rio de Janeiro, in our work this specie showed high cytotoxicity against VERO cells and none antiviral activity was detected.
To summarize, the present work reports the antimicrobial, antiprotozoal and antiviral evaluation of organic extracts from nine sponges, two octocorals, one ascidian, one bryozoan, and 27 seaweeds species, collected along the Brazilian coastline. Of a total of 95 extracts and fractions, 53 (56%) showed some anti-infective activity against S. aureus, E. faecalis, P. aeruginosa, E. coli, C. albicans, L. braziliensis, T. cruzi, and HSV-1.
Clearly, the marine invertebrates and seaweeds from the Brazilian coast could play an important part in the future control of the global infectious-disease burden. Although substantial progress has been made in identifying new biotechnological potential from these organisms, further chemical analysis and biological studies are required for investigating the mechanism of action, the chemical content as well as the potential use of these marine organisms extracts in the prevention of pathologies.


{kind=link}