Isoferulic Acid, a New Anti-Glycation Agent, Inhibits Fructose- and Glucose-Mediated Protein Glycation in Vitro



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
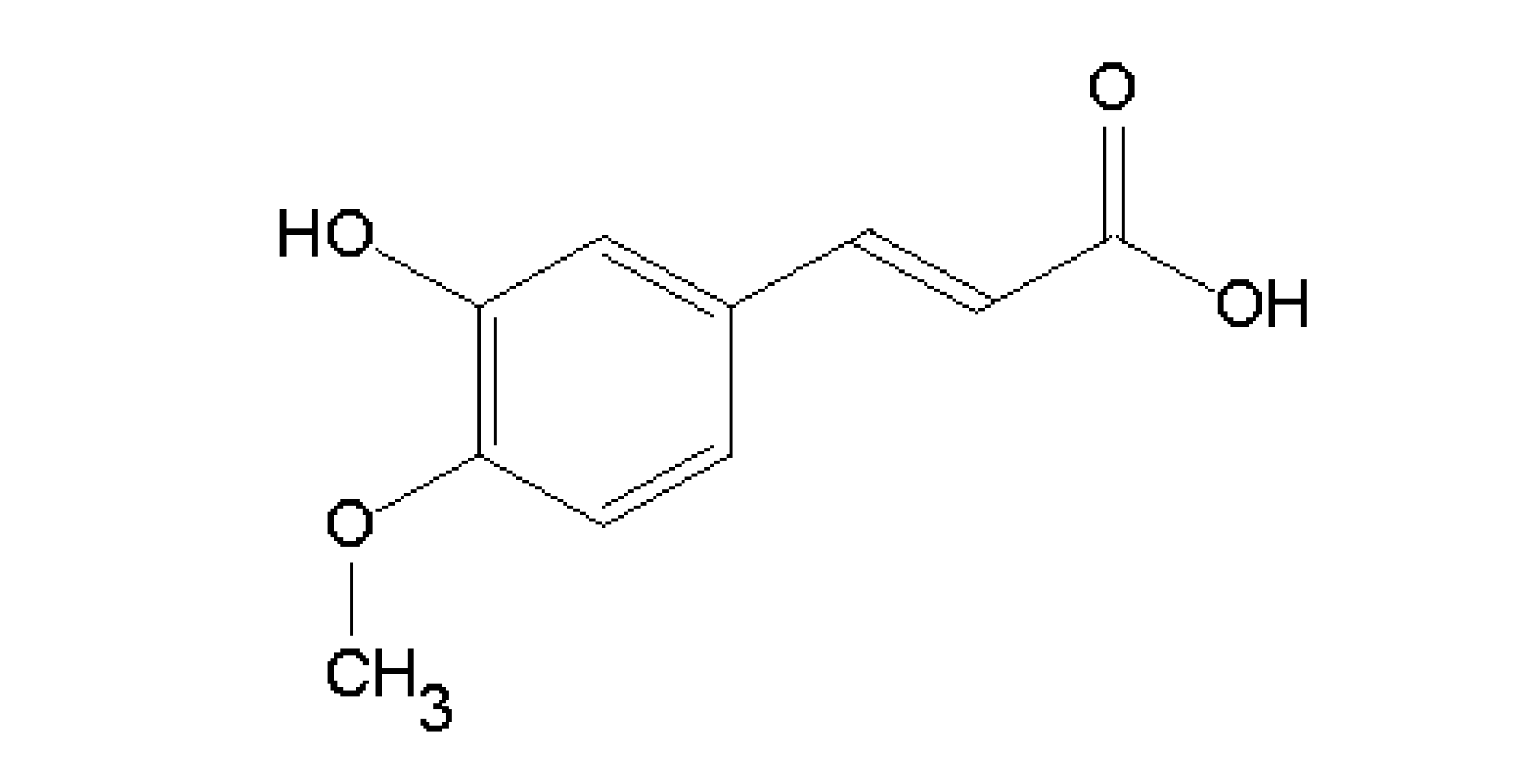
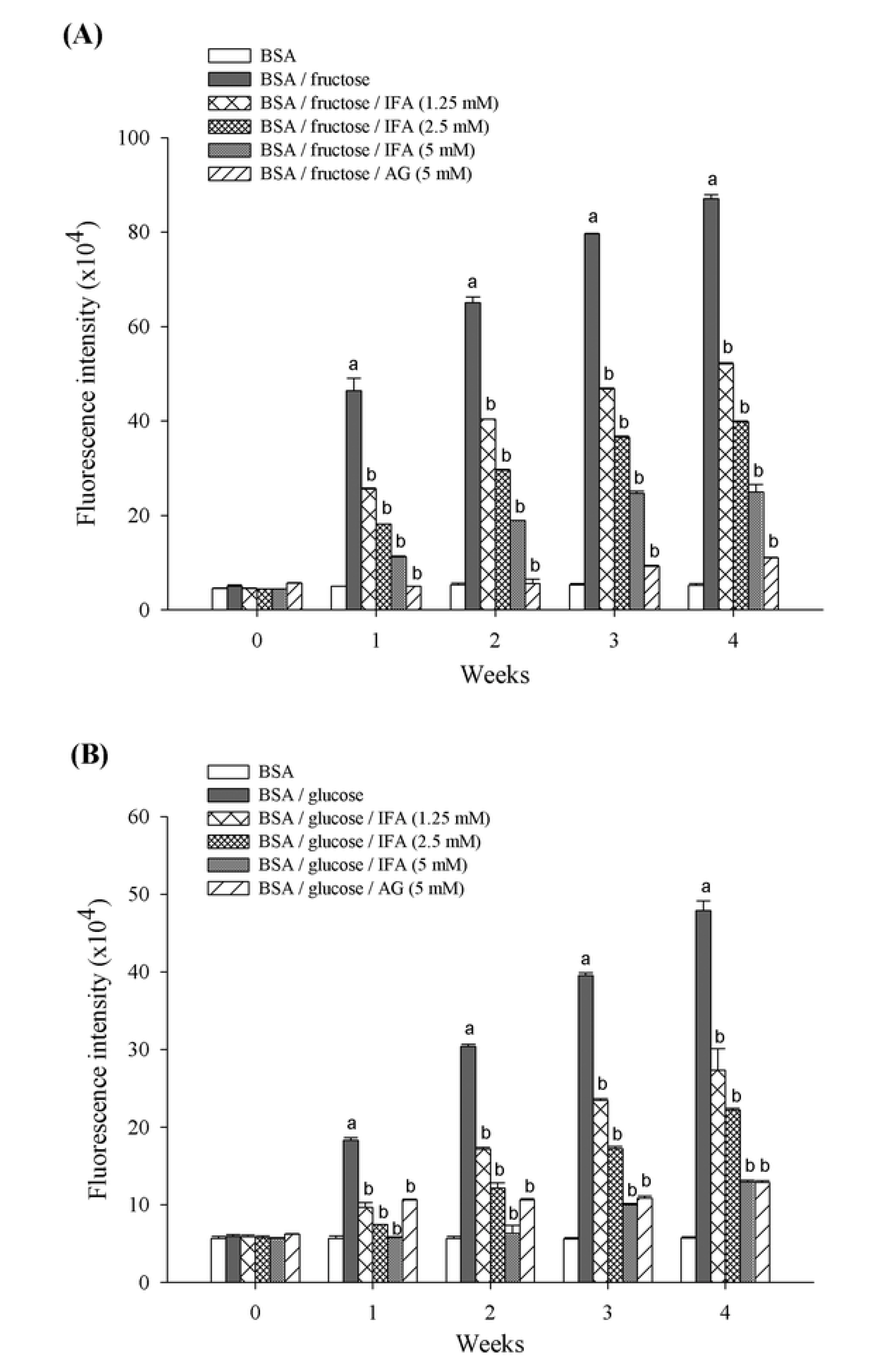
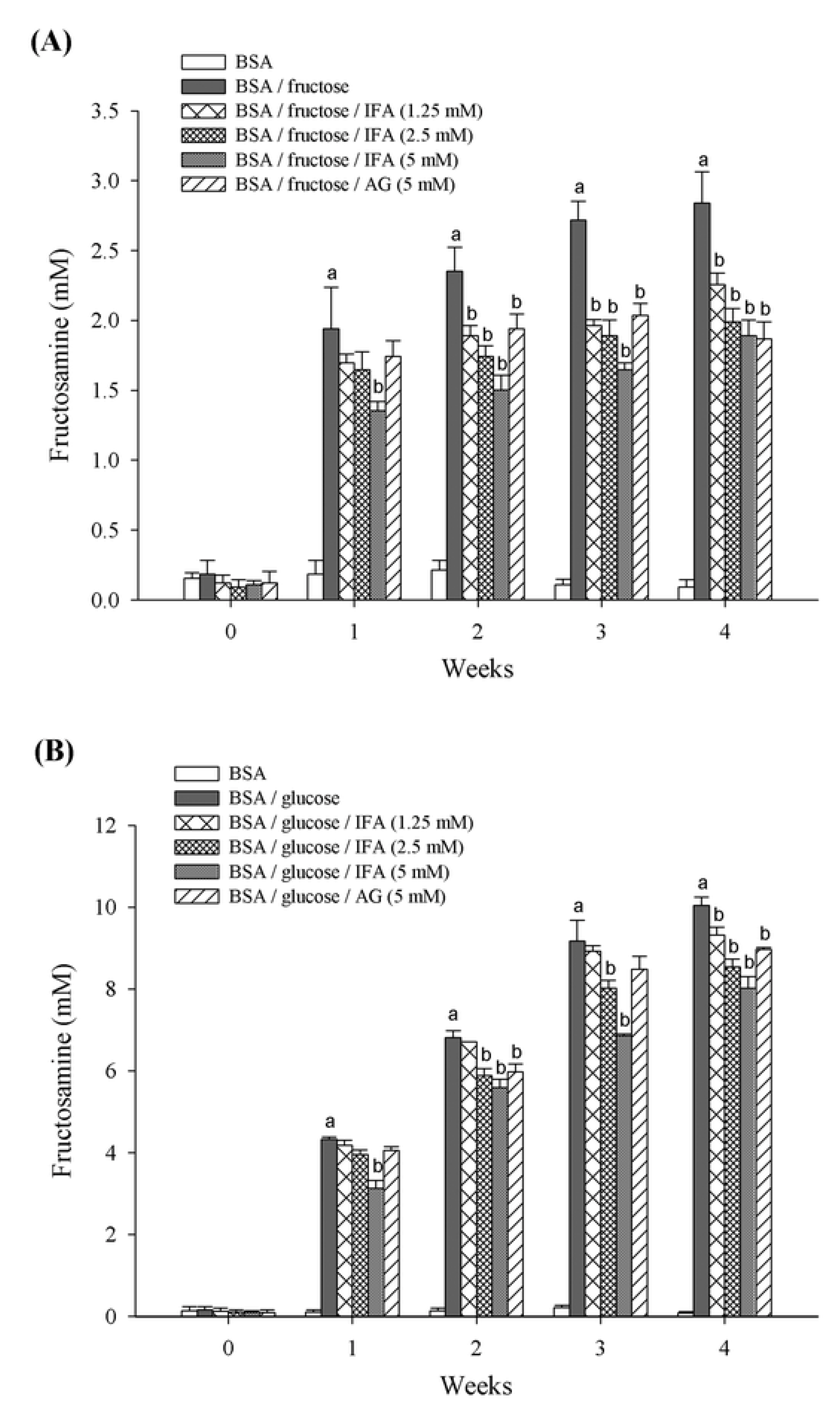
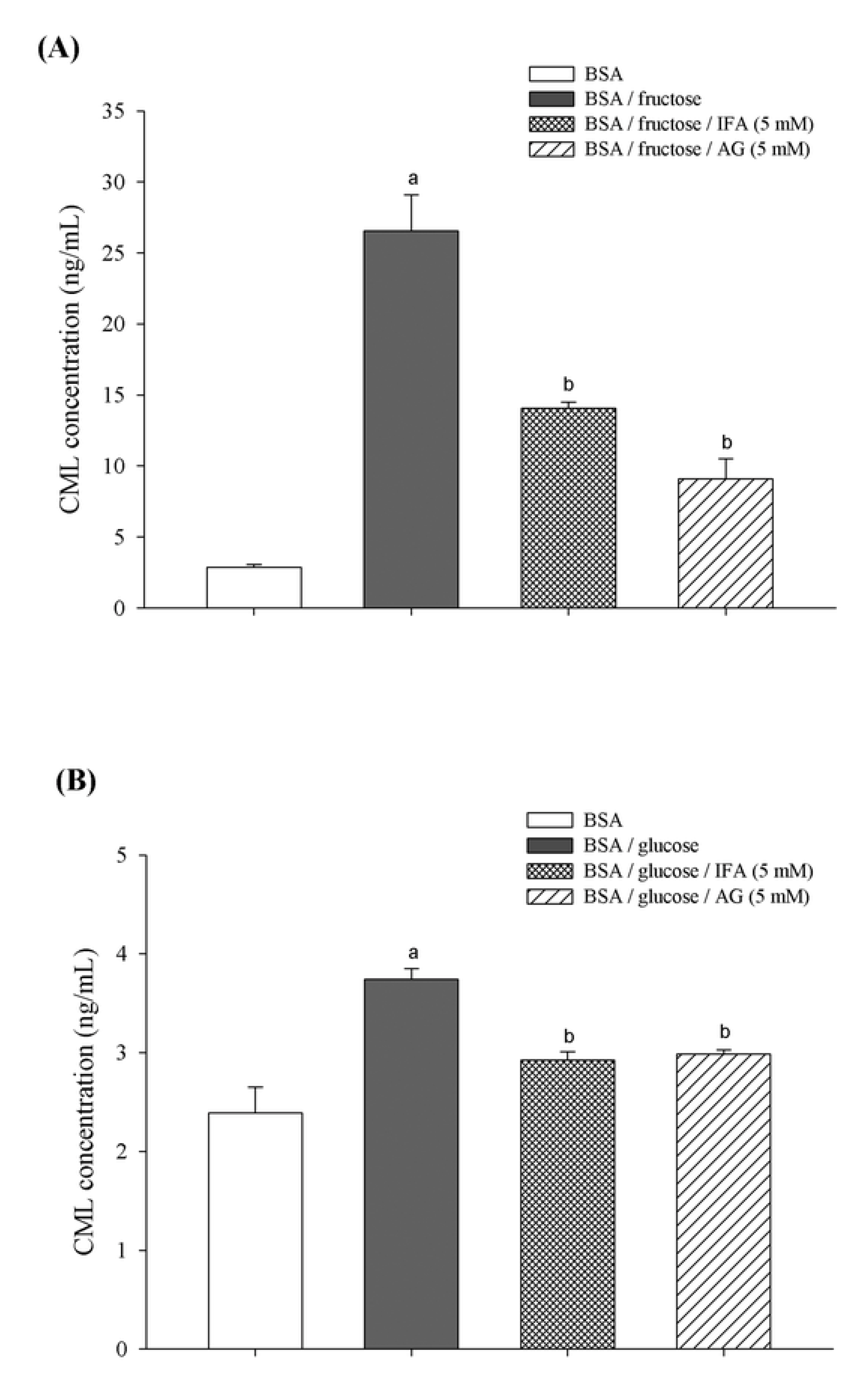
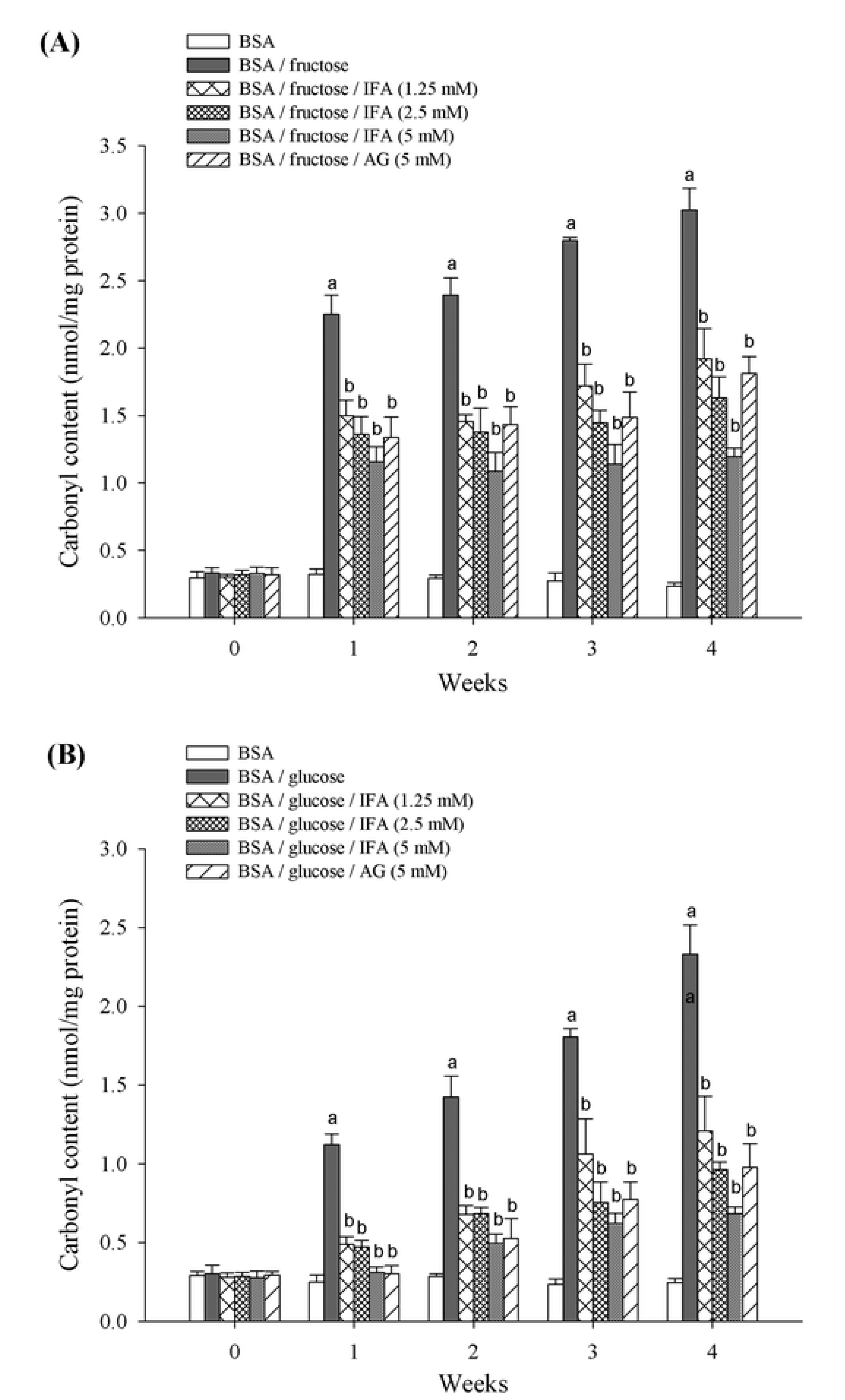
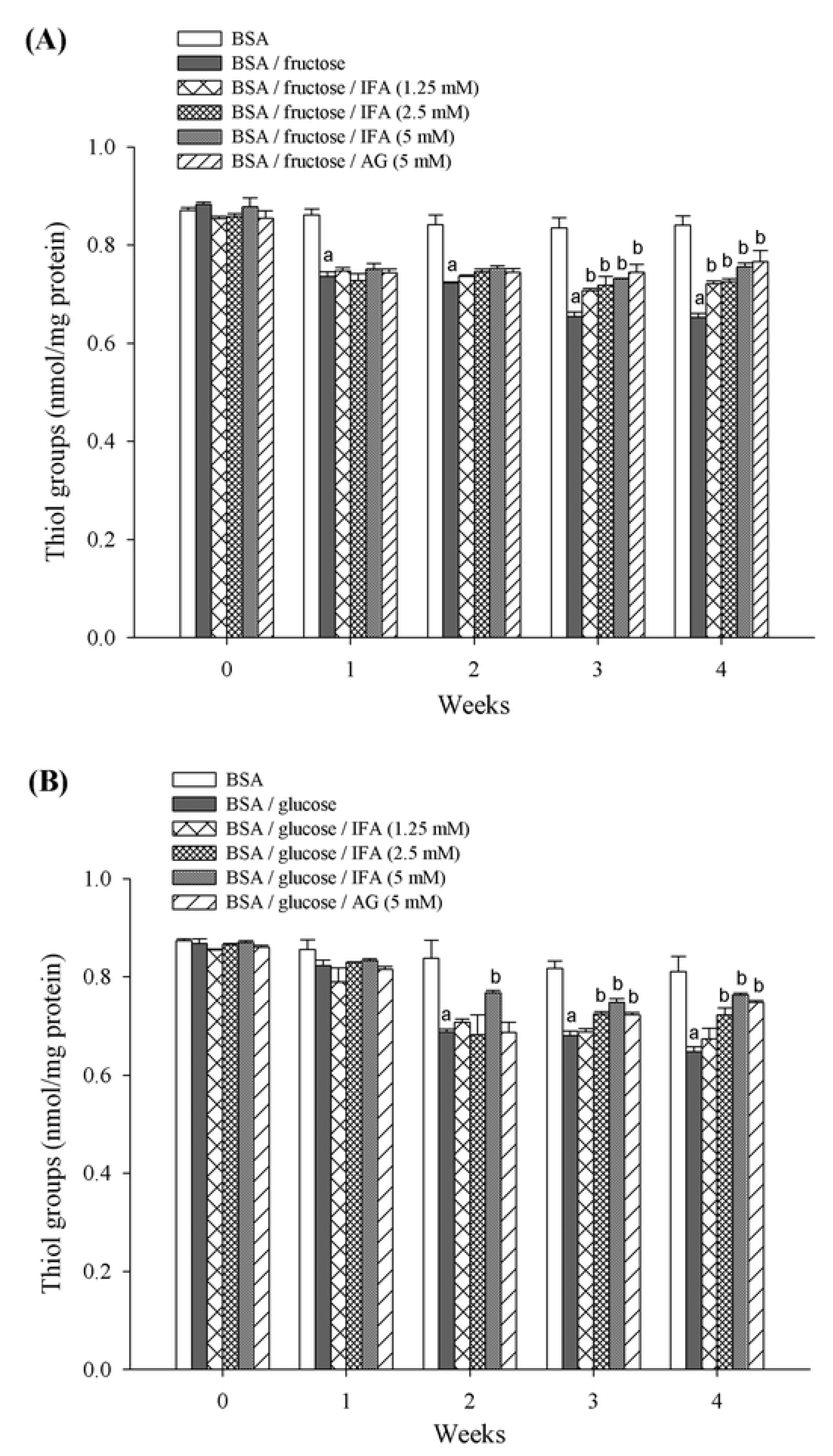
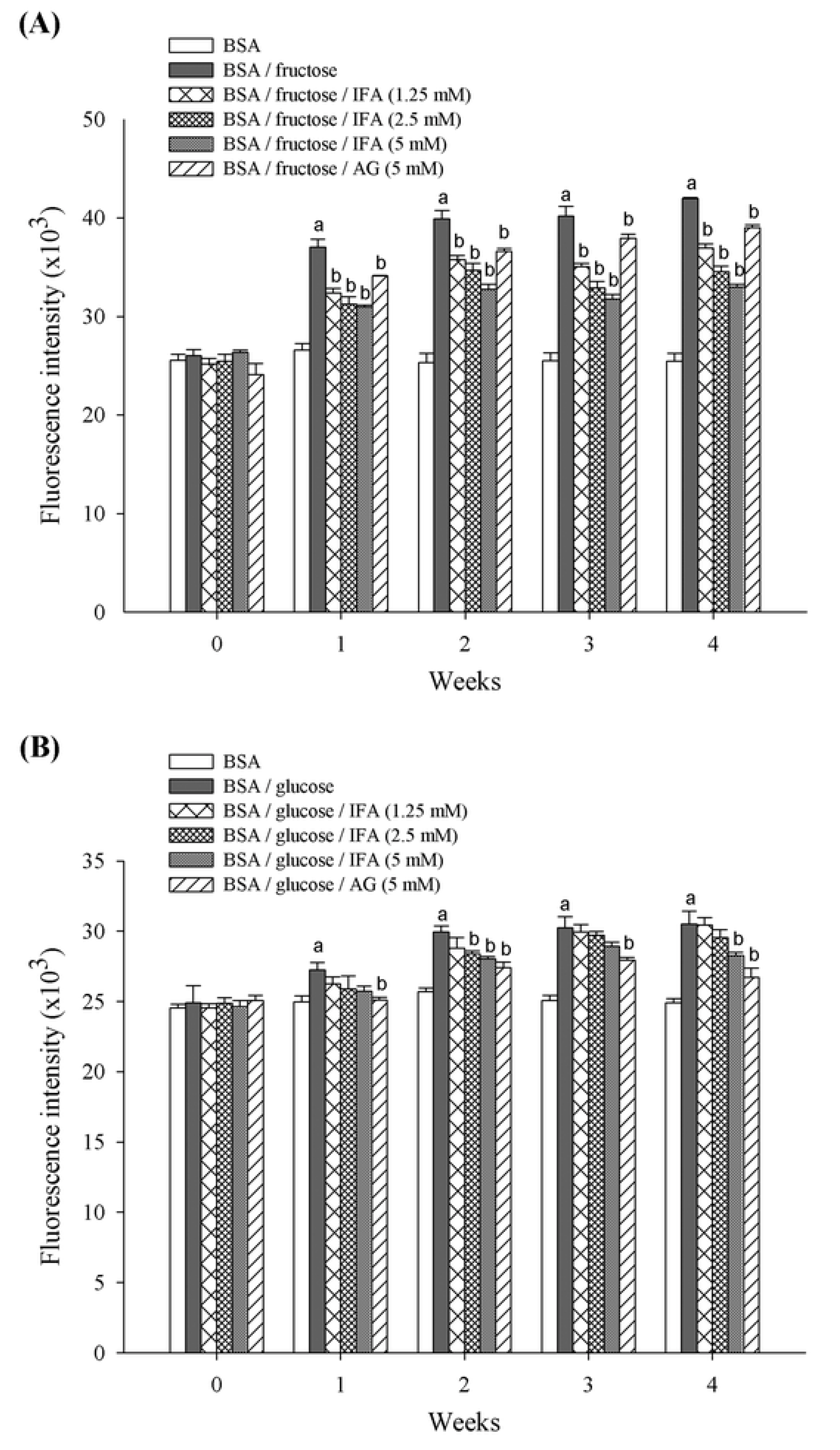
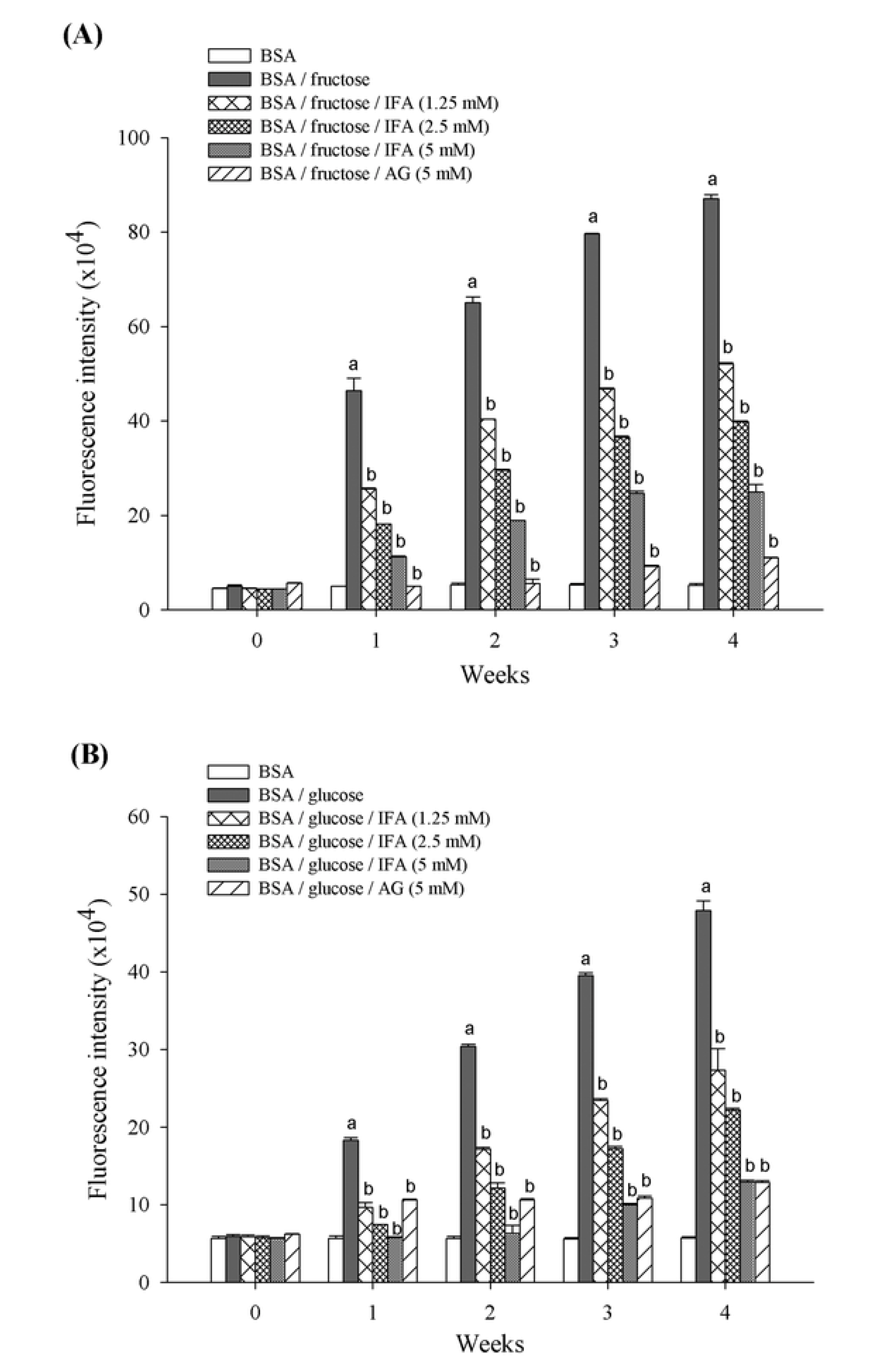
2. Results and Discussion
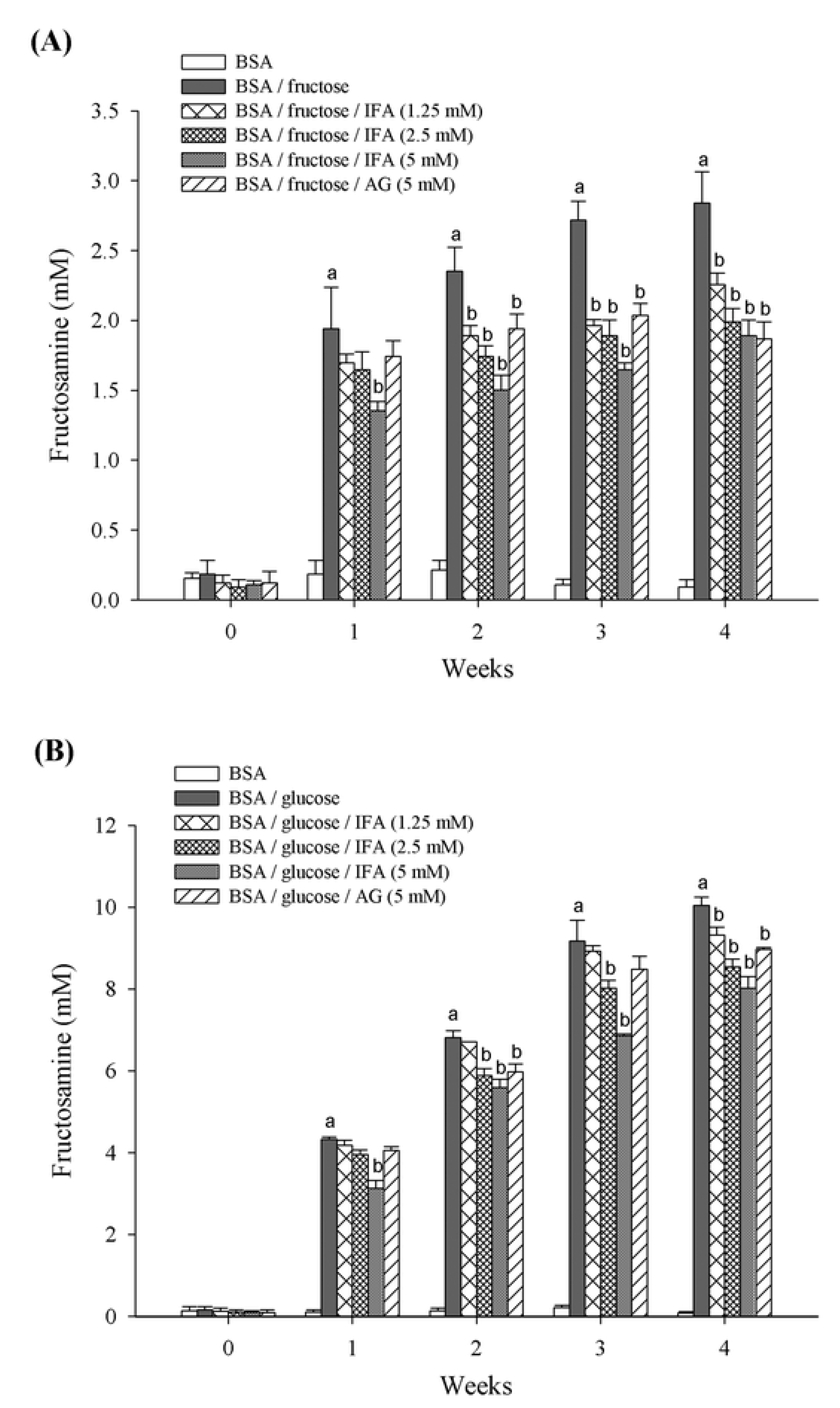
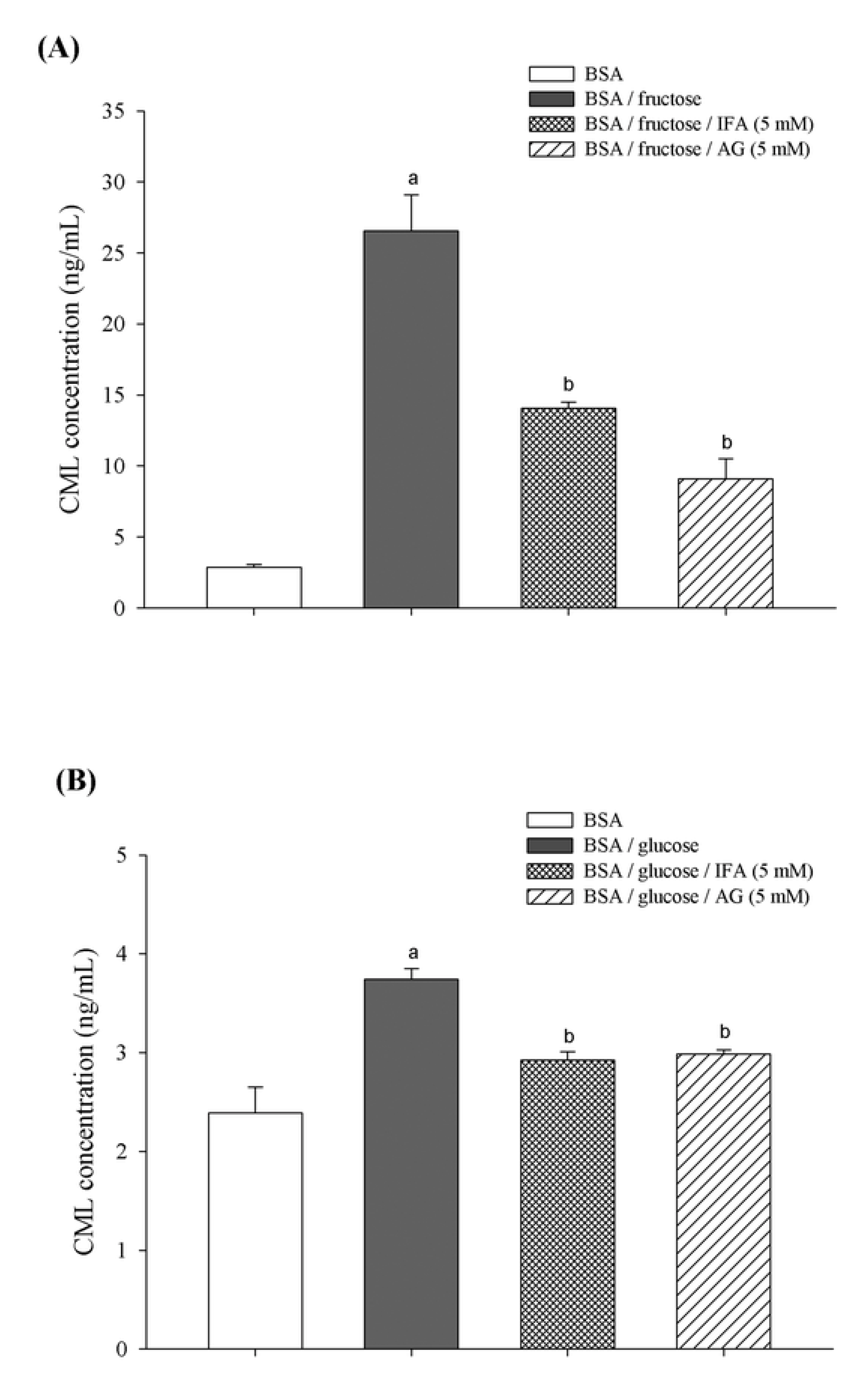


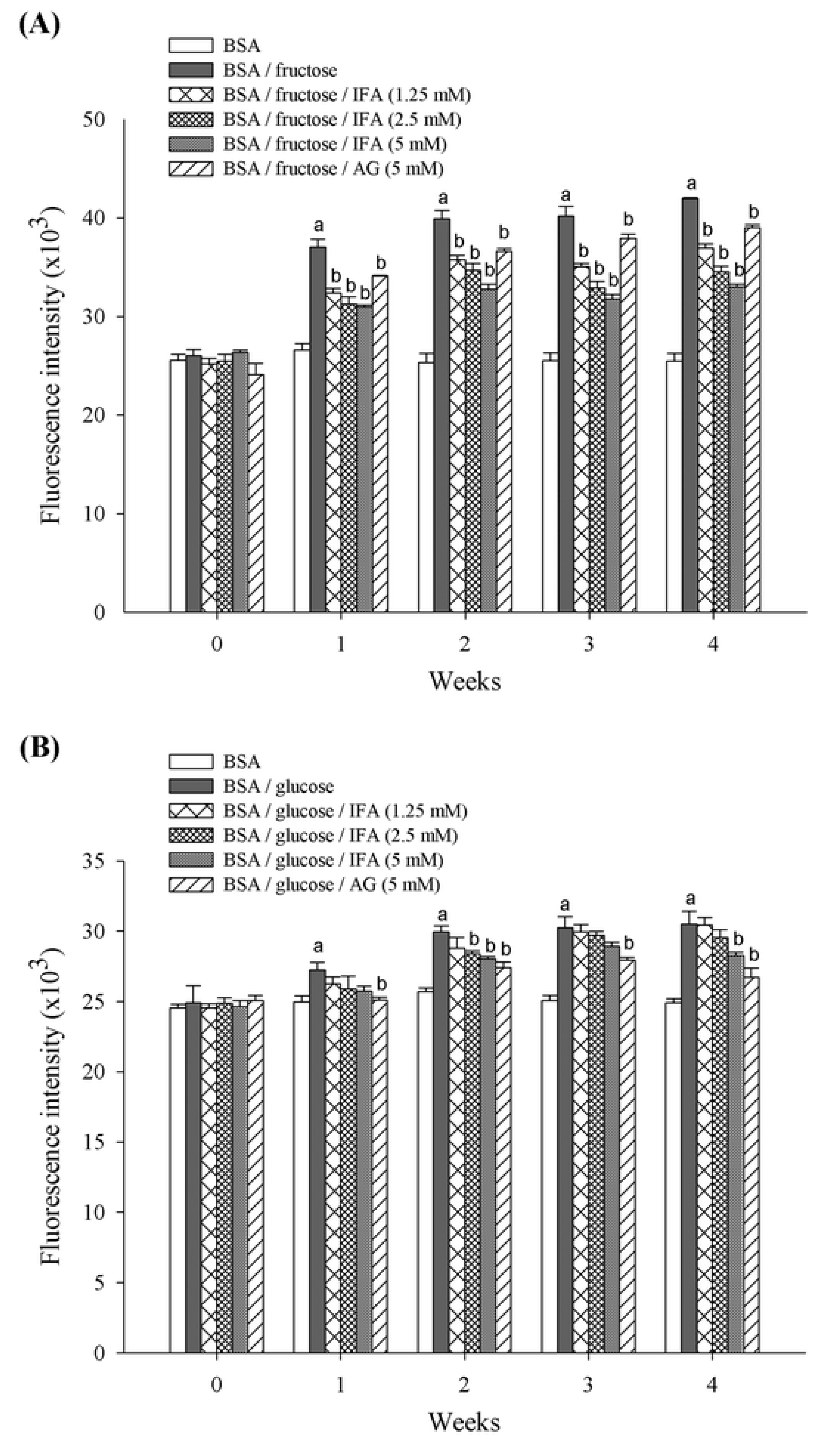
3. Experimental
3.1. Chemicals and Reagents
3.2. In Vitro Glycation of Bovine Serum Albumin (BSA)
3.3. Determination of AGEs Formation
3.4. Fructosamine Measurement
3.5. Determination of Nε-(carboxymethyl) Lysine (CML)
3.6. Determination of Protein Carbonyl Content
3.7. Thiol Group Estimation
3.8. Thioflavin T Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ahmed, N. Advanced glycation endproducts — Role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar]
- Negre-Salvayre, A.; Salvayre, R.; Augé, N.; Pamplona, R.; Portero-Otín, M. Hyperglycemia and glycation in diabetic complications. Antioxid. Redox. Signal. 2009, 11, 3071–3109. [Google Scholar]
- Vinson, J.A.; Howard, T.B., III. Inhibition of protein glycation and advanced glycation end products by ascorbic acid and other vitamins and nutrients. J. Nutr. Biochem. 1996, 7, 659–663. [Google Scholar]
- Ardestani, A.; Yazdanparast, R. Inhibitory effects of ethyl acetate extract of teucrium polium on in vitro protein glycoxidation. Food Chem. Toxicol. 2007, 45, 2402–2411. [Google Scholar]
- Bär, K.J.; Franke, S.; Wenda, B.; Müller, S.; Kientsch-Engel, R.; Stein, G.; Sauer, H. Pentosidine and n(epsilon)-(carboxymethyl)-lysine in alzheimer’s disease and vascular dementia. Neurobiol. Aging. 2003, 24, 333–338. [Google Scholar]
- Dworacka, M.; Winiarska, H.; Szymańska, M.; Szczawińska, K.; Wierusz-Wysocka, B. Serum n-epsilon-(carboxymethyl)lysine is elevated in nondiabetic coronary heart disease patients. J. Basic Clin. Physiol. Pharmacol. 2002, 13, 201–213. [Google Scholar]
- Ulrich, P.; Cerami, A. Protein glycation, diabetes, and aging. Recent Prog. Horm. Res. 2001, 56, 1–21. [Google Scholar]
- Khazaei, M.R.; Bakhti, M.; Habibi-Rezaei, M. Nicotine reduces the cytotoxic effect of glycated proteins on microglial cells. Neurochem. Res. 2010, 35, 548–558. [Google Scholar]
- Kousar, S.; Sheikh, M.A.; Asghar, M.; Sarwar, M. Effect of aminoguanidine on advanced glycation end products (ages) using normal and diabetic plasma. J. Chem. Soc. Pak. 2009, 31, 109–114. [Google Scholar]
- Yamagishi, S.; Nakamura, K.; Matsui, T.; Ueda, S.; Fukami, K.; Okuda, S. Agents that block advanced glycation end product (age)-rage (receptor for ages)-oxidative stress system: A novel therapeutic strategy for diabetic vascular complications. Expert Opin. Investig. Drugs 2008, 17, 983–996. [Google Scholar]
- Giardino, I.; Fard, A.K.; Hatchell, D.L.; Brownlee, M. Aminoguanidine inhibits reactive oxygen species formation, Lipid peroxidation, And oxidant-induced apoptosis. Diabetes 1998, 47, 1114–1120. [Google Scholar]
- Ihm, S.-H.; Yoo, H.J.; Park, S.W.; Ihm, J. Effect of aminoguanidine on lipid peroxidation in streptozotocin-induced diabetic rats. Metabolism 1999, 48, 1141–1145. [Google Scholar]
- Bolton, W.K.; Cattran, D.C.; Williams, M.E.; Adler, S.G.; Appel, G.B.; Cartwright, K.; Foiles, P.G.; Freedman, B.I.; Raskin, P.; Ratner, R.E.; et al. Randomized trial of an inhibitor of formation of advanced glycation end products in diabetic nephropathy. Am. J. Nephrol. 2004, 24, 32–40. [Google Scholar]
- Tupe, R.; Agte, V. Interaction of zinc, ascorbic acid, and folic acid in glycation with albumin as protein model. Biol. Trace Elem. Res. 2010, 138, 346–357. [Google Scholar]
- Ardestani, A.; Yazdanparast, R. Cyperus rotundus suppresses age formation and protein oxidation in a model of fructose-mediated protein glycoxidation. Int. J. Biol. Macromol. 2007, 41, 572–578. [Google Scholar]
- Iwanaga, A.; Kusano, G.; Warashina, T.; Miyase, T. Hyaluronidase inhibitors from “cimicifugae rhizoma” (a mixture of the rhizomes of cimicifuga dahurica and c. Heracleifolia). J. Nat. Prod. 2010, 73, 573–578. [Google Scholar]
- Sakai, S.; Kawamata, H.; Kogure, T.; Mantani, N.; Terasawa, K.; Umatake, M.; Ochiai, H. Inhibitory effect of ferulic acid and isoferulic acid on the production of macrophage inflammatory protein-2 in response to respiratory syncytial virus infection in raw264.7 cells. Mediators Inflamm. 1999, 8, 173–175. [Google Scholar]
- Hirabayashi, T.; Ochiai, H.; Sakai, S.; Nakajima, K.; Terasawa, K. Inhibitory effect of ferulic acid and isoferulic acid on murine interleukin-8 production in response to influenza virus infections in vitro and in vivo. Planta Med. 1995, 61, 221–226. [Google Scholar]
- Schmid, D.; Woehs, F.; Svoboda, M.; Thalhammer, T.; Chiba, P.; Moeslinger, T. Aqueous extracts of cimicifuga racemosa and phenolcarboxylic constituents inhibit production of proinflammatory cytokines in lps-stimulated human whole blood. Can. J. Physiol. Pharmacol. 2009, 87, 963–972. [Google Scholar]
- Sakai, S.; Ochiai, H.; Mantani, N.; Kogure, T.; Shibahara, N.; Terasawa, K. Administration of isoferulic acid improved the survival rate of lethal influenza virus pneumonia in mice. Mediators Inflamm. 2001, 10, 93–96. [Google Scholar]
- Hirata, A.; Murakami, Y.; Atsumi, T.; Shoji, M.; Ogiwara, T.; Shibuya, K.; Ito, S.; Yokoe, I.; Fujisawa, S. Ferulic acid dimer inhibits lipopolysaccharide-stimulated cyclooxygenase-2 expression in macrophages. In Vivo 2005, 19, 849–853. [Google Scholar]
- Wang, X.; Li, X.; Chen, D. Evaluation of antioxidant activity of isoferulic acid in vitro. Nat. Prod. Commun. 2011, 6, 1285–1288. [Google Scholar]
- Liu, I.M.; Hsu, F.L.; Chen, C.F.; Cheng, J.T. Antihyperglycemic action of isoferulic acid in streptozotocin-induced diabetic rats. Br. J. Pharmacol. 2000, 129, 631–636. [Google Scholar]
- Liu, I.M.; Chen, W.C.; Cheng, J.T. Mediation of beta-endorphin by isoferulic acid to lower plasma glucose in streptozotocin-induced diabetic rats. J. Pharmacol. Exp. Ther. 2003, 307, 1196–1204. [Google Scholar]
- Adisakwattana, S.; Sompong, W.; Meeprom, A.; Ngamukote, S.; Yibchok-anun, S. Cinnamic acid and its derivatives inhibit fructose-mediated protein glycation. Int. J. Mol. Sci. 2012, 13, 1778–1789. [Google Scholar]
- Booth, A.A.; Khalifah, R.G.; Todd, P.; Hudson, B.G. In vitro kinetic studies of formation of antigenic advanced glycation end products (AGEs). Novel inhibition of post-amadori glycation pathways. J. Biol. Chem. 1997, 272, 5430–5437. [Google Scholar]
- Goh, S.Y.; Cooper, M.E. Clinical review: The role of advanced glycation end products in progression and complications of diabetes. J. Clin. Endocrinol. MeTable 2008, 93, 1143–1152. [Google Scholar]
- Takagi, Y.; Kashiwagi, A.; Tanaka, Y.; Asahina, T.; Kikkawa, R.; Shigeta, Y. Significance of fructose-induced protein oxidation and formation of advanced glycation end product. J. Diabetes Complications 1995, 9, 87–91. [Google Scholar]
- Suárez, G.; Rajaram, R.; Oronsky, A.L.; Gawinowicz, M.A. Nonenzymatic glycation of bovine serum albumin by fructose (fructation). Comparison with the maillard reaction initiated by glucose. J. Biol. Chem. 1989, 264, 3674–3679. [Google Scholar]
- Shield, J.P.; Poyser, K.; Hunt, L.; Pennock, C.A. Fructosamine and glycated haemoglobin in the assessment of long term glycaemic control in diabetes. Arch. Dis. Child. 1994, 71, 443–445. [Google Scholar]
- Wu, C.H.; Huang, S.M.; Lin, J.A.; Yen, G.C. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011, 2, 224–234. [Google Scholar]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar]
- Hunt, J.V.; Wolff, S.P. Oxidative glycation and free radical production: A causal mechanism of diabetic complications. Free Radic. Res. Commun. 1991, 12–13, 115–123. [Google Scholar]
- Kalousová, M.; Zima, T.; Tesar, V.; Lachmanová, J. Advanced glycation end products and advanced oxidation protein products in hemodialyzed patients. Blood Purif. 2002, 20, 531–536. [Google Scholar]
- Aćimović, J.M.; Stanimirović, B.D.; Mandić, L.M. The role of the thiol group in protein modification with methylglyoxal. J. Serb. Chem. Soc. 2009, 74, 867–883. [Google Scholar]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar]
- Balu, M.; Sangeetha, P.; Murali, G.; Panneerselvam, C. Age-related oxidative protein damages in central nervous system of rats: Modulatory role of grape seed extract. Int. J. Dev. Neurosci. 2005, 23, 501–507. [Google Scholar]
- Cai, Y.-Z.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar]
- Peyroux, J.; Sternberg, M. Advanced glycation endproducts (AGEs): Pharmacological inhibition in diabetes. Pathol. Biol. 2006, 54, 405–419. [Google Scholar]
- Price, D.L.; Rhett, P.M.; Thorpe, S.R.; Baynes, J.W. Chelating activity of advanced glycation end-product inhibitors. J. Biol. Chem. 2001, 276, 48967–48972. [Google Scholar]
- Peng, X.; Ma, J.; Chen, F.; Wang, M. Naturally occurring inhibitors against the formation of advanced glycation end-products. Food Funct. 2011, 2, 289–301. [Google Scholar]
- Bouma, B.; Kroon-Batenburg, L.M.; Wu, Y.P.; Brünjes, B.; Posthuma, G.; Kranenburg, O.; de Groot, P.G.; Voest, E.E.; Gebbink, M.F. Glycation induces formation of amyloid cross-beta structure in albumin. J. Biol. Chem. 2003, 278, 41810–41819. [Google Scholar]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar]
- Marzban, L.; Soukhatcheva, G.; Verchere, C.B. Role of carboxypeptidase e in processing of pro-islet amyloid polypeptide in β-cells. Endocrinology 2005, 146, 1808–1817. [Google Scholar]
- Tokuyama, T.; Yagui, K.; Yamaguchi, T.; Huang, C.I.; Kuramoto, N.; Shimada, F.; Miyazaki, J.; Horie, H.; Saito, Y.; Makino, H.; et al. Expression of human islet amyloid polypeptide/amylin impairs insulin secretion in mouse pancreatic beta cells. Metabolism 1997, 46, 1044–1051. [Google Scholar]
- Marzban, L.; Verchere, C.B. The role of islet amyloid polypeptide in type 2 diabetes. Can. J. Diabetes 2004, 28, 39–47. [Google Scholar]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar]
- Tupe, R.S.; Agte, V.V. Role of zinc along with ascorbic acid and folic acid during long-term in vitro albumin glycation. Br. J. Nutr. 2010, 103, 370–377. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Meeprom, A.; Sompong, W.; Chan, C.B.; Adisakwattana, S. Isoferulic Acid, a New Anti-Glycation Agent, Inhibits Fructose- and Glucose-Mediated Protein Glycation in Vitro. Molecules 2013, 18, 6439-6454. https://doi.org/10.3390/molecules18066439
Meeprom A, Sompong W, Chan CB, Adisakwattana S. Isoferulic Acid, a New Anti-Glycation Agent, Inhibits Fructose- and Glucose-Mediated Protein Glycation in Vitro. Molecules. 2013; 18(6):6439-6454. https://doi.org/10.3390/molecules18066439
Chicago/Turabian StyleMeeprom, Aramsri, Weerachat Sompong, Catherine B. Chan, and Sirichai Adisakwattana. 2013. "Isoferulic Acid, a New Anti-Glycation Agent, Inhibits Fructose- and Glucose-Mediated Protein Glycation in Vitro" Molecules 18, no. 6: 6439-6454. https://doi.org/10.3390/molecules18066439