Saikosaponin D Isolated from Bupleurum falcatum Inhibits Selectin-Mediated Cell Adhesion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
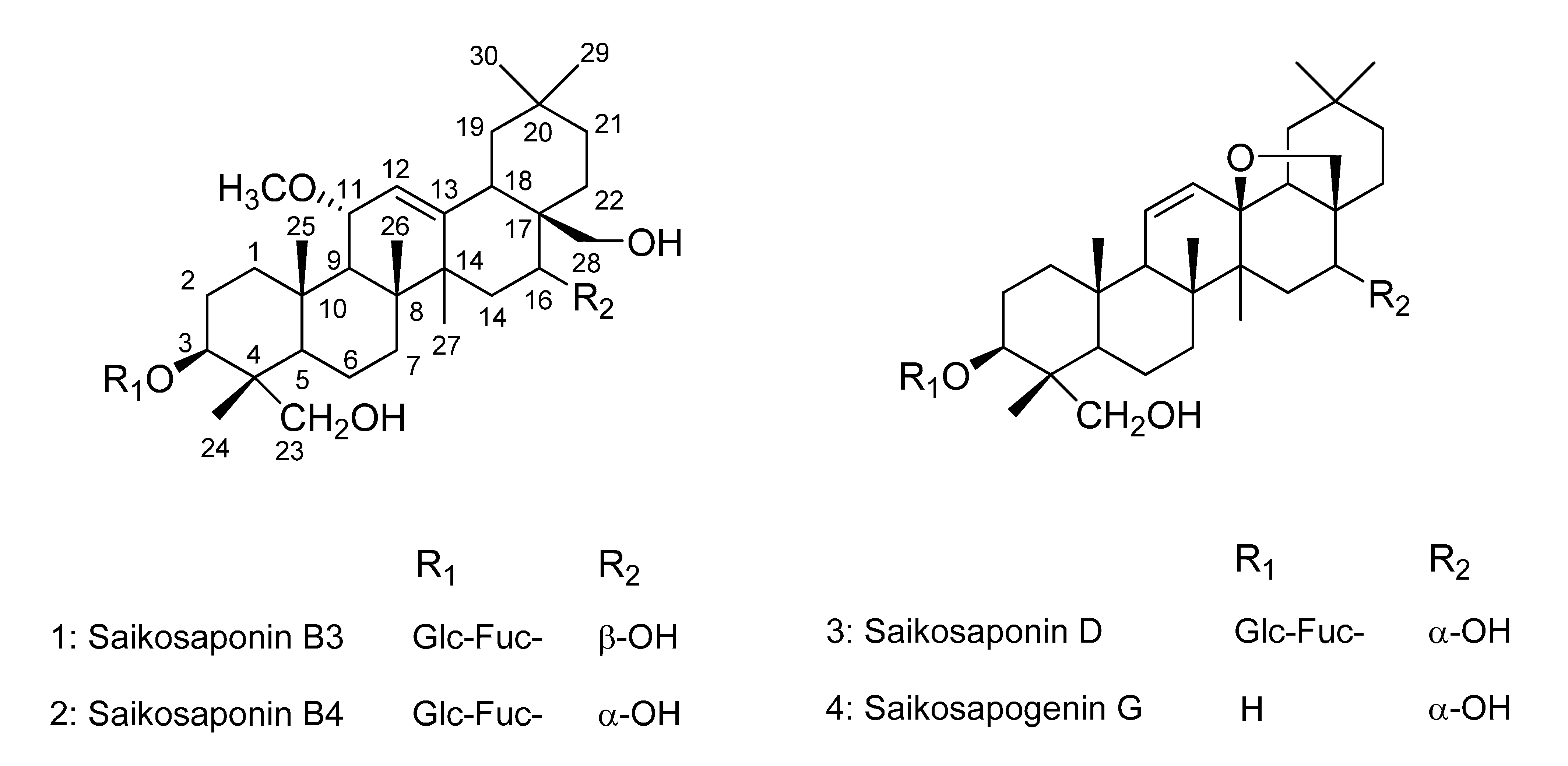
2.1. Isolation and Identification of Compounds 1–3

2.2. Cell Adhesion Inhibitory Activities
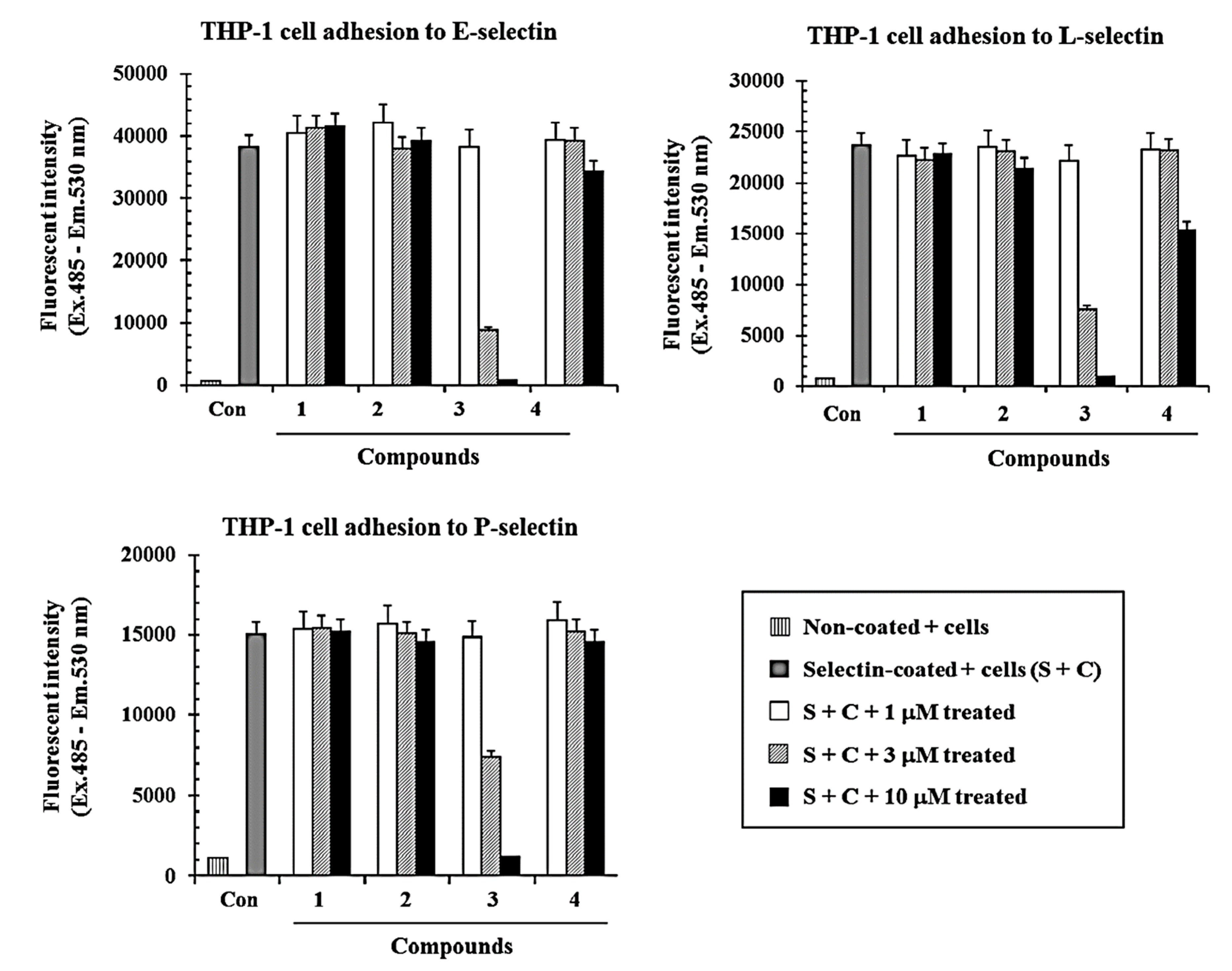
2.2.1. Inhibitory Effects of Compounds 1–4 on Selectins (E, L, and P)-Mediated Cell Adhesion Activities in Vitro


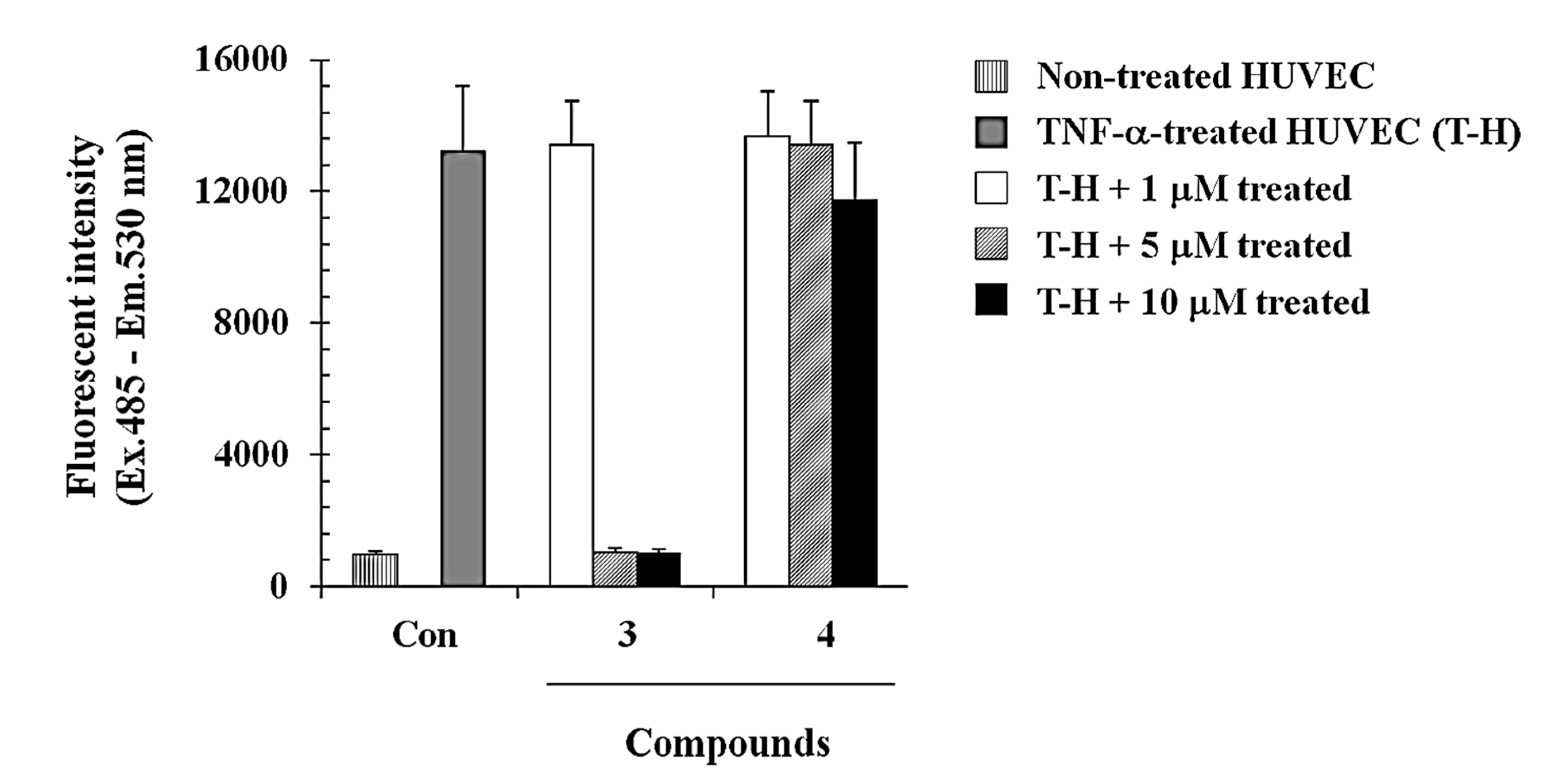
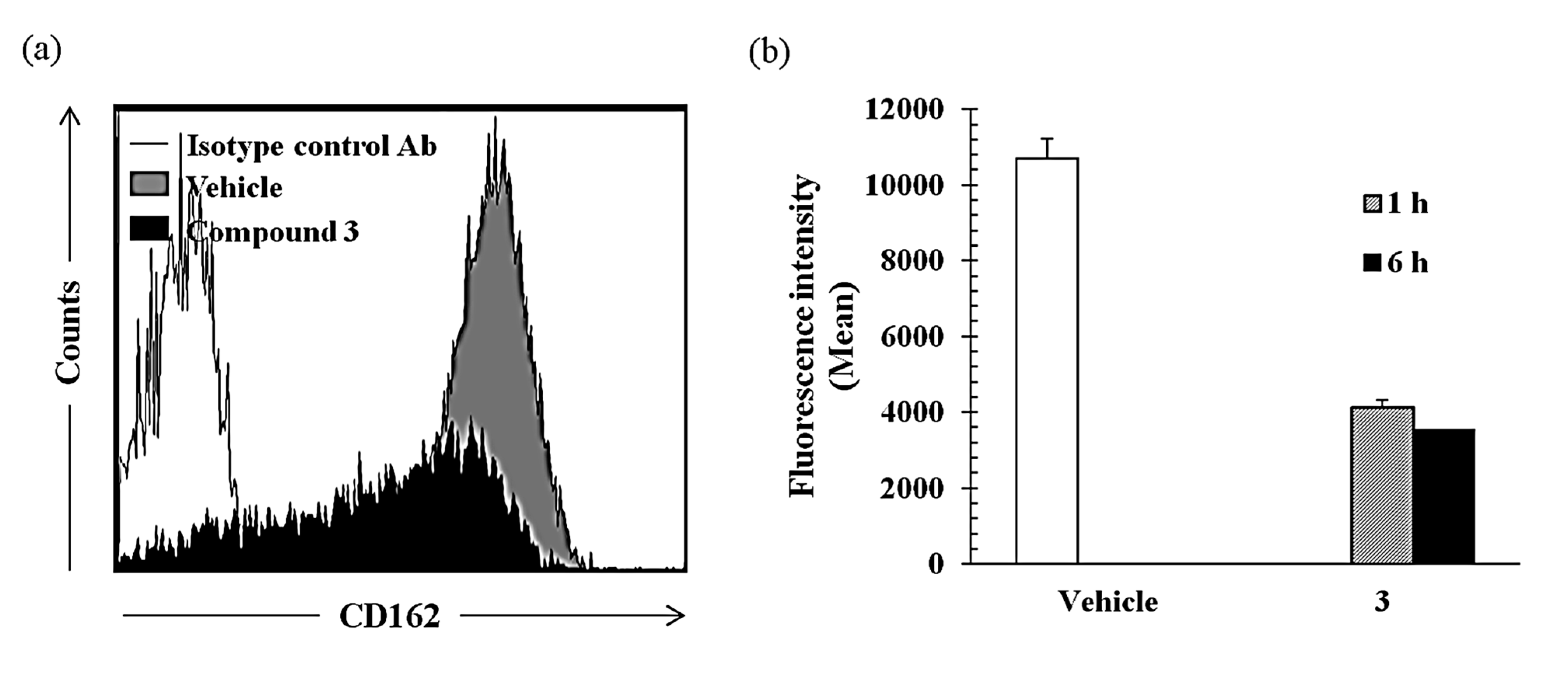
2.2.2. Compound 3 Inhibits Monocyte Adhesion onto Endothelial Cells Activated by TNF-α
2.2.3. Compound 3 Inhibits TNF-α-Induced Cell Adhesion Molecule Expression in Monocytes


3. Experimental Section
3.1. Plant Material, Extraction and Isolation
3.2. Cell Adhesion to E-Selectin, L-Selectin, and P-Selectin
3.3. Cell Adhesion to HUVEC
3.4. Cell Adhesion Molecule Expression on THP-1 Cells
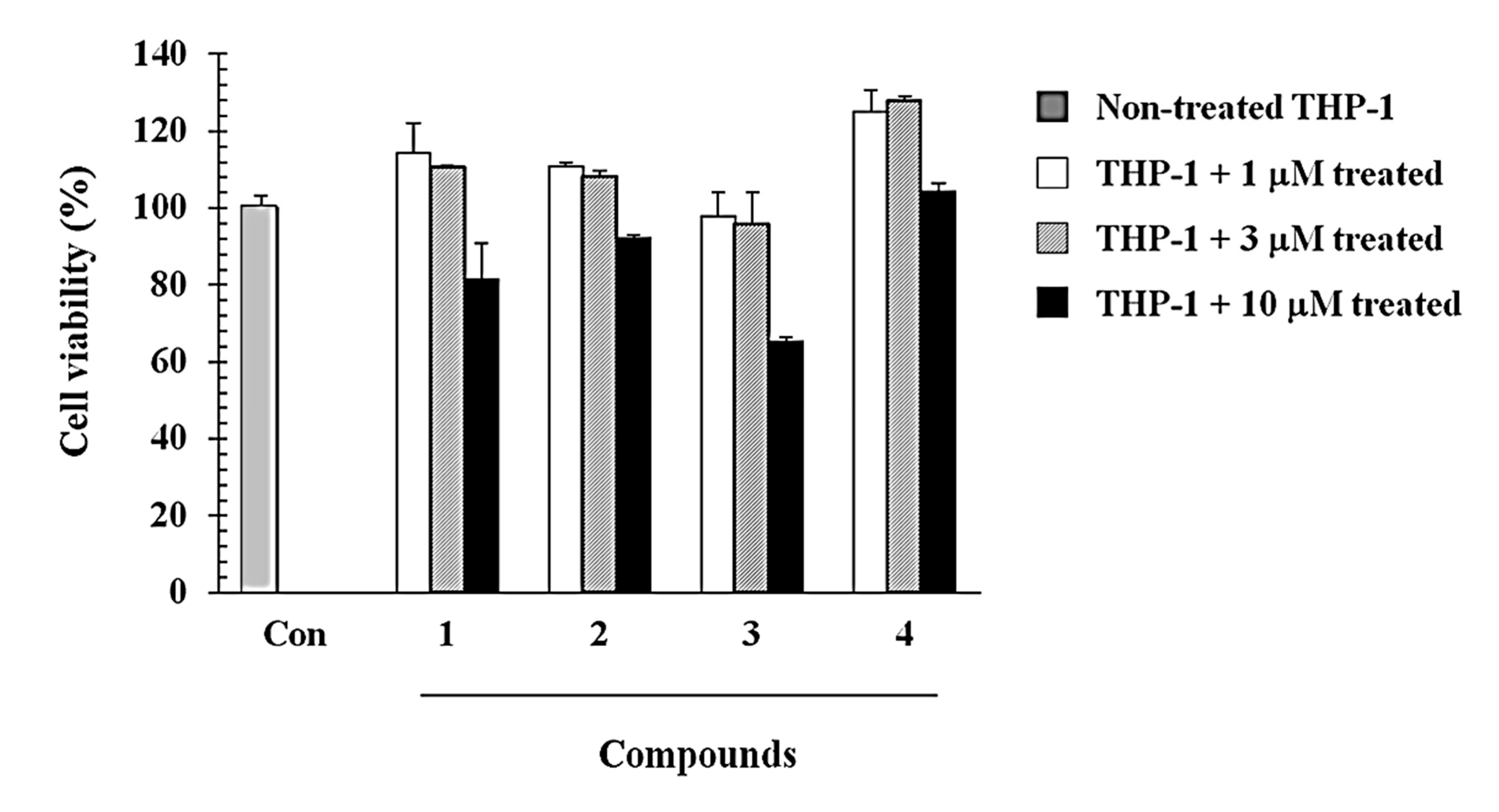
3.5. Cell Viability Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Springer, T.A. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell 1994, 76, 301–314. [Google Scholar]
- Vanderslice, P.; Biediger, R.J.; Woodside, D.G.; Berens, K.L.; Holland, G.W.; Dixon, R.A. Development of cell adhesion molecule antagonists as therapeutics for asthma and COPD. Pulm. Pharmacol. Ther. 2004, 17, 1–10. [Google Scholar]
- Auvinen, K.; Jalkanen, S.; Salmi, M. Expression and function of endothelial selectins during human development. Immunology 2014, 143, 406–415. [Google Scholar]
- Patel, K.D.; Cuvelier, S.L.; Wiehler, S. Selectins: Critical mediators of leukocyte recruitment. Semin. Immunol. 2002, 14, 73–81. [Google Scholar]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 3, 678–689. [Google Scholar]
- Kansas, G.S. Selectins and their ligands: Current concepts and controversies. Blood 1996, 88, 3259–3287. [Google Scholar]
- Ley, K. The role of selectins in inflammation and disease. Trends Mol. Med. 2003, 9, 263–268. [Google Scholar]
- Arbones, M.L.; Ord, D.C.; Ley, K.; Ratech, H.; Maynard-Curry, C.; Otten, G.; Capon, D.J.; Tedder, T.F. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity 1994, 1, 247–260. [Google Scholar]
- Yamada, H.; Sun, X.B.; Matsumoto, T.; Ra, K.S.; Hirano, M.; Kiyohara, H. Purification of anti-ulcer polysaccharides from the roots of Bupleurum falcatum. Planta Med. 1991, 57, 555–559. [Google Scholar]
- Park, K.H.; Park, J.; Koh, D.; Lim, Y. Effect of Saikosaponin-A, a Triterpenoid Glycoside, Isolated from Bupleurum falcatum on Experimental Allergic Asthma. Phytother. Res. 2002, 16, 359–363. [Google Scholar]
- Sun, Y.; Cai, T-.T.; Zhou, X-.B.; Xu, Q. Saikosaponin a inhibits the proliferation and activation of T cells through cell cycle arrest and induction of apoptosis. Int. Immunopharmacol. 2009, 9, 978–983. [Google Scholar]
- Leung, C.Y.; Liu, L.; Wong, R.N.S.; Zeng, Y.Y.; Li, M.; Zhou, H. Saikosaponin-d inhibits T cell activation through the modulation of PKCθ, JNK, and NF-κB transcription factor. Biochem. Biophys. Res. Commun. 2005, 338, 1920–1927. [Google Scholar]
- Lee, S.W.; Kim, M.S.; Lim, J.H.; Chang, J.S.; Ling, J.; Bae, K.H.; Lee, W.S.; Rho, M.C. Inhibition of VLA-4/VCAM-1-mediated Cell Adhesion by Triterpenoid Saponins from Bupleurum falcatum L. Bull. Korean Chem. Soc. 2010, 31, 1931–1936. [Google Scholar]
- Nose, M.; Amagaya, S.; Takeda, T.; Ogihara, Y. New derivatives of saikosaponin c. Chem. Pharm. Bull. 1989, 37, 1293–1296. [Google Scholar]
- Ishii, H.; Nakamura, M.; Seo, S.; Tori, K.; Tozyo, T.; Yoshimura, Y. Isolation, characterization, and nuclear magnetic resonance spectra of new saponins from the roots of Bupleurum falcatum L. Chem. Pharm. Bull. 1980, 28, 2367–2383. [Google Scholar]
- Tori, K.; Seo, S.; Yoshimura, Y.; Nakamura, M.; Yutaka, T.; Ishii, H. Carbon-13 NMR spectra of saikosaponin A, C, D and F isolated from Bupleurum falcatum L. Tetrahedron Lett. 1976, 46, 4167–4170. [Google Scholar]
- Kuboata, T.; Hinoh, H. The constitution of saponins isolated from Bupleurum falcatum L. Tetrahedron Lett. 1967, 9, 303–306. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, M.-J.; Kim, Y.S.; Bae, E.Y.; Oh, T.-S.; Choi, H.-J.; Lee, J.-H.; Oh, H.-M.; Lee, S.W. Saikosaponin D Isolated from Bupleurum falcatum Inhibits Selectin-Mediated Cell Adhesion. Molecules 2014, 19, 20340-20349. https://doi.org/10.3390/molecules191220340
Jang M-J, Kim YS, Bae EY, Oh T-S, Choi H-J, Lee J-H, Oh H-M, Lee SW. Saikosaponin D Isolated from Bupleurum falcatum Inhibits Selectin-Mediated Cell Adhesion. Molecules. 2014; 19(12):20340-20349. https://doi.org/10.3390/molecules191220340
Chicago/Turabian StyleJang, Myoung-Jun, Ye Sol Kim, Eun Young Bae, Tae-Seok Oh, Hwa-Jung Choi, Jung-Hee Lee, Hyun-Mee Oh, and Seung Woong Lee. 2014. "Saikosaponin D Isolated from Bupleurum falcatum Inhibits Selectin-Mediated Cell Adhesion" Molecules 19, no. 12: 20340-20349. https://doi.org/10.3390/molecules191220340