Anti-Inflammatory Effect of Methylpenicinoline from a Marine Isolate of Penicillium sp. (SF-5995): Inhibition of NF-κB and MAPK Pathways in Lipopolysaccharide-Induced RAW264.7 Macrophages and BV2 Microglia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
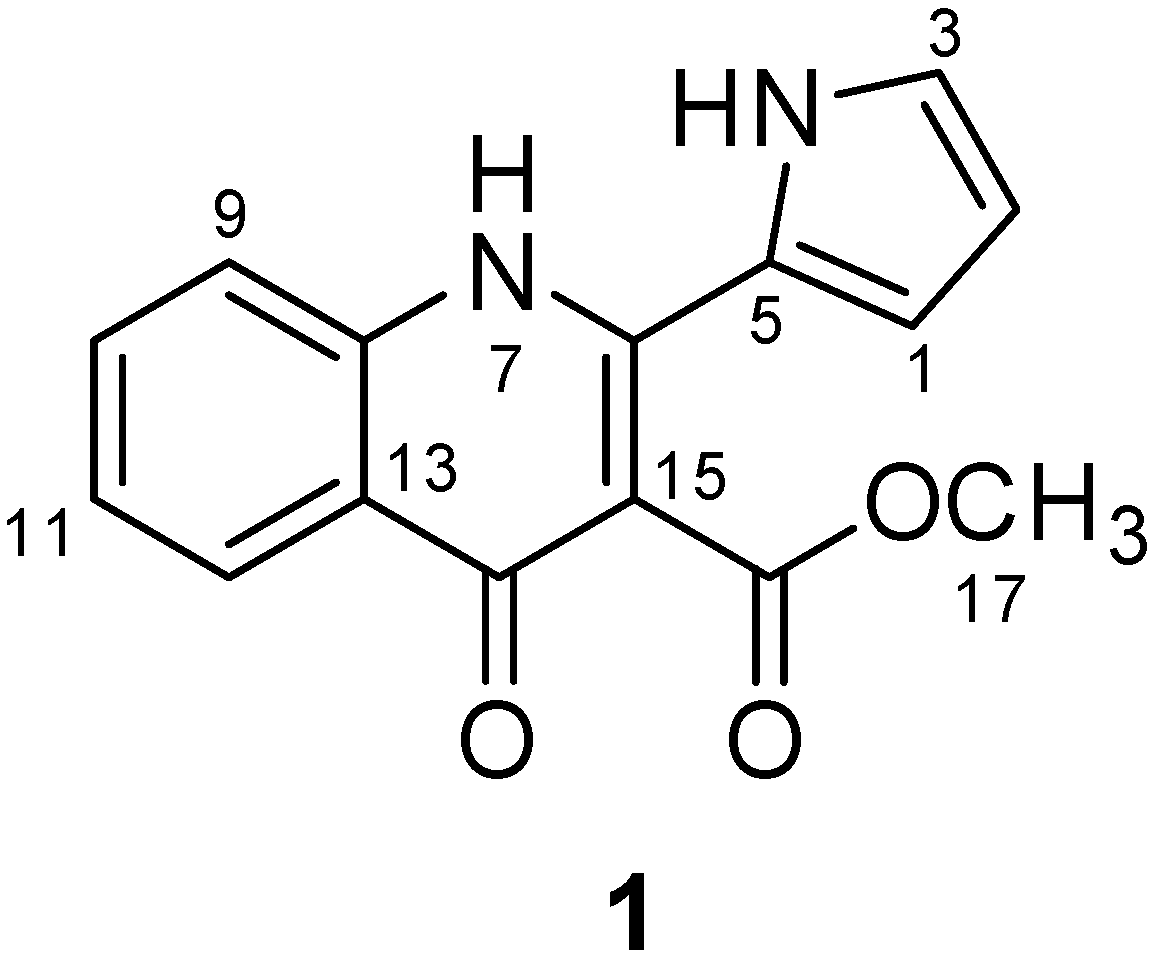
2.1. Isolation and Structure Determination of Methylpenicinoline (1)

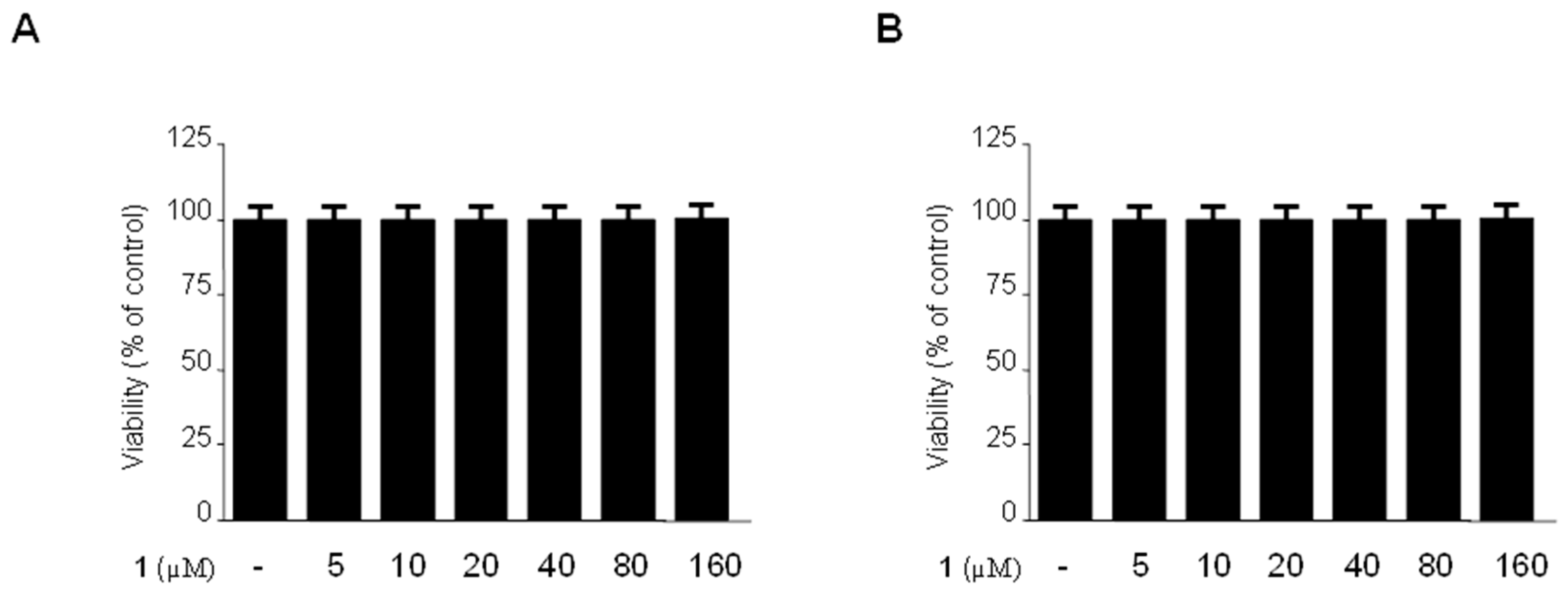
2.2. Effects of Methylpenicinoline (1) on the Viability of Mouse-Derived RAW264.7 and BV2 Cells

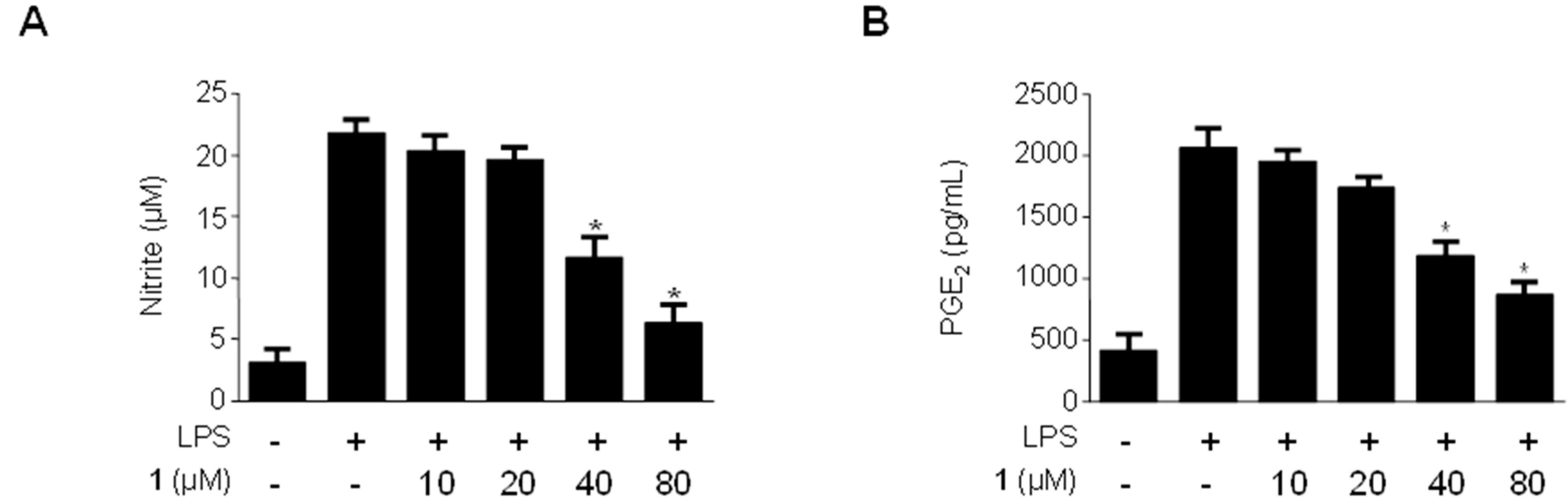
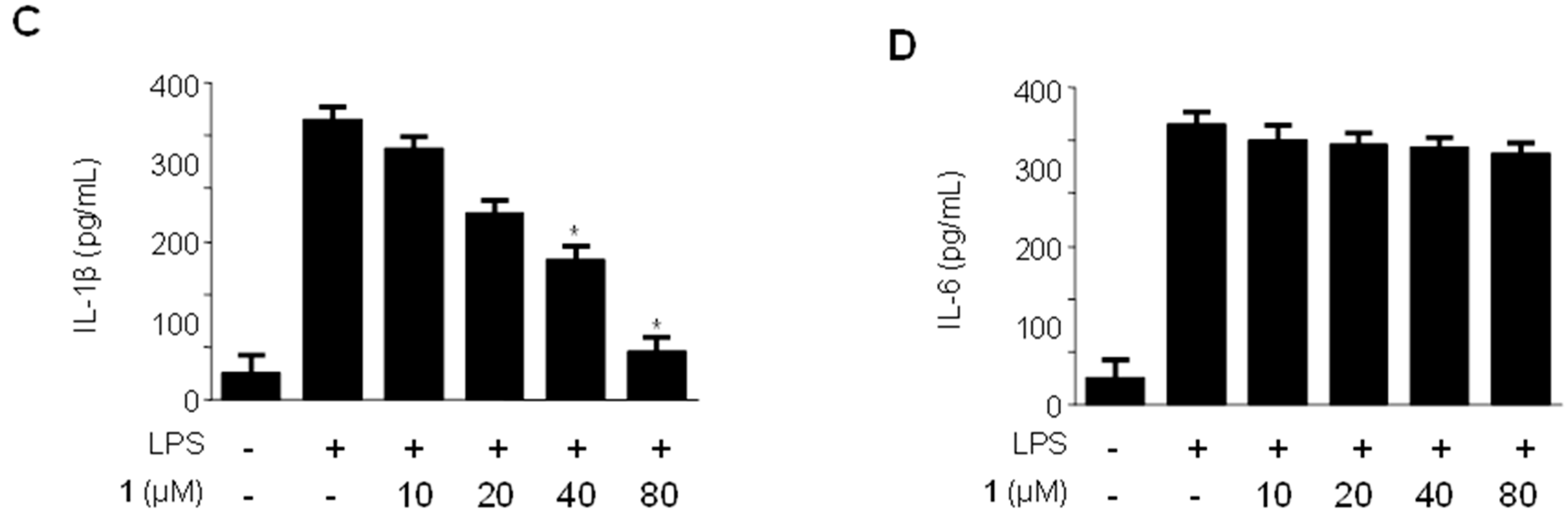
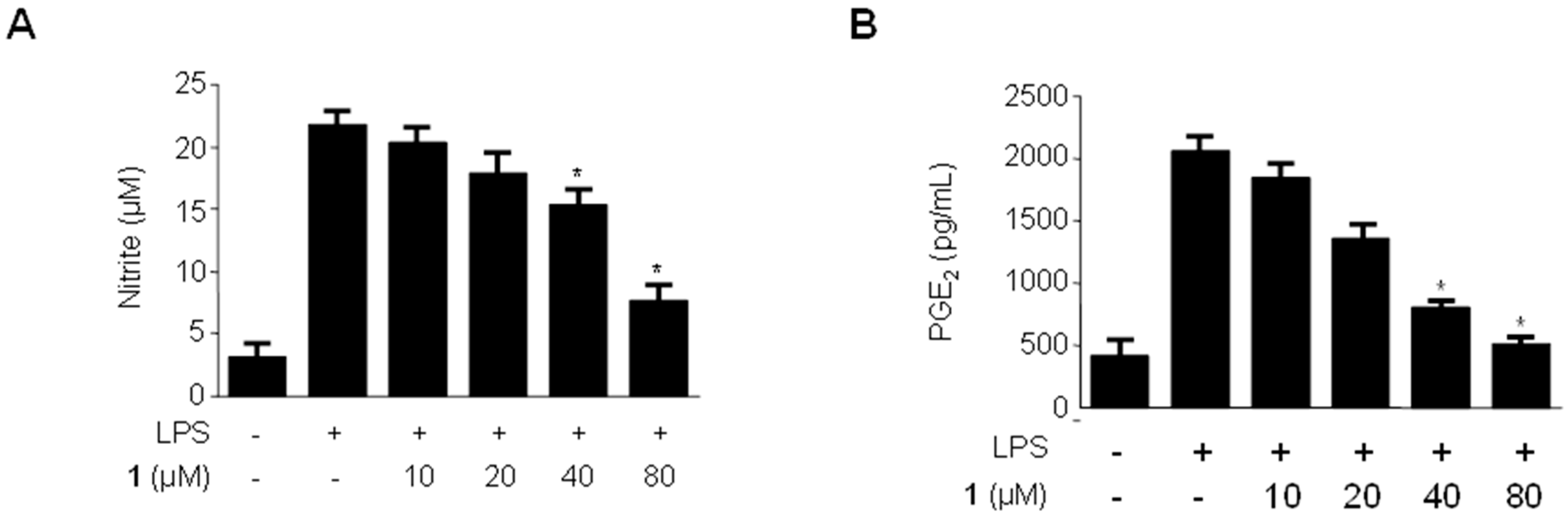
2.3. Effects of Methylpenicinoline (1) on the Production of Pro-Inflammatory Mediators and Cytokines in RAW264.7 Macrophages Stimulated with LPS


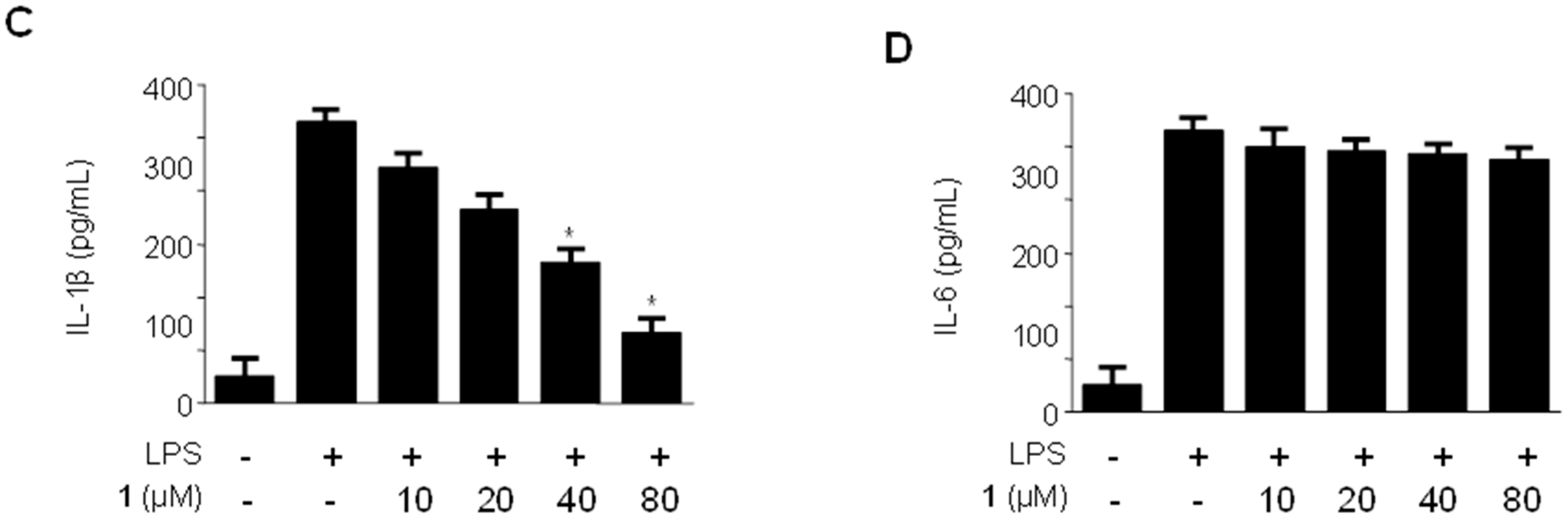
2.4. Effects of Methylpenicinoline (1) on the Production of Pro-Inflammatory Mediators and Cytokines in BV2 Microglia Stimulated with LPS



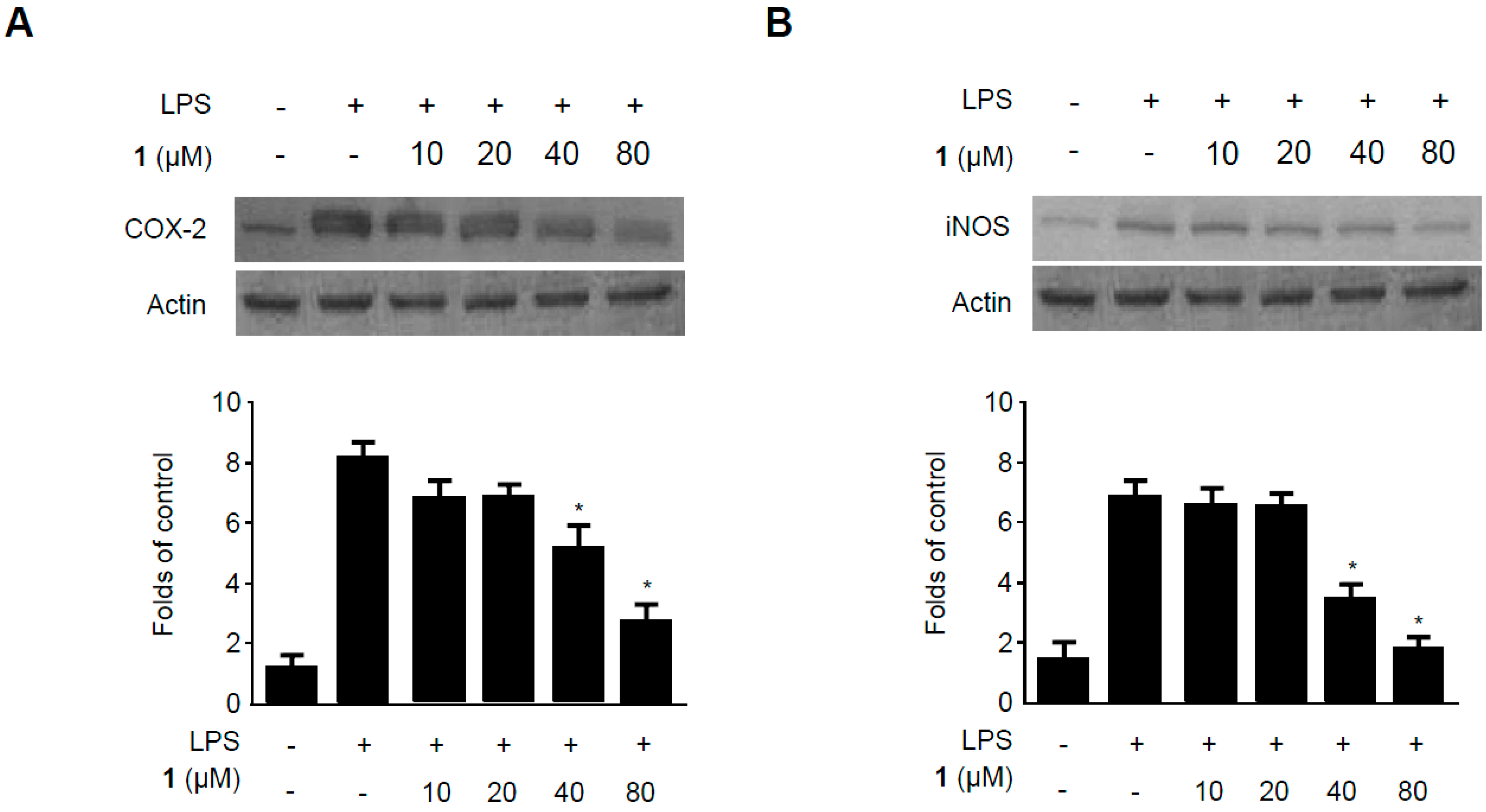
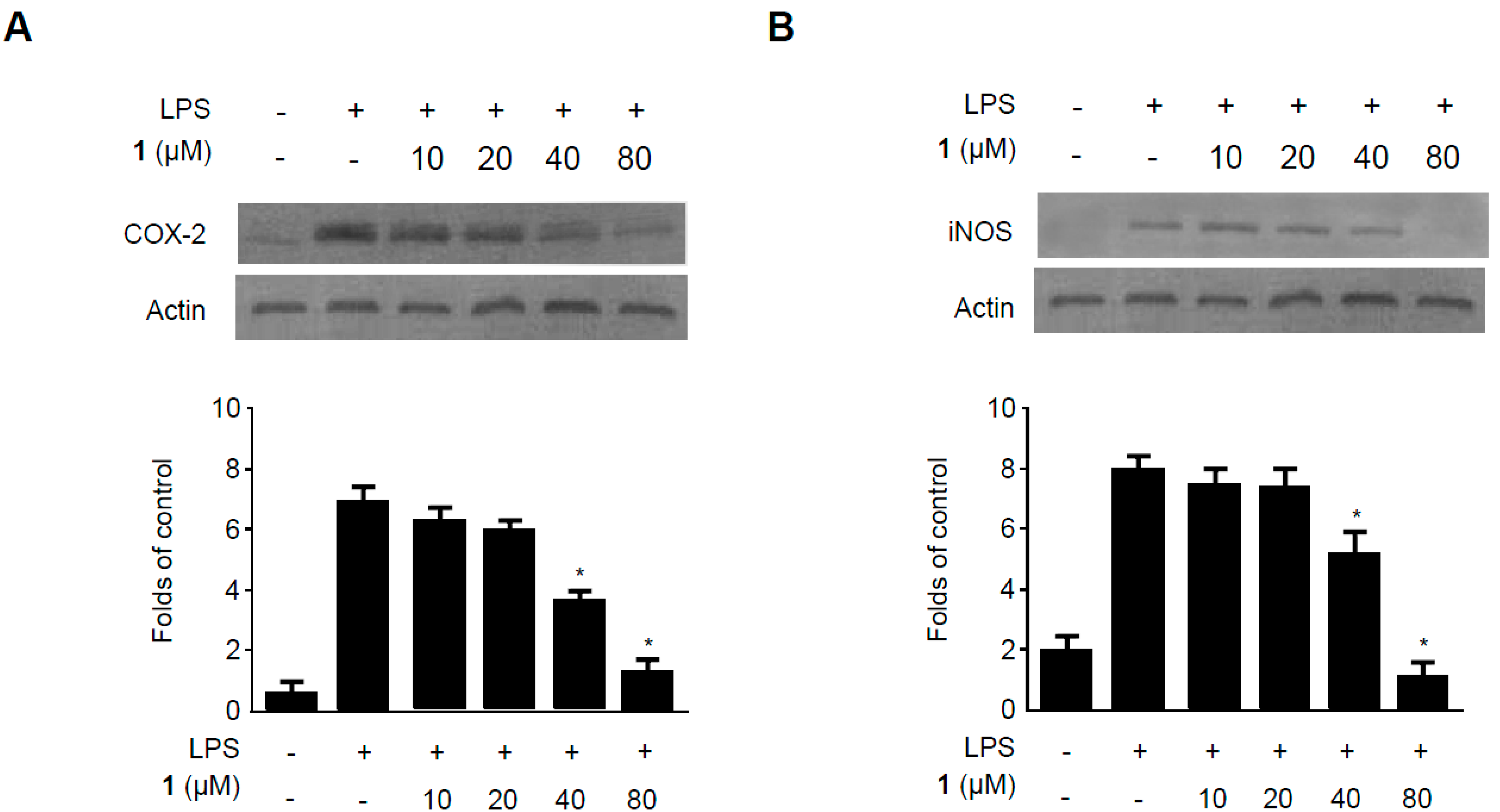
2.5. Effects of Methylpenicinoline (1) on the Expression of Pro-Inflammatory Enzymes in RAW264.7 Macrophages and BV2 Microglia Stimulated with LPS

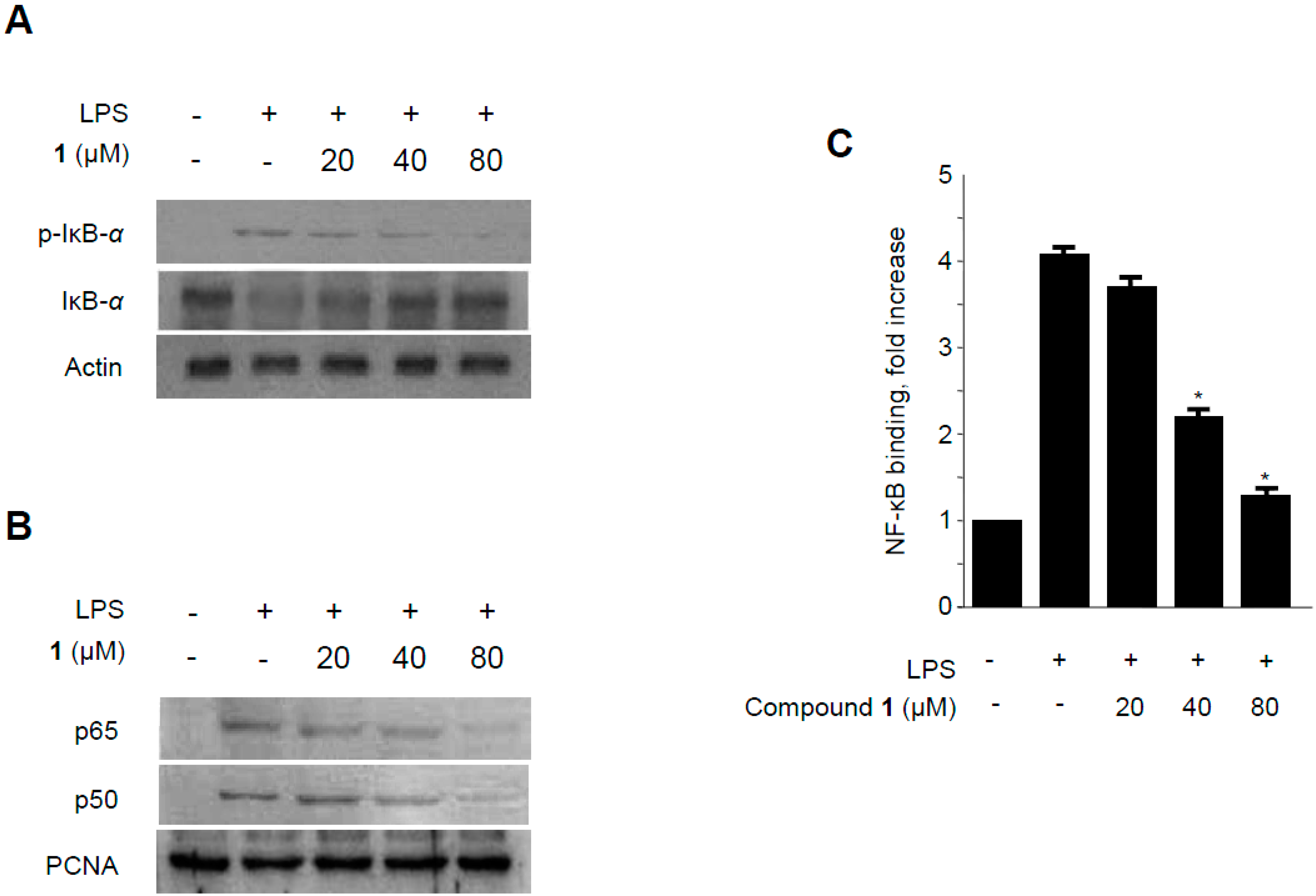
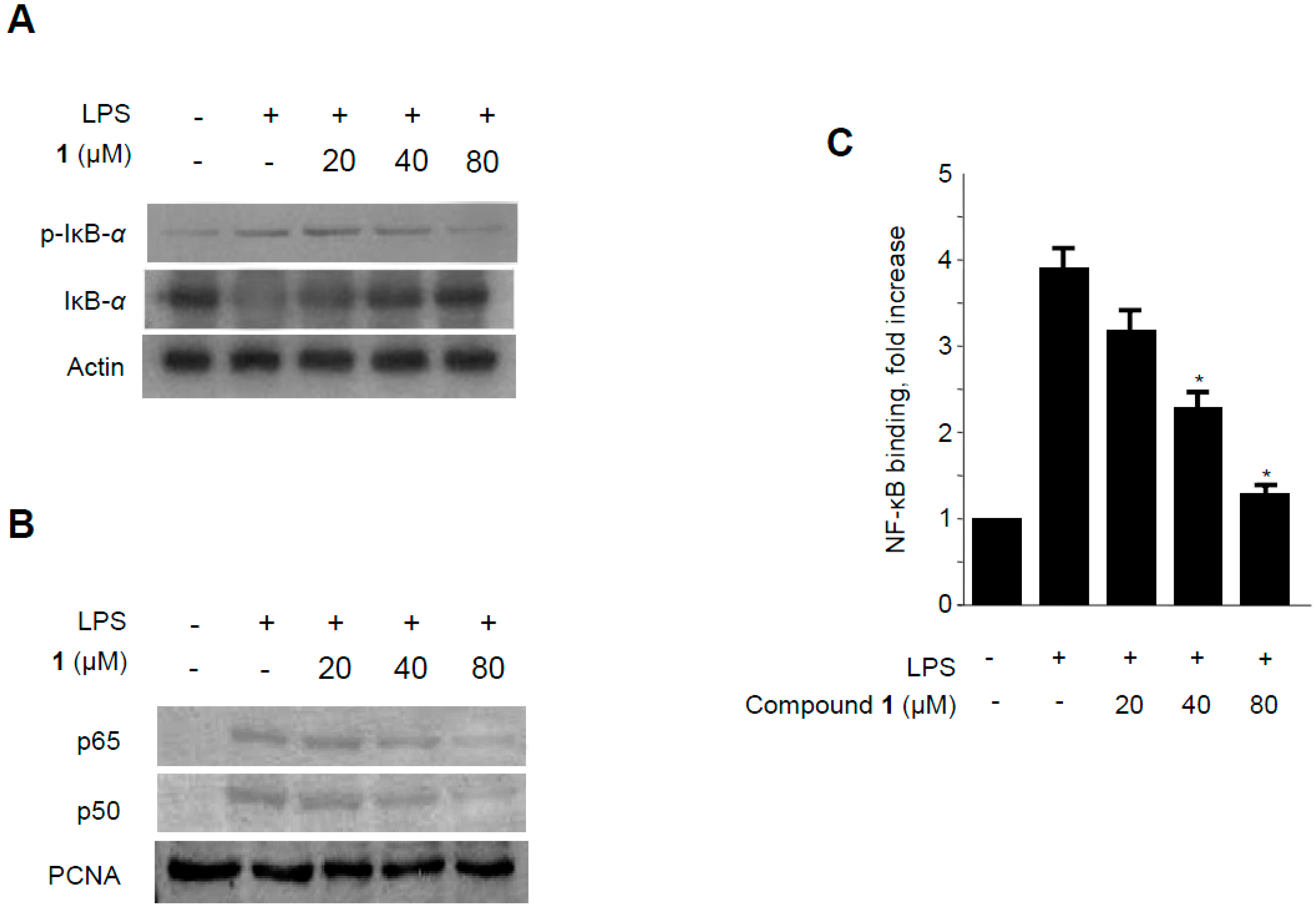
2.6. Effects of Methylpenicinoline (1) on NF-κB Activation in RAW264.7 Macrophages and BV2 Microglia Stimulated with LPS


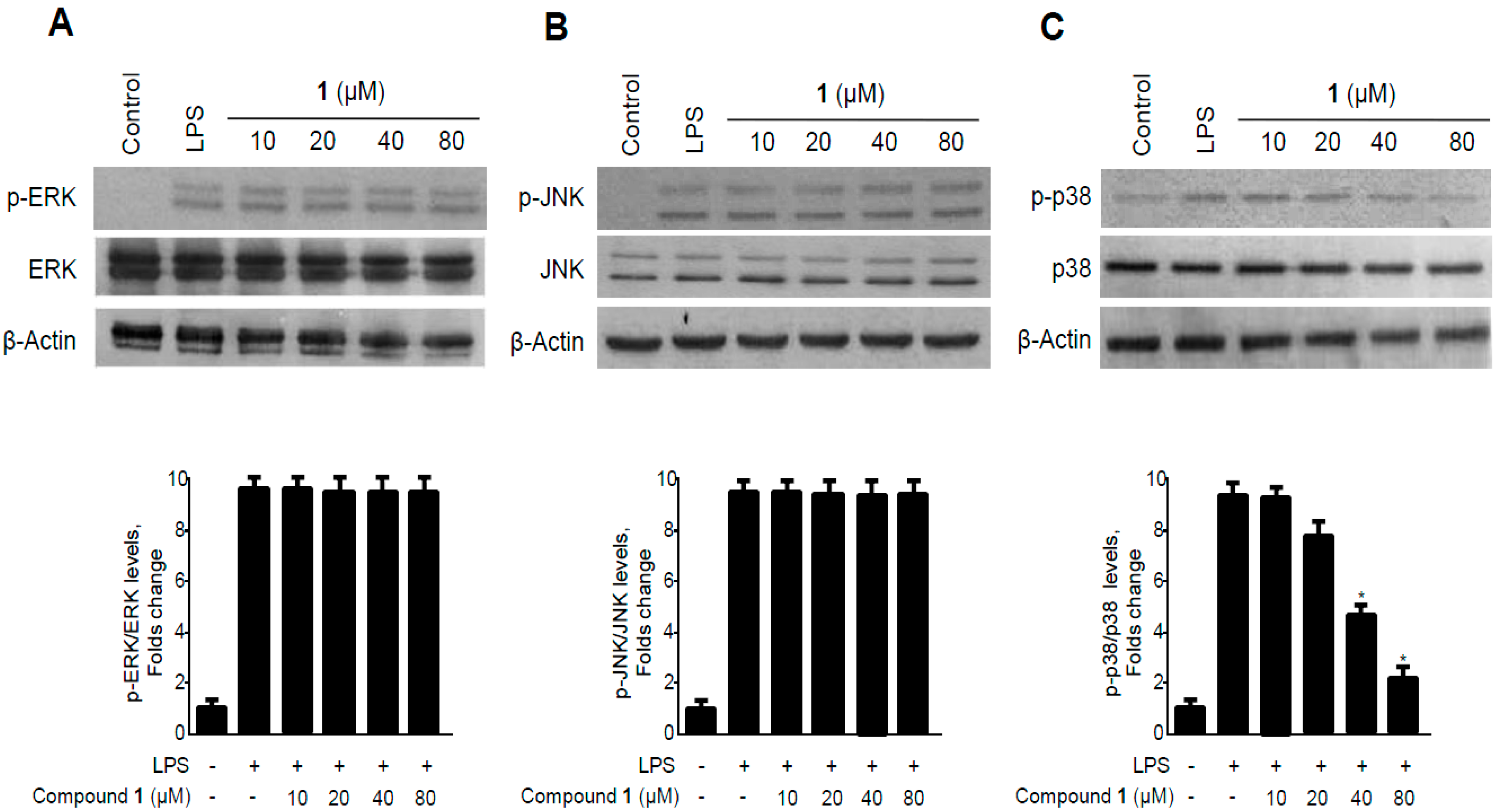
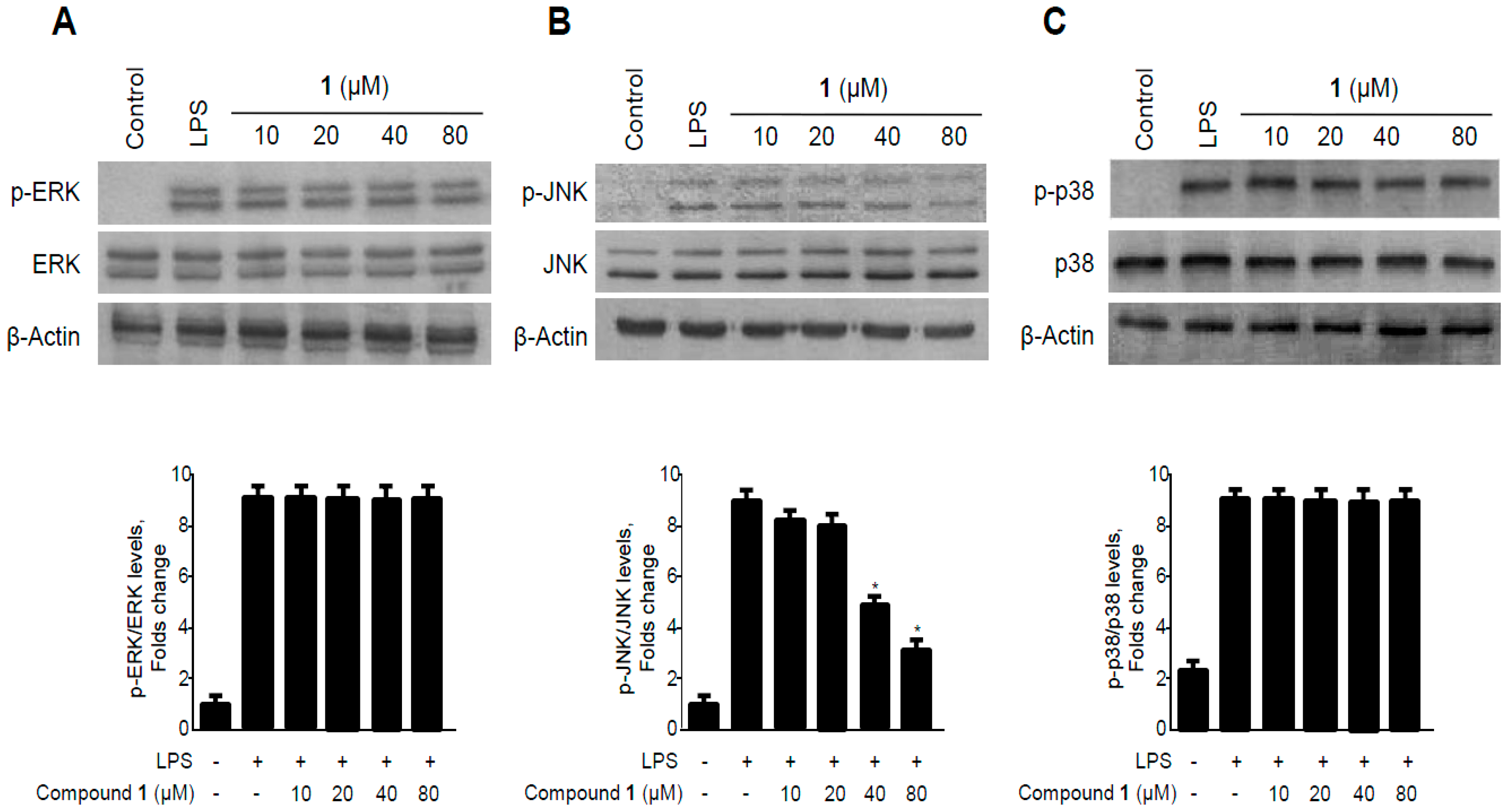
2.7. Effects of Methylpenicinoline (1) on the Phosphorylation of MAPKs in RAW264.7 Macrophages and BV2 Microglia Stimulated with LPS


3. Experimental Section
3.1. General
3.2. Specimen Collection and Identification of the Marine-Derived Fungus Penicillium sp. SF-5995
3.3. Fermentation, Extraction, and Isolation of Methylpenicinoline (1) from Penicillium sp. SF-5995
3.4. Cell Culture and Viability Assay
3.5. Determination of the Production of Nitrite, PGE2, IL-6, and IL-1β
3.6. Preparation of Cytosolic and Nuclear Fractions
3.7. Western Blot Analysis
3.8. DNA Binding Activity of NF-κB
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fujihara, M.; Muroi, M.; Tanamoto, K.; Suzuki, T.; Azuma, H.; Ikeda, H. Molecular mechanisms of macrophage activation and deactivation by lipopolysaccharide: Roles of the receptor complex. Pharmacol. Ther. 2003, 100, 171–194. [Google Scholar]
- Doyle, S.L.; O’Neill, L.A. Toll-like receptors: From the discovery of NFkappaB to new insights into transcriptional regulations in innate immunity. Biochem. Pharmacol. 2006, 72, 1102–1113. [Google Scholar]
- Brown, G.C.; Bal-Price, A. Inflammatory neurodegeneration mediated by nitric oxide, glutamate, and mitochondria. Mol. Neurobiol. 2003, 27, 325–355. [Google Scholar]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar]
- Bocchini, V.; Mazzolla, R.; Barluzzi, R.; Blasi, E.; Sick, P.; Kettenmann, H.J. An immortalized cell line expresses properties of activated microglial cells. J. Neurosci. Res. 1992, 31, 616–621. [Google Scholar]
- Choi, R.J.; Chun, J.; Khan, S.; Kim, Y.S. Desoxyrhapontigenin, a potent anti-inflammatory phytochemical, inhibits LPS-induced inflammatory responses via suppressing NF-κB and MAPK pathways in RAW264.7 cells. Int. Immunopharmacol. 2014, 18, 182–190. [Google Scholar]
- Rogerio, A.P.; Sá-Nunes, A.; Faccioli, L.H. The activity of medicinal plants and secondary metabolites on eosinophilic inflammation. Pharmacol. Res. 2010, 62, 298–307. [Google Scholar]
- Pan, M.H.; Lai, C.S.; Ho, C.T. Anti-inflammatory activity of natural dietary flavonoids. Food Funct. 2010, 1, 15–31. [Google Scholar]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; De Girolamo, M.; Cusenza, S.; Riccioni, G. Marine bioactives: Pharmacological properties and potential applications against inflammatory diseases. Mar. Drugs 2012, 10, 812–833. [Google Scholar]
- Senthilkumar, K.; Kim, S.K. Marine invertebrate natural products for anti-inflammatory and chronic diseases. Evid. Based Complement. Altern. Med. 2013, 2013, 572859. [Google Scholar]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar]
- Saleem, M.; Ali, M.S.; Hussain, S.; Jabbar, A.; Ashraf, M.; Lee, Y.S. Marine natural products of fungal origin. Nat. Prod. Rep. 2007, 24, 1142–1152. [Google Scholar]
- Lee, D.S.; Jeong, G.S.; Li, B.; Lee, S.U.; Oh, H.; Kim, Y.C. Asperlin from the marine-derived fungus Aspergillus sp. SF-5044 exerts anti-inflammatory effects through heme oxygenase-1 expression in murine macrophages. J. Pharmacol. Sci. 2011, 116, 283–295. [Google Scholar]
- Lee, D.S.; Jang, J.H.; Ko, W.; Kim, K.S.; Sohn, J.H.; Kang, M.S.; Ahn, J.S.; Kim, Y.C.; Oh, H. PTP1B inhibitory and anti-inflammatory effects of secondary metabolites isolated from the marine-derived fungus Penicillium sp. JF-55. Mar. Drugs 2013, 11, 1409–1426. [Google Scholar]
- Lee, S.U.; Asami, Y.; Lee, D.; Jang, J.-H.; Ahn, J.S.; Oh, H. Protuboxepins A and B and protubonines A and B from the marine-derived fungus Aspergillus sp. SF-5044. J. Nat. Prod. 2011, 74, 1284–1287. [Google Scholar]
- Elsebai, M.F.; Rempel, V.; Schnakenburg, G.; Kehraus, S.; Müller, C.E.; König, G.M. Identification of a potent and selective cannabinoid cb1 receptor antagonist from Auxarthron reticulatum. ACS Med. Chem. Lett. 2011, 2, 866–869. [Google Scholar]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar]
- Ohshima, H.; Bartsch, H. Chronic infections and inflammatory processes as cancer risk factors: Possible role of nitric oxide in carcinogenesis. Mutat. Res. 1994, 305, 253–264. [Google Scholar]
- Mancino, A.; Lawrence, T. Nuclear factor-kappaB and tumor-associated macrophages. Clin. Cancer Res. 2010, 16, 784–789. [Google Scholar]
- St-Onge, M.; Flamand, N.; Biarc, J.; Picard, S.; Bouchard, L.; Dussault, A.A.; Laflamme, C.; James, M.J.; Caughey, G.E.; Cleland, L.G.; et al. Characterization of prostaglandin E2 generation through the cyclooxygenase (COX)-2 pathway in human neutrophils. Biochim. Biophys. Acta 2007, 1771, 1235–1245. [Google Scholar]
- Turini, M.E.; DuBois, R.N. Cyclooxygenase-2: A therapeutic target. Annu. Rev. Med. 2002, 53, 35–57. [Google Scholar]
- Rocca, B.; FitzGerald, G.A. Cyclooxygenases and prostaglandins: Shaping up the immune response. Int. Immunopharmacol. 2002, 2, 603–630. [Google Scholar]
- Singh, V.P.; Patil, C.S.; Jain, N.K.; Kulkarni, S.K. Aggravation of inflammatory bowel disease by cyclooxygenase-2 inhibitors in rats. Pharmacology 2004, 72, 77–84. [Google Scholar]
- Lynch, M.A. The multifaceted profile of activated microglia. Mol. Neurobiol. 2009, 40, 139–156. [Google Scholar]
- Murphy, S. Production of nitric oxide by glial cells: Regulation and potential roles in the CNS. Glia 2000, 29, 1–13. [Google Scholar]
- Heo, S.K.; Yun, H.J.; Noh, E.K.; Park, W.H.; Park, S.D. LPS induces inflammatory responses in human aortic vascular smooth muscle cells via Toll-like receptor 4 expression and nitric oxide production. Immunol. Lett. 2008, 120, 57–64. [Google Scholar]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar]
- Kim, J.B.; Han, A.R.; Park, E.Y.; Kim, J.Y.; Cho, W.; Lee, J.; Seo, E.K.; Lee, K.T. Inhibition of LPS-induced iNOS, COX-2 and cytokines expression by poncirin through the NF-kappaB inactivation in RAW264.7 macrophage cells. Biol. Pharm. Bull. 2007, 30, 2345–2351. [Google Scholar]
- Moynagh, P.N. The NF-kappaB pathway. J. Cell Sci. 2005, 118, 4589–4592. [Google Scholar]
- Gomez, P.F.; Pillinger, M.H.; Attur, M.; Marjanovic, N.; Dave, M.; Park, J.; Bingham, C.O., 3rd; Al-Mussawir, H.; Abramson, S.B. Resolution of inflammation: Prostaglandin E2 dissociates nuclear trafficking of individual NF-kappaB subunits (p65, p50) in stimulated rheumatoid synovial fibroblasts. J. Immunol. 2005, 175, 6924–6230. [Google Scholar]
- Strayhorn, W.D.; Wadzinski, B.E. A novel in vitro assay for deubiquitination of I kappa B alpha. Arch. Biochem. Biophys. 2002, 40, 76–84. [Google Scholar]
- Bondeson, J. The mechanisms of action of disease-modifying antirheumatic drugs: A review with emphasis on macrophage signal transduction and the induction of proinflammatory cytokines. Gen. Pharmacol. 1997, 29, 127–150. [Google Scholar]
- Zhang, Y.; Dong, C. Regulatory mechanisms of mitogen-activated kinase signaling. Cell. Mol. Life Sci. 2007, 64, 2771–2789. [Google Scholar]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar]
- Titheradge, M.A. The enzymatic measurement of nitrate and nitrite. Methods Mol. Biol. 1998, 100, 83–91. [Google Scholar]
- Gao, H.; Zhang, L.; Zhu, T.; Gu, Q.; Li, D. Unusual pyrrolyl 4-quinolinone alkaloids from the marine-derived fungus Penicillium sp. ghq208. Chem. Pharm. Bull. 2012, 60, 1458–1460. [Google Scholar]
- Kim, S.H.; Smith, C.J.; Van Eldik, L.J. Importance of MAPK pathways for microglial pro-inflammatory cytokine IL-1 beta production. Neurobiol. Aging 2004, 25, 431–439. [Google Scholar]
- Sample Availability: Samples of the compound methylpenicinoline is available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-C.; Lee, H.-S.; Ko, W.; Lee, D.-S.; Sohn, J.H.; Yim, J.H.; Kim, Y.-C.; Oh, H. Anti-Inflammatory Effect of Methylpenicinoline from a Marine Isolate of Penicillium sp. (SF-5995): Inhibition of NF-κB and MAPK Pathways in Lipopolysaccharide-Induced RAW264.7 Macrophages and BV2 Microglia. Molecules 2014, 19, 18073-18089. https://doi.org/10.3390/molecules191118073
Kim D-C, Lee H-S, Ko W, Lee D-S, Sohn JH, Yim JH, Kim Y-C, Oh H. Anti-Inflammatory Effect of Methylpenicinoline from a Marine Isolate of Penicillium sp. (SF-5995): Inhibition of NF-κB and MAPK Pathways in Lipopolysaccharide-Induced RAW264.7 Macrophages and BV2 Microglia. Molecules. 2014; 19(11):18073-18089. https://doi.org/10.3390/molecules191118073
Chicago/Turabian StyleKim, Dong-Cheol, Hee-Suk Lee, Wonmin Ko, Dong-Sung Lee, Jae Hak Sohn, Joung Han Yim, Youn-Chul Kim, and Hyuncheol Oh. 2014. "Anti-Inflammatory Effect of Methylpenicinoline from a Marine Isolate of Penicillium sp. (SF-5995): Inhibition of NF-κB and MAPK Pathways in Lipopolysaccharide-Induced RAW264.7 Macrophages and BV2 Microglia" Molecules 19, no. 11: 18073-18089. https://doi.org/10.3390/molecules191118073