Bioactive Compounds from the Roots of Asiasarum heterotropoides


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
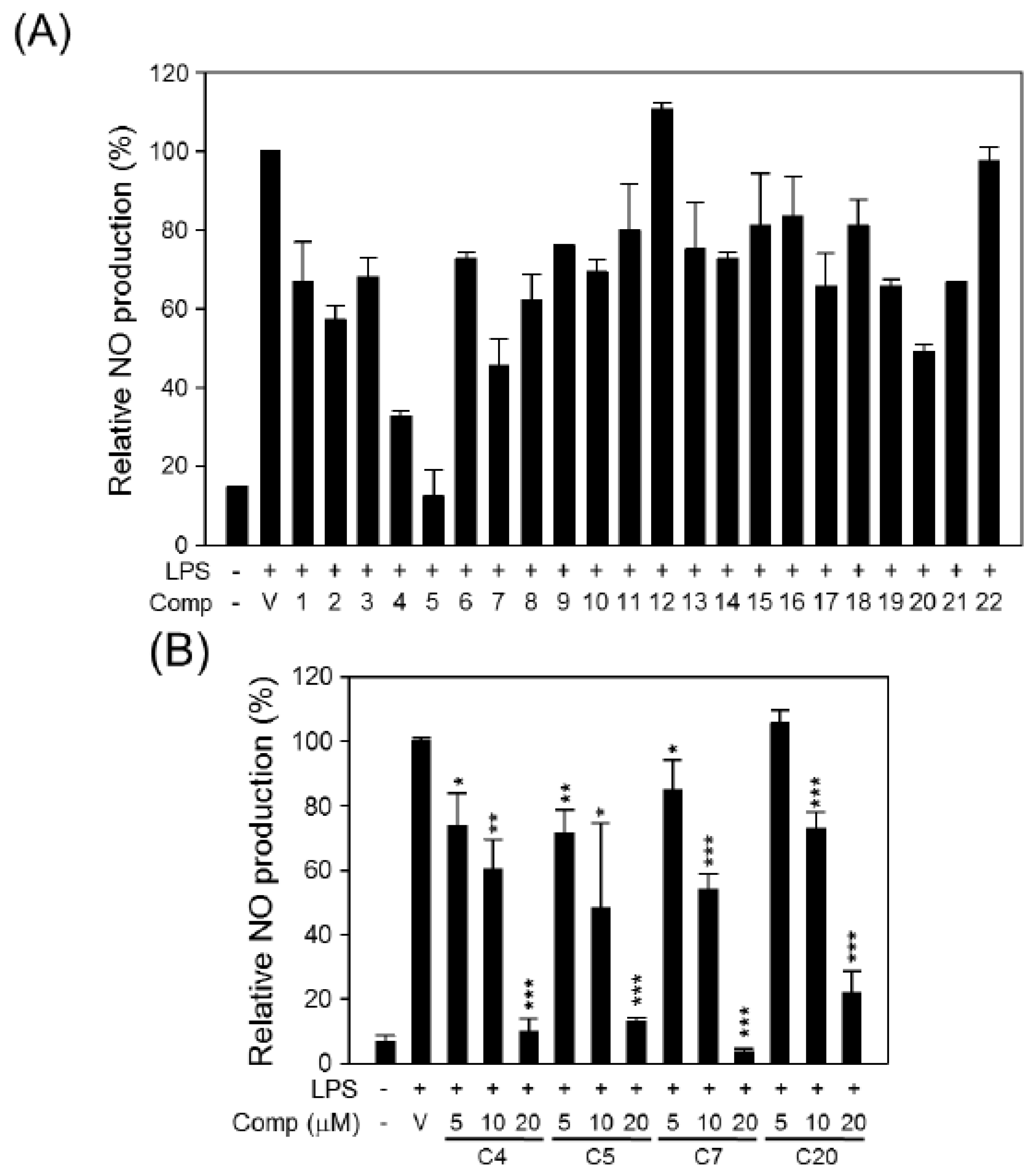
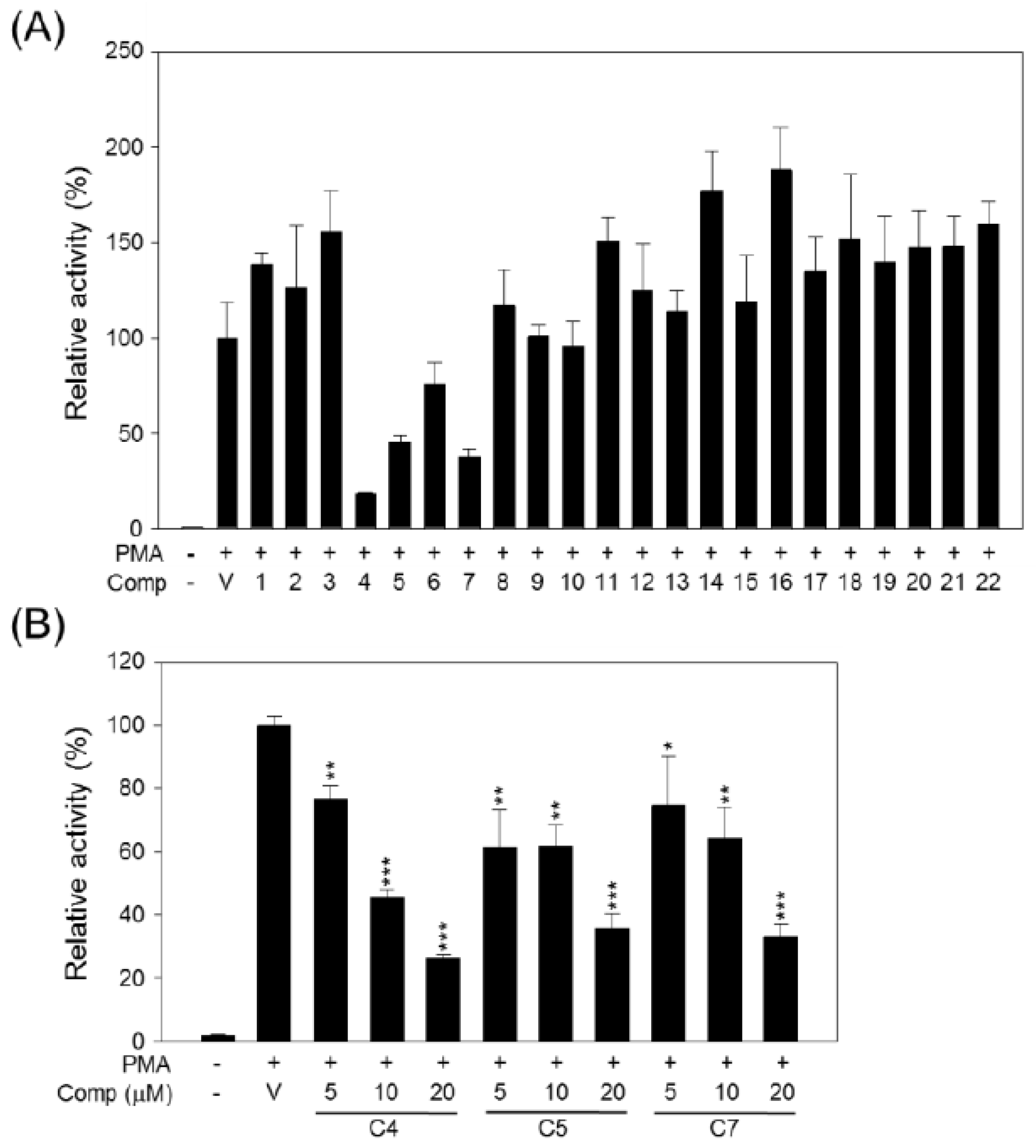
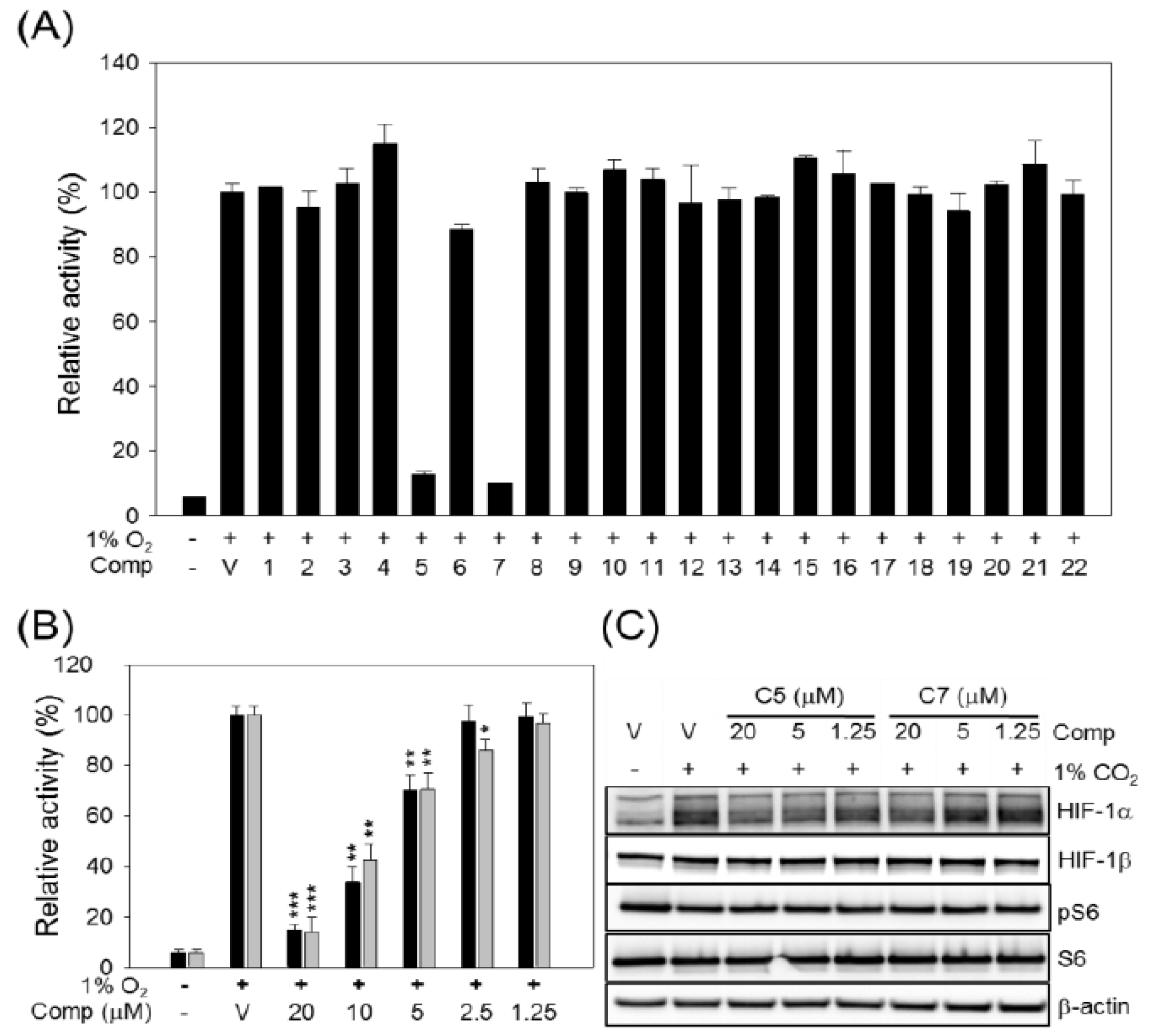
2. Results and Discussion



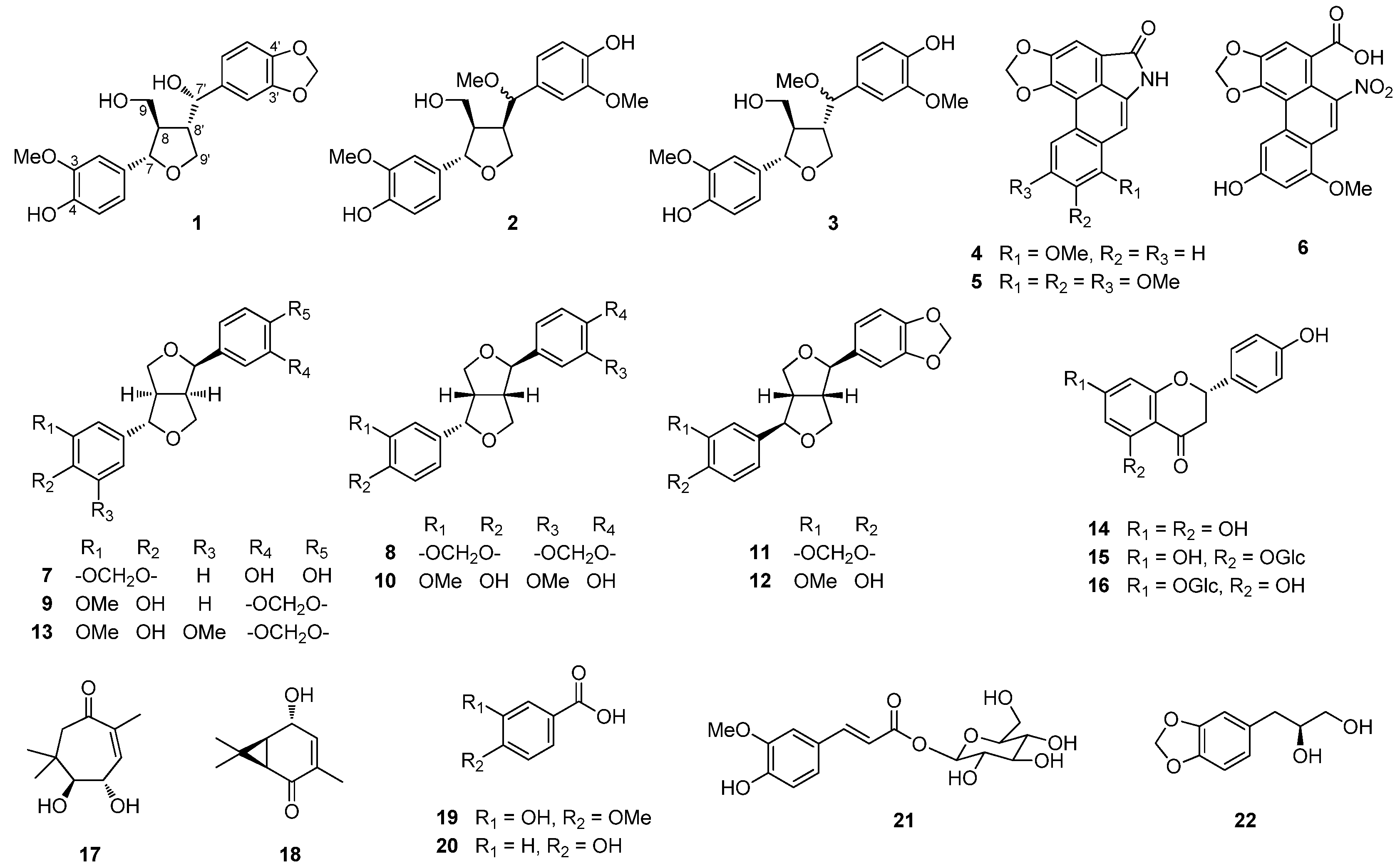
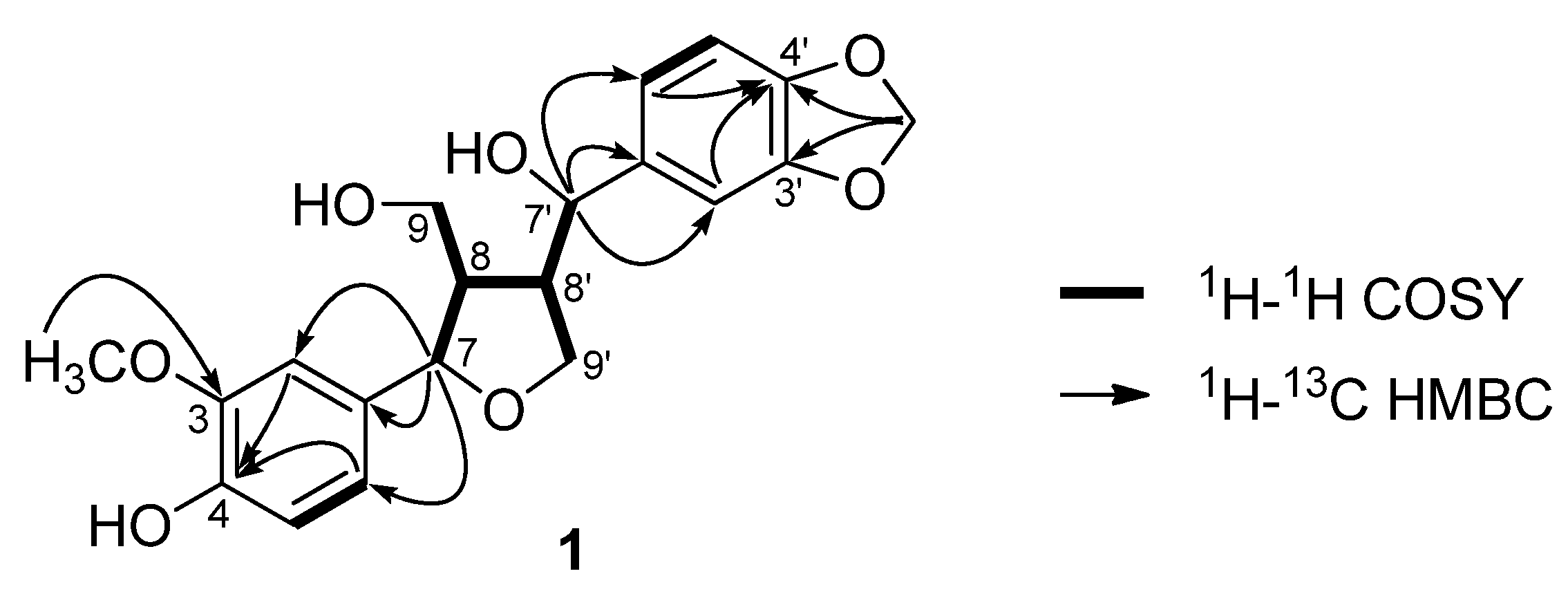
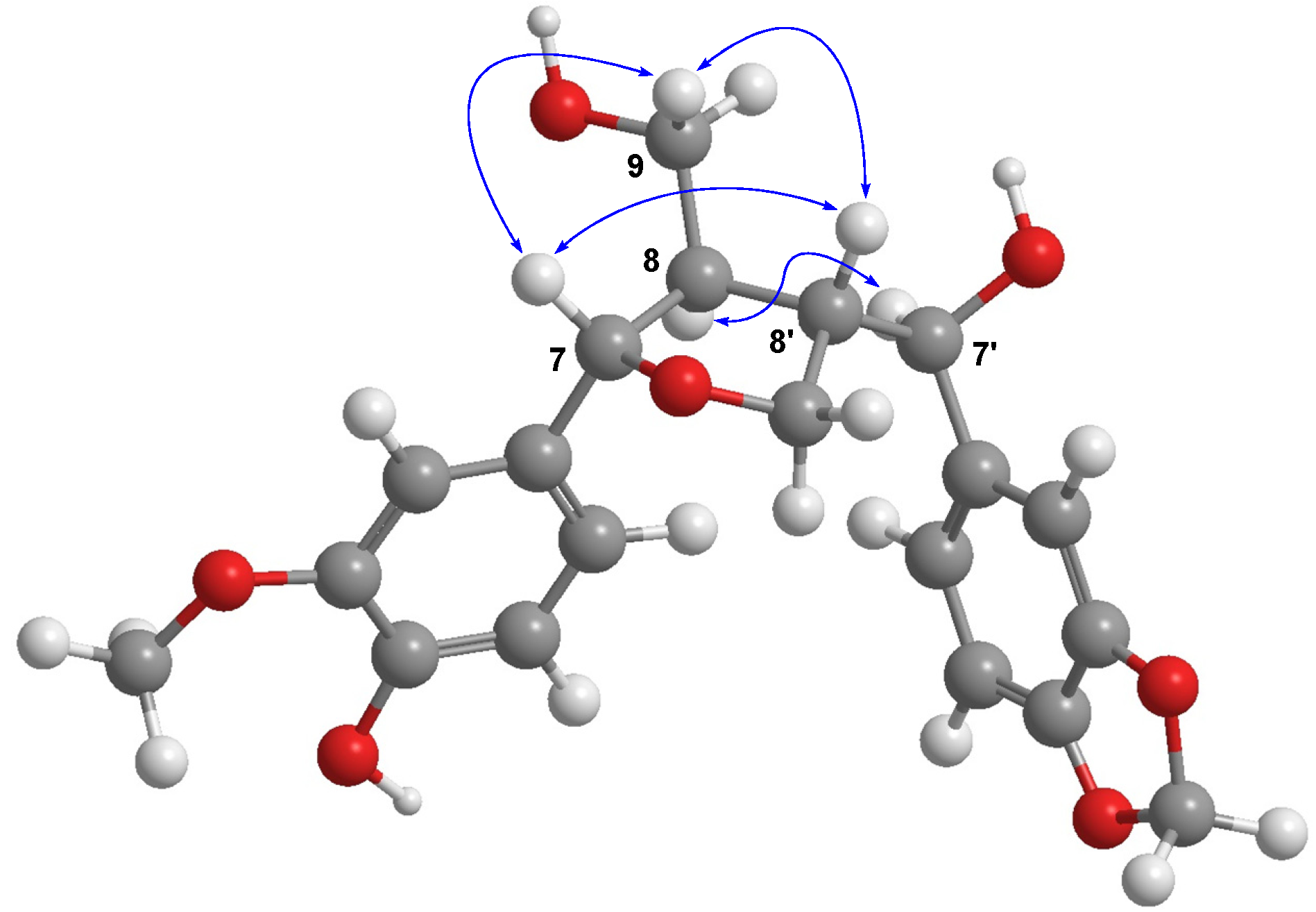
 +39.2, MeOH) showed positive signals at 245 and 293 nm [15,16,17]. Therefore, compound 1 was determined to be (7S,8R,7'S,8'S)-3-methoxy-3',4'-methylenedioxy-7,9'-epoxylignane-4,7',9-triol.
+39.2, MeOH) showed positive signals at 245 and 293 nm [15,16,17]. Therefore, compound 1 was determined to be (7S,8R,7'S,8'S)-3-methoxy-3',4'-methylenedioxy-7,9'-epoxylignane-4,7',9-triol.



3. Experimental
3.1. General
3.2. Plant Material
3.3. Extraction and Isolation
3.4. Characterization Data of (7S,8R,7'S,8'S)-3-Methoxy-3',4'-methylenedioxy-7,9'-epoxylignane-4,7',9-triol (1)
+39.2 (c 0.1, MeOH); UV (MeOH) λmax (log ε) 227 (4.92), 283 (4.44) nm; CD (MeOH, c = 5.3 × 10−3 M) Δε(nm): +19.9 (245), +8.4 (293); 1H-NMR (CDCl3, 500 MHz) δ 6.86 (3H, m, overlapping, H-2'/H-5/H-2), 6.79 (1H, m, overlapping, H-6), 6.77 (1H, m, overlapping, H-6'), 6.75 (1H, d, J = 8.0 Hz, H-5'), 5.96 (2H, s, -OCH2O-), 5.85 (1H, brs, 4-OH), 4.41 (1H, d, J = 9.5 Hz, H-7'), 4.34 (1H, d, J = 9.0 Hz, H-7), 3.89 (3H, s, OMe), 3.69 (1H, m, H-9a), 3.60 (2H, m, H-9') 3.56 (1H, m, H-9b), 2.54 (1H, m, H-8'), 2.25 (1H, m, H-8); 13C-NMR (CDCl3, 125 MHz) δ 148.0 (C-3'), 147.5 (C-4'), 146.8 (C-3), 145.6 (C-4), 136.8 (C-1'), 132.5 (C-1), 120.3 (C-6'), 119.7 (C-6), 114.3 (C-5), 108.8 (C-2), 108.1 (C-5'), 106.8 (C-2'), 101.2 (-OCH2O-), 84.5 (C-7), 76.9 (C-7'), 70.3 (C-9'), 63.1 (C-9), 56.0 (3-OMe), 55.7 (C-8), 53.5 (C-8'); HRESIMS m/z 397.1263 [M+Na]+ (calcd. for C20H22O7Na, 397.1263).3.5. Cell Culture
3.6. FOXP3-Lluciferase Stable Cell Line
3.7. Hypoxia Inducible Factor (HIF)-Luciferase Stable Cell Line
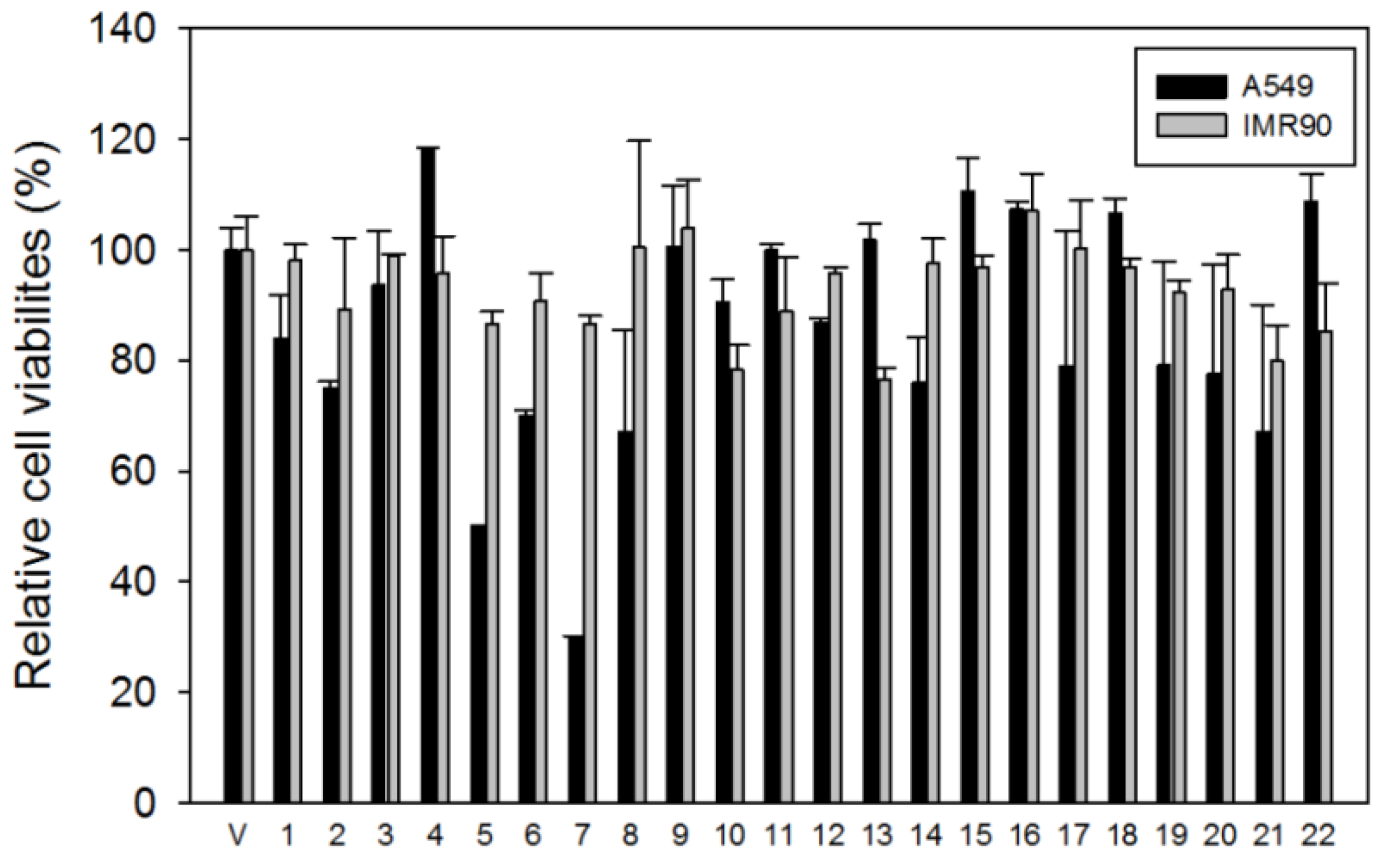
3.8. Cell Viability
3.9. NO Assay
3.10. Luciferase Reporter Assay
3.11. Western Blot Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ishizaki, S.; Narumi, T.; Mizushima, M.; Ohara, M. Effect of the specialist herbivore Luehdorfia puziloi on the performance of a woodland herbaceous plant, Asarum heterotropoides. Plant Spec. Biol. 2010, 25, 61–67. [Google Scholar] [CrossRef]
- Cai, S.-Q.; Yu, J.; Wang, X.; Wang, R.-Q.; Ran, F.-X.; Shang, M.-Y.; Cui, J.-R.; Komatsu, K.; Namba, T. Cytotoxic activity of some Asarum plants. Fitoterapia 2008, 79, 293–297. [Google Scholar] [CrossRef]
- Li, Y.-L.; Tian, M.; Yu, J.; Shang, M.-Y.; Cai, S.-Q. Studies on morphology and aristolochic acid analogue constituents of Asarum campaniflorum and a comparison with two official species of Asari radix et rhizoma. J. Nat. Med. 2010, 64, 442–451. [Google Scholar] [CrossRef]
- Suzuki, Y.; Yuzurihara, M.; Hibino, T.; Yano, S.; Kase, Y. Aqueous extract of Asiasari radix inhibits formalin-induced hyperalgesia via NMDA receptors. J. Ethnopharmacol. 2009, 123, 128–133. [Google Scholar] [CrossRef]
- Kim, H.M.; Moon, Y.S. Asiasari radix inhibits immunoglobulin E production on experimental models in vitro and in vivo. Immunopharmacol. Immunotoxicol. 1999, 21, 469–481. [Google Scholar] [CrossRef]
- Han, Y.; Kim, S.-J. Memory enhancing actions of Asiasari radix extracts via activation of insulin receptor and extracellular signal regulated kinase (ERK) I/II in rat hippocampus. Brain Res. 2003, 974, 193–201. [Google Scholar] [CrossRef]
- Rho, S.-S.; Park, S.-J.; Hwang, S.-L.; Lee, M.-H.; Kim, C.D.; Lee, I.-H.; Chang, S.-Y.; Rang, M.-J. The hair growth promoting effect of Asiasari radix extract and its molecular regulation. J. Dermatol. Sci. 2005, 38, 89–97. [Google Scholar] [CrossRef]
- Jang, J.-Y.; Lee, J.-H.; Shin, H.-K.; Choi, Y.-H.; Lee, J.-D.; Choi, B.-T. Partially purified Asiasari radix inhibits melanogenesis through extracellular signal-regulated kinase signaling in B16F10 cells. Int. J. Mol. Med. 2010, 25, 287–292. [Google Scholar]
- Oh, S.-M.; Kim, J.; Lee, J.; Yi, J.-M.; Oh, D.-S.; Bang, O.-S.; Kim, N.S. Anticancer potential of an ethanol extract of Asiasari radix against HCT-116 human colon cancer cells in vitro. Oncol. Lett. 2013, 5, 305–310. [Google Scholar]
- Dan, Y.; Liu, H.-Y.; Gao, W.-W.; Chen, S.-L. Activities of essential oils from Asarum heterotropoides var. Mandshuricum against five phytopathogens. Crop Prot. 2010, 29, 295–299. [Google Scholar] [CrossRef]
- Endo, J.; Nakamura, T. Studies in Asiasarum (aristolochiaceae). I. Three new compounds in Asiasarum heteropoides. Yakugaku Zasshi 1978, 98, 789–793. [Google Scholar]
- Takasaki, M.; Konoshima, T.; Yasuda, I.; Hamano, T.; Tokuda, H. Inhibitory effects of shouseiryu-to on two-stage carcinogenesis. II1. Anti-tumor-promoting activities of lignans from Asiasarum heterotropoides var. Mandshuricum. Biol. Pharm. Bull. 1997, 20, 776–780. [Google Scholar] [CrossRef]
- Huang, J.; Wang, H.Q.; Zhang, C.; Li, G.Y.; Lin, R.C.; Wang, J.H. A new tetrahydrofuran-type lignan with anti-inflammatory activity from Asarum heterotropoides fr. Schmidt var. Mandshuricum. J. Asian. Nat. Prod. Res. 2013. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Chang, K.S.; Park, C.; Ahn, Y.-J. Larvicidal activity of Asarum heterotropoides root constituents against insecticide-susceptible and -resistant Culex pipiens pallens and Aedes aegypti and Ochlerotatus togoi. J. Agric. Food. Chem. 2010, 58, 10001–10006. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.; Jang, D.S.; Nam, J.-W.; Kim, J.-P.; Park, K.H.; Yang, M.S.; Seo, E.-K. Two new stereoisomers of tetrahydrofuranoid lignans from the flower buds of Magnolia fargesii. Chem. Pharm. Bull. 2007, 55, 137–139. [Google Scholar] [CrossRef]
- Li, W.; Koike, K.; Liu, L.; Lin, L.; Fu, X.; Chen, Y.; Nikaido, T. New lignan glucosides from the stems of Tinospora sinensis. Chem. Pharm. Bull. 2004, 52, 638–640. [Google Scholar] [CrossRef]
- Macias, F.A.; Lopez, A.; Varela, R.M.; Torres, A.; Molinillo, J.M.G. Bioactive lignans from a cultivar of Helianthus annuus. J. Agric. Food. Chem. 2004, 52, 6443–6447. [Google Scholar] [CrossRef]
- Sun, Y.-J.; Li, Z.-L.; Chen, H.; Liu, X.-Q.; Zhou, W.; Hua, H.-M. Four new cytotoxic tetrahydrofuranoid lignans from Sinopodophyllum emodi. Planta Med. 2012, 78, 480–484. [Google Scholar] [CrossRef]
- Lee, H.S.; Han, D.S. Aristolactam derivaties and their N-glycosides from Aristolochia contorta. Korean J. Pharmacogn. 1993, 24, 32–37. [Google Scholar]
- Dong, S.-W.; Shang, M.-Y.; Wang., X.; Zhang, S.-X.; Li., C.; Cai, S.-Q. Chemical constituents isolated from Saruma henryi. J. Chin. Pharm. Sci. 2009, 18, 146–150. [Google Scholar]
- Ding, Z.-H.; Yiao, L.-H.; Chen, Z.-L.; Ding, J.-K. The chemical constituents of Asarum petelotii. Acta Bot. Yunnan. 1994, 16, 305–308. [Google Scholar]
- Zhang, J.; Xiao, Y.; Feng, J.; Wu, S.L.; Xue, X.; Zhang, X.; Liang, X. Selectively preparative purification of aristolochic acids and aristololactams from Aristolochia plants. J. Pharm. Biomed. Anal. 2010, 52, 446–451. [Google Scholar]
- Tomimori, N.; Nakai, M.; Ono, Y.; Kitagawa, Y.; Kiso, Y.; Shibata, H. Identification of the metabolites of episesamin in rat bile and human liver microsomes. Biol. Pharm. Bull. 2012, 35, 709–716. [Google Scholar] [CrossRef]
- Choi, E.H.; Choi, J.Y.; Lee, J.K.; Oh, J.S.; Kim, D.C.; Lee, H.S.; Son, J.K.; Ryang, S.A.; Kim, J.A.; Lee, S.H. Isolation of Melanin Biosynthesis inhibitory compounds from the Roots of Asarum sieboldii. Korean J. Pharmacogn. 2007, 38, 394–399. [Google Scholar]
- Swain, N.A.; Brown, R.C.; Bruton, G. A versatile stereoselective synthesis of endo, exo-furofuranones: Application to the enantioselective synthesis of furofuran lignans. J. Org. Chem. 2004, 69, 122–129. [Google Scholar] [CrossRef]
- Meselhy, M.R. Constituents from Moghat, the roots of glossostemon bruguieri (desf.). Molecules 2003, 8, 614–621. [Google Scholar] [CrossRef]
- Roy, S.C.; Rana, K.K.; Guin, C. Short and stereoselective total synthesis of furano lignans (+/−)-dihydrosesamin, (+/−)-lariciresinol dimethyl ether, (+/−)-acuminatin methyl ether, (+/−)-sanshodiol methyl ether, (+/−)-lariciresinol, (+/−)-acuminatin, and (+/−)-lariciresinol monomethyl ether and furofuran lignans (+/−)-sesamin, (+/−)-eudesmin, (+/−)-piperitol methyl ether, (+/−)-pinoresinol, (+/−)-piperitol, and (+/−)-pinoresinol monomethyl ether by radical cyclization of epoxides using a transition-metal radical source. J. Org. Chem. 2002, 67, 3242–3248. [Google Scholar] [CrossRef]
- Ina, H.; Ono, M.; Sashida, Y.; Iida, H. (+)-Piperitol from Paulownia tomentosa. Planta Med. 1987, 53, 504. [Google Scholar] [CrossRef]
- Quang, T.H.; Ngan, N.T.T.; Minh, C.V.; Kiem, P.V.; Tai, B.H.; Thao, N.P.; Song, S.B.; Kim, Y.H. Anti-inflammatory and PPAR transactivational effects of secondary metabolites from the roots of Asarum sieboldii. Bioorg. Med. Chem. Lett. 2012, 22, 2527–2533. [Google Scholar] [CrossRef]
- Andrade, C.A.; Carvalho, J.L.S.; Cunico, M.M.; Lordello, A.L.L.; Higaskino, C.E.K.; Almeida, S.C.C.; Dias, J.F.G.; Kerber, V.A.; Miguel, M.D.; Miguel, O.G. Antioxidant and antibacterial activity of extracts, fractions and isolated substances from the flowers of Acacia podalyriifolia A. Cunn. ex g. Don. Braz. J. Pharm. Sci. 2010, 46, 715–721. [Google Scholar]
- Shimoda, K.; Kubota, N.; Taniuchi, K.; Sato, D.; Nakajima, N.; Hamada, H.; Hamada, H. Biotransformation of naringin and naringenin by cultured Eucalyptus perriniana cells. Phytochemistry 2010, 71, 201–205. [Google Scholar] [CrossRef]
- Baek, S.W.; Kim, E.R.; Kim, J.; Kim, Y.C. Chemical constituents of Abies kireana leaves with inhibitory activity against nitric oxide production in BV2 microglia cells. Nat. Prod. Sci. 2011, 17, 175–180. [Google Scholar]
- Hashimoto, K.; Katsuhara, T.; Niitsu, K.; Ikeya, Y.; Hayashi, K.; Maruno, M.; Fujita, T. Enantioexcess monoterpenes from roots of Asiasarum sieboldi. Phytochemistry 1994, 35, 969–973. [Google Scholar] [CrossRef]
- Oh, J.; Hwang, I.H.; Kim, D.C.; Kang, S.-C.; Jang, T.-S.; Lee, S.H.; Na, M. Anti-listerial compounds from Asari radix. Arch. Pharm.Res. 2010, 33, 1339–1345. [Google Scholar] [CrossRef]
- Lee, C.-K.; Lee, P.-H.; Kuo, Y.-H. The chemical constituents from the aril of Cassia fistula L. J. Chin. Chem. Soc. 2001, 48, 1053–1058. [Google Scholar]
- Youn, U.J.; Lee, Y.J.; Jeon, H.R.; Shin, H.J.; Son, Y.M.; Nam, J.-W.; Han, A.R.; Seo, E.-K. A prydyl alkaloid and benzoic acid deriviatives from the rhizomes of Anemarrhena asphodeloides. Nat. Prod. Sci. 2010, 16, 203–206. [Google Scholar]
- Materska, M.; Perucka, I.; Stochmal, A.; Piacente, S.; Oleszek, W. Quantitative and qualitative determination of flavonoids and phenolic acid derivatives from pericarp of hot pepper fruit cv. Bronowicka ostra. Pol. J. Food Nutr. Sci. 2003, 12, 72–76. [Google Scholar]
- Sawant, R.T.; Waghmode, S.B. Organocatalytic approach to (S)-1-Arylpropan-2-ols: Enantioselective synthesis of the key intermediate of antiepileptic agent (−)-talampanel. Synth. Commun. 2010, 40, 2269–2277. [Google Scholar]
- Jadeski, L.C.; Hum, K.O.; Chakraborty, C.; Lala, P.K. Nitric oxide promotes murine mammary tumour growth and metastasis by stimulating tumour cell migration, invasiveness and angiogenesis. Int. J. Cancer 2000, 86, 30–39. [Google Scholar]
- Al, D.Y.; Attoub, S.; Arafat, K.; Abuqamar, S.; Viallet, J.; Saleh, A.; Al, A.H.; Eid, A.; Iratni, R. Anti-metastatic and anti-tumor growth effects of origanum majorana on highly metastatic human breast cancer cells: Inhibition of NFκB signaling and reduction of nitric oxide production. PloS One 2013, 8, e68808. [Google Scholar]
- Gallo, O.; Masini, E.; Morbidelli, L.; Franchi, A.; Fini-Storchi, I.; Vergari, W.A.; Ziche, M. Role of nitric oxide in angiogenesis and tumor progression in head and neck cancer. J. Natl. Cancer Inst. 1998, 90, 587–596. [Google Scholar]
- Punathil, T.; Katiyar, S.K. Inhibition of non-small cell lung cancer cell migration by grape seed proanthocyanidins is mediated through the inhibition of nitric oxide, guanylate cyclase, and ERK1/2. Mol. Carcinog. 2009, 48, 232–242. [Google Scholar]
- Lala, P.K.; Chakraborty, C. Role of nitric oxide in carcinogenesis and tumour progression. Lancet Oncol. 2001, 2, 149–156. [Google Scholar]
- Saleem, W.; Suzuki, Y.; Mobaraki, A.; Yoshida, Y.; Noda, S.; Saitoh, J.I.; Nakano, T. Reduction of nitric oxide level enhances the radiosensitivity of hypoxic non-small cell lung cancer. Cancer Sci. 2011, 102, 2150–2156. [Google Scholar]
- Li, B.; Samanta, A.; Song, X.; Furuuchi, K.; Iacono, K.T.; Kennedy, S.; Katsumata, M.; Saouaf, S.J.; Greene, M.I. Foxp3 ensembles in T-cell regulation. Immunol. Rev. 2006, 212, 99–113. [Google Scholar]
- Fu, H.Y.; Li, C.; Yang, W.; Gai, X.D.; Jia, T.; Lei, Y.M.; Li, Y. Foxp3 and TLR4 protein expression are correlated in non-small cell lung cancer: Implications for tumor progression and escape. Acta Histochem. 2013, 115, 151–157. [Google Scholar]
- Xia, M.; Zhao, M.Q.; Wu, K.; Lin, X.Y.; Liu, Y.; Qin, Y.J. Investigations on the clinical significance of FOXP3 protein expression in cervical oesophageal cancer and the number of FOXP3+ tumour-infiltrating lymphocytes. J. Int. Med. Res. 2013, 41, 1002–1008. [Google Scholar]
- Wang, L.; Zhang, B.; Xu, X.; Zhang, S.; Yan, X.; Kong, F.; Feng, X.; Wang, J. Clinical significance of FOXP3 expression in human gliomas. Clin. Transl. Oncol. 2013. [Google Scholar] [CrossRef]
- Lal, A.; Chan, L.; Devries, S.; Chin, K.; Scott, G.K.; Benz, C.C.; Chen, Y.Y.; Waldman, F.M.; Hwang, E.S. FOXP3-positive regulatory T lymphocytes and epithelial FOXP3 expression in synchronous normal, ductal carcinoma in situ, and invasive cancer of the breast. Breast Cancer Res. Treat. 2013, 139, 381–390. [Google Scholar]
- Li, C.; Yang, W.; Gai, X.; Zhang, Y.; Li, Y.; Fu, H. Foxp3 overexpression decreases sensitivity to chemotherapy in mouse Lewis lung cancer cells. Mol. Med. Rep. 2012, 6, 977–982. [Google Scholar]
- Li, W.; Katoh, H.; Wang, L.; Yu, X.; Du, Z.; Yan, X.; Zheng, P.; Liu, Y. FOXP3 regulates sensitivity of cancer cells to irradiation by transcriptional repression of BRCA1. Cancer Res. 2013, 73, 2170–2180. [Google Scholar]
- Zhang, C.Y.; Wang, X.; Su, T.; Ma, C.M.; Wen, Y.J.; Shang, M.Y.; Li, X.M.; Liu, G.X.; Cai, S.Q. New aristolochic acid, aristololactam and renal cytotoxic constituents from the stem and leaves of Aristolochia contorta. Pharmazie 2005, 60, 785–788. [Google Scholar]
- Wen, Y.J.; Su, T.; Tang, J.W.; Zhang, C.Y.; Wang, X.; Cai, S.Q.; Li, X.M. Cytotoxicity of phenanthrenes extracted from Aristolochia contorta in human proximal tubular epithelial cell line. Nephron Exp. Nephrol. 2006, 103, e95–e102. [Google Scholar]
- Zong, Y.; Sun, L.; Liu, B.; Deng, Y.S.; Zhan, D.; Chen, Y.L.; He, Y.; Liu, J.; Zhang, Z.J.; Sun, J.; et al. Resveratrol inhibits LPS-induced MAPKs activation via activation of the phosphatidylinositol 3-kinase pathway in murine RAW 264.7 macrophage cells. PloS One 2012, 7, e44107. [Google Scholar]
- Aizman, E.; Mor, A.; Levy, A.; George, J.; Kloog, Y. Ras inhibition by FTS attenuates brain tumor growth in mice by direct antitumor activity and enhanced reactivity of cytotoxic lymphocytes. Oncotarget 2012, 3, 144–157. [Google Scholar]
- Yonekura, S.; Itoh, M.; Okuhashi, Y.; Takahashi, Y.; Ono, A.; Nara, N.; Tohda, S. Effects of the HIF1 inhibitor, echinomycin, on growth and NOTCH signalling in leukaemia cells. Anticancer Res. 2013, 33, 3099–3103. [Google Scholar]
- Cardeno, A.; Sanchez-Hidalgo, M.; Rosillo, M.A.; Alarcon de la Lastra, C. Oleuropein, a secoiridoid derived from olive tree, inhibits the proliferation of human colorectal cancer cell through downregulation of HIF-1α. Nutr. Cancer 2013, 65, 147–156. [Google Scholar]
- Shin, H.W.; Cho, C.H.; Kim, T.Y.; Park, J.W. Sunitinib deregulates tumor adaptation to hypoxia by inhibiting HIF-1alpha synthesis in HT-29 colon cancer cells. Biochem. Biophys. Res. Commun. 2010, 398, 205–211. [Google Scholar]
- Shen, Y.; Wu, Y.; Chen, M.; Shen, W.; Huang, S.; Zhang, L.; Zou, X. Effects of pantoprazole as a HIF-1α inhibitor on human gastric adenocarcinoma sgc-7901 cells. Neoplasma 2012, 59, 142–149. [Google Scholar]
- Burroughs, S.K.; Kaluz, S.; Wang, D.; Wang, K.; van Meir, E.G.; Wang, B. Hypoxia inducible factor pathway inhibitors as anticancer therapeutics. Future Med. Chem. 2013, 5, 553–572. [Google Scholar]
- Xia, Y.; Choi, H.K.; Lee, K. Recent advances in hypoxia-inducible factor (HIF)-1 inhibitors. Eur. J. Med. Chem. 2012, 49, 24–40. [Google Scholar]
- Kim, H.P.; Leonard, W.J. Creb/atf-dependent t cell receptor-induced FoxP3 gene expression: A role for DNA methylation. J. Exp. Med. 2007, 204, 1543–1551. [Google Scholar]
- Lee, Y.J.; Kim, N.S.; Kim, H.; Yi, J.-M.; Oh, S.-M.; Bang, O.-S.; Lee, J. Cytotoxic and anti-inflammatory constituents from the seeds of Descurainia sophia. Arch. Pharm. Res. 2013, 36, 536–541. [Google Scholar]
- Sample Availability: Not available.
© 2013 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, J.; Lee, Y.J.; Oh, S.-M.; Yi, J.-M.; Kim, N.S.; Bang, O.-S. Bioactive Compounds from the Roots of Asiasarum heterotropoides. Molecules 2014, 19, 122-138. https://doi.org/10.3390/molecules19010122
Lee J, Lee YJ, Oh S-M, Yi J-M, Kim NS, Bang O-S. Bioactive Compounds from the Roots of Asiasarum heterotropoides. Molecules. 2014; 19(1):122-138. https://doi.org/10.3390/molecules19010122
Chicago/Turabian StyleLee, Jun, You Jin Lee, Se-Mi Oh, Jin-Mu Yi, No Soo Kim, and Ok-Sun Bang. 2014. "Bioactive Compounds from the Roots of Asiasarum heterotropoides" Molecules 19, no. 1: 122-138. https://doi.org/10.3390/molecules19010122