Griffipavixanthone from Garcinia oblongifolia Champ Induces Cell Apoptosis in Human Non-Small-Cell Lung Cancer H520 Cells in Vitro

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Anti-Proliferative Effect of GPX on NSCLC Cells

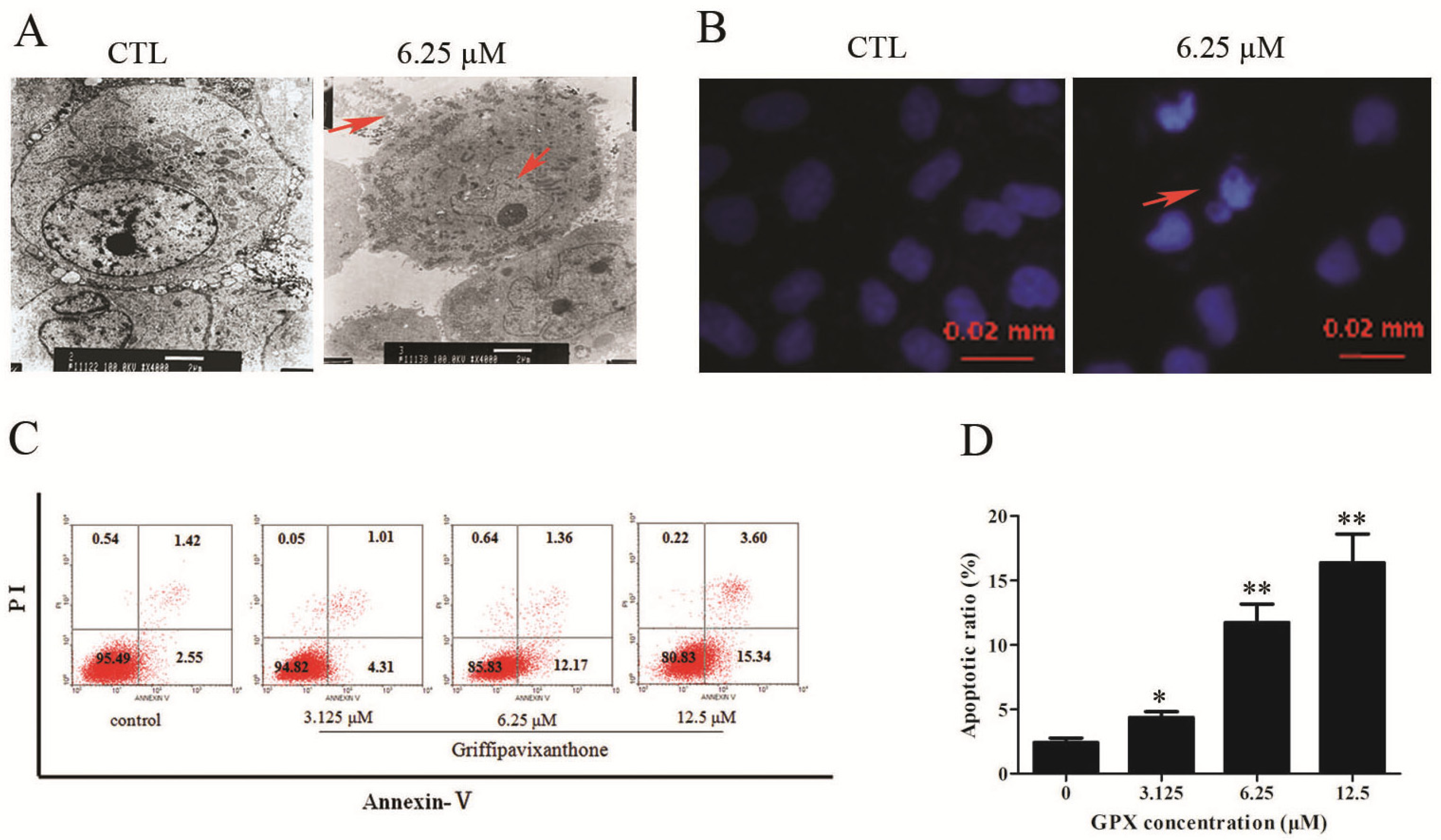
2.2. GPX Induced Apoptosis on H520 Cells
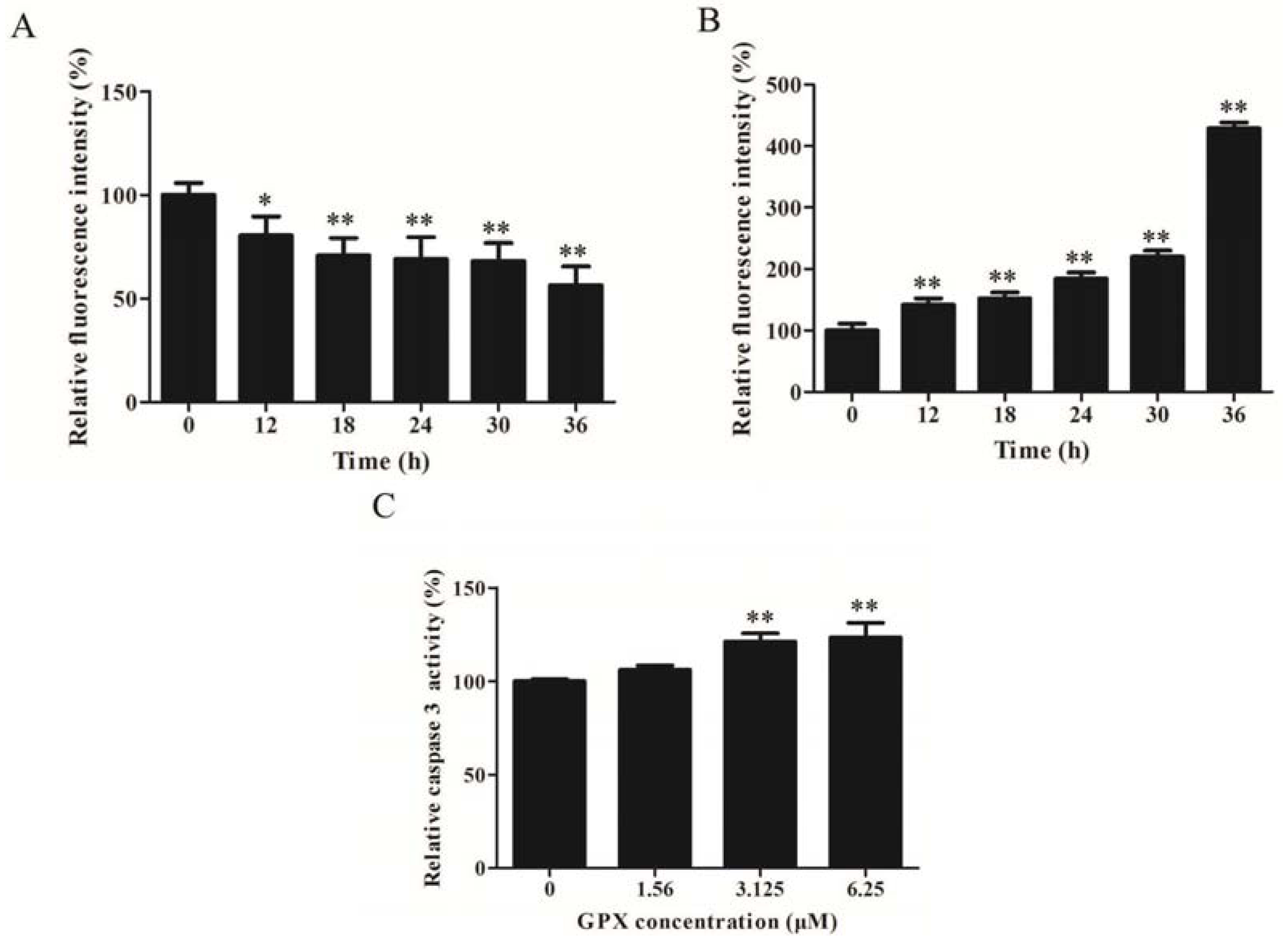
2.3. GPX Induced Apoptosis via Mitochondrial Apoptotic Pathway


3. Discussion
4. Experimental
4.1. Reagents and Chemicals
4.2. Plant Materials
4.3. Extraction and Isolation
4.4. Cell Culture
4.5. MTT Assay
4.6. NRU Assay
4.7. Hoechst 33,258 Staining
4.8. Transmission Electron Microscope Analysis
4.9. Cell Apoptosis Analysis
4.10. Mitochondrial Membrane Potential (ΔΨm) Assay and Intracellular ROS Assay
4.11. Caspase-3 Activity Assay
4.12. Statistical Analysis of Biological Assays
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Tran, Q.N. A novel method for finding non-small cell lung cancer diagnosis biomarkers. BMC Med. Genomics 2013, 6, 11–20. [Google Scholar] [CrossRef]
- Meoni, G.; Cecere, F.L.; Lucherini, E.; di Costanzo, F. Medical treatment of advanced non-small cell lung cancer in elderly patients: A review of the role of chemotherapy and targeted agents. J. Geriatr. Oncol. 2013, 4, 282–290. [Google Scholar] [CrossRef]
- Lynch, T.J.; Bell, D.W.; Sordella, R.; Gurubhagavatula, S.; Okimoto, R.A.; Brannigan, B.W.; Harris, P.L.; Haserlat, S.M.; Supko, J.G.; Haluska, F.G.; et al. Activating mutations in the epidermal growth factor receptor underlying responsiveness of non-small-cell lung cancer to gefitinib. N. Engl. J. Med. 2004, 350, 2129–2139. [Google Scholar] [CrossRef]
- Mayo, C.; Bertran-Alamillo, J.; Molina-Vila, M.Á.; Giménez-Capitán, A.; Costa, C.; Rosell, R. Pharmacogenetics of EGFR in lung cancer: Perspectives and clinical applications. Pharmacogenomics 2012, 13, 789–802. [Google Scholar] [CrossRef]
- Nakata, A.; Gotoh, N. Recent understanding of the molecular mechanisms for the efficacy and resistance of EGF receptor-specific tyrosine kinase inhibitors in non-small cell lung cancer. Expert Opin. Ther. Targets 2012, 16, 771–781. [Google Scholar] [CrossRef]
- Galvani, E.; Alfieri, R.; Giovannetti, E.; Cavazzoni, A.; La Monica, S.; Galetti, M.; Fumarola, C.; Bonelli, M.; Mor, M.; Tiseo, M.; et al. Epidermal growth factor receptor tyrosine kinase inhibitors: Current status and future perspectives in the development of novel irreversible inhibitors for the treatment of mutant non-small cell lung cancer. Curr. Pharm. Des. 2013, 19, 818–832. [Google Scholar] [CrossRef]
- Ayoola, A.; Barochia, A.; Belani, K.; Belani, C.P. Primary and acquired resistance to epidermal growth factor receptor tyrosine kinase inhibitors in non-small cell lung cancer: An update. Cancer Invest. 2012, 30, 433–446. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar]
- Xu, Y.J.; Cao, S.G.; Wu, X.H.; Lai, Y.H.; Tan, B.H.K.; Pereira, J.T.; Goh, S.H.; Venkatraman, G.; Harrison, L.J.; Sim, K.Y. Griffipavixanthone, a novel cytotoxic bixanthone from Garcinia griffithii and G. pavifolia. Tetrahedron Lett. 1998, 39, 9103–9106. [Google Scholar]
- Feng, S.; Jiang, Y.; Li, J.; Qiu, S.; Chen, T. A new bixanthone derivative from the bark of Garcinia oblongifolia. Nat. Prod. Res. 2013. [Google Scholar] [CrossRef]
- Merza, J.; Aumond, M.C.; Rondeau, D.; Dumontet, V.; le Ray, A.M.; Séraphin, D.; Richomme, P. Prenylated xanthones and tocotrienols from Garcinia virgata. Phytochemistry 2004, 65, 2915–2920. [Google Scholar] [CrossRef]
- Owa, T.; Yoshino, H.; Yoshimatsu, K.; Nagasu, T. Cell cycle regulation in the G1 phase: A promising target for the development of new chemotherapeutic anticancer agents. Curr. Med. Chem. 2001, 8, 1487–1503. [Google Scholar] [CrossRef]
- Shan, T.; Ma, Q.; Guo, K.; Liu, J.; Li, W.; Wang, F.; Wu, E. Xanthones from mangosteen extracts as natural chemopreventive agents: Potential anticancer drugs. Curr. Mol. Med. 2011, 11, 666–677. [Google Scholar] [CrossRef]
- Aisha, A.F.; Abu-Salah, K.M.; Ismail, Z.; Majid, A.M. In vitro and in vivo anti-colon cancer effects of Garcinia mangostana xanthones extract. BMC Complement. Altern. Med. 2012, 20, 104–114. [Google Scholar]
- Liu, Z.; Antalek, M.; Nguyen, L.; Li, X.; Tian, X.; Le, A.; Zi, X. The effect of gartanin, a naturally occurring xanthone in mangosteen juice, on the mTOR pathway, autophagy, apoptosis, and the growth of human urinary bladder cancer cell lines. Nutr. Cancer 2013, 65, 68–77. [Google Scholar] [CrossRef]
- Johnson, J.J.; Petiwala, S.M.; Syed, D.N.; Rasmussen, J.T.; Adhami, V.M.; Siddiqui, I.A.; Kohl, A.M.; Mukhtar, H. α-Mangostin, a xanthone from mangosteen fruit, promotes cell cycle arrest in prostate cancer and decreases xenograft tumor growth. Carcinogenesis 2012, 33, 413–419. [Google Scholar] [CrossRef]
- Mizushina, Y.; Kuriyama, I.; Nakahara, T.; Kawashima, Y.; Yoshida, H. Inhibitory effects of α-mangostin on mammalian DNA polymerase, topoisomerase, and human cancer cell proliferation. Food Chem. Toxicol. 2013, 59, 793–800. [Google Scholar] [CrossRef]
- Pontius, A.; Krick, A.; Mesry, R.; Kehraus, S.; Foegen, S.E.; Müller, M.; Klimo, K.; Gerhäuser, C.; König, G.M. Monodictyochromes A and B, dimeric xanthone derivatives from the marine algicolous fungus Monodictys putredinis. J. Nat. Prod. 2008, 71, 1793–1799. [Google Scholar] [CrossRef]
- Ondeyka, J.G.; Dombrowski, A.W.; Polishook, J.P.; Felcetto, T.; Shoop, W.L.; Guan, Z.; Singh, S.B. Isolation and insecticidal/anthelmintic activity of xanthonol, a novel bis-xanthone, from a non-sporulating fungal species. J. Antibiot. 2006, 59, 288–292. [Google Scholar] [CrossRef]
- Isaka, M.; Jaturapat, A.; Rukseree, K.; Danwisetkanjana, K.; Tanticharoen, M.; Thebtaranonth, Y. Phomoxanthones A and B, novel xanthone dimers from the endophytic fungus Phomopsis species. J. Nat. Prod. 2001, 64, 1015–1018. [Google Scholar] [CrossRef]
- Kagawa, S.; Gu, J.; Honda, T.; McDonnell, T.J.; Swisher, S.G.; Roth, J.A.; Fang, B. Deficiency of caspase-3 in MCF7 cells blocks Bax-mediated nuclear fragmentation but not cell death. Clin. Cancer Res. 2001, 7, 1474–1480. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shi, J.-M.; Huang, H.-J.; Qiu, S.-X.; Feng, S.-X.; Li, X.-E. Griffipavixanthone from Garcinia oblongifolia Champ Induces Cell Apoptosis in Human Non-Small-Cell Lung Cancer H520 Cells in Vitro. Molecules 2014, 19, 1422-1431. https://doi.org/10.3390/molecules19021422
Shi J-M, Huang H-J, Qiu S-X, Feng S-X, Li X-E. Griffipavixanthone from Garcinia oblongifolia Champ Induces Cell Apoptosis in Human Non-Small-Cell Lung Cancer H520 Cells in Vitro. Molecules. 2014; 19(2):1422-1431. https://doi.org/10.3390/molecules19021422
Chicago/Turabian StyleShi, Jun-Min, Hui-Juan Huang, Sheng-Xiang Qiu, Shi-Xiu Feng, and Xu-E Li. 2014. "Griffipavixanthone from Garcinia oblongifolia Champ Induces Cell Apoptosis in Human Non-Small-Cell Lung Cancer H520 Cells in Vitro" Molecules 19, no. 2: 1422-1431. https://doi.org/10.3390/molecules19021422