It has been widely reported that polyphenols exhibit a diverse range of beneficial biological effects, and as such, their consumption as part of the daily diet cannot be ignored. Although teas and herbal infusions contribute to the daily intake of dietary flavonoids, the pharmacological actions of these extracts cannot directly be attributed to single compounds within these beverages, unless identified and assayed in isolation. Flavonoids modulate steroid hormone biosynthesis and metabolism due to their interaction with steroidogenic enzymes, either inhibiting or stimulating specific enzymes [
7,
8]. We recently showed that unfermented rooibos extract influenced the steroid flux in the mineralocorticoid, glucocorticoid and androgen precursor pathways in human adrenal H295R cells under both basal and forskolin stimulated conditions [
8]. While the effect of flavonoid compounds on the flux through the adrenal steroidogenic pathways can be determined, the influence on specific enzymes in these pathways is less obvious due to upstream inhibition of steroid intermediates. In the present study, we assayed the effect of flavonoid compounds on individual steroidogenic enzymes as well as on steroid metabolite levels in H295R cells using ultra performance liquid chromatography/tandem mass spectrometry (UPLC-MS/MS).
2.1. The Influence of Rooibos and Selected Flavonoid Compounds on Adrenal Steroidogenic Enzymes Expressed in Non-Steroidogenic COS-1 Cells
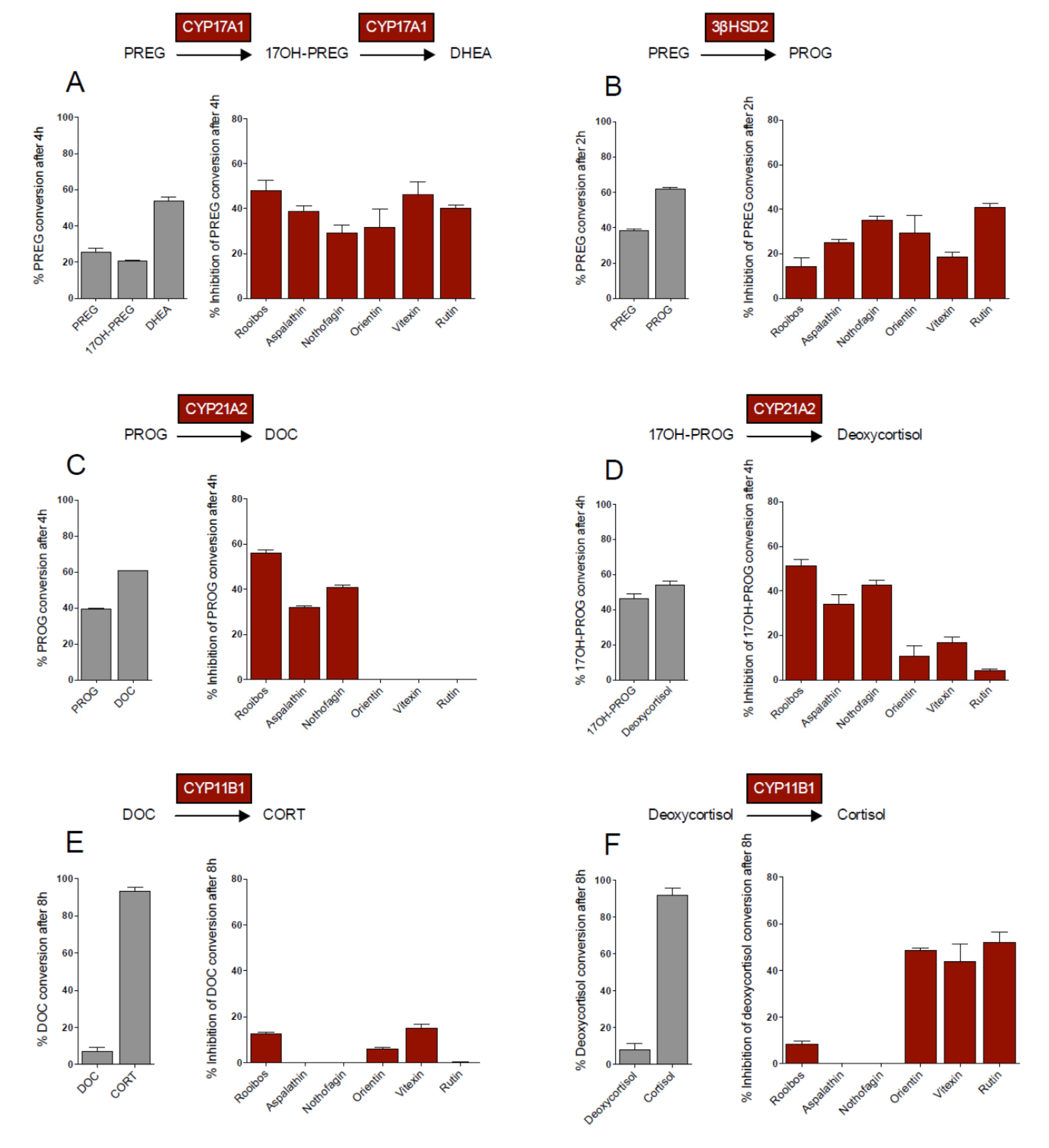
CYP17A1 plays a central role at the branch point in adrenal steroid biosynthesis, and together with 3βHSD2, determines the shunt of steroid metabolites in the glucocorticoid, mineralocorticoid and adrenal androgen pathways (
Scheme 1).
Scheme 1.
Steroid hormones produced in human adrenal cells.
Scheme 1.
Steroid hormones produced in human adrenal cells.
CYP17A1 catalyses the 17α-hydroxylation of pregnenolone (PREG), yielding 17OH-pregnenolone (17OH-PREG), which is in turn converted to dehydroepiandrosterone (DHEA). These three metabolites are substrates for 3βHSD2, which catalyzes their conversion to progesterone (PROG), 17OH-progesterone (17OH-PROG), and androstenedione (A4), respectively, thereby shunting metabolites into the respective pathways. In addition, CYP17A1 also catalyses the conversion of PROG to 17OH-PROG and 16OH-progesterone (16OH-PROG), with the latter not being further metabolized in the adrenal. PREG conversion by CYP17A1 was inhibited by all the flavonoid compounds assayed, with the compounds exhibiting similar inhibitory effects (
Figure 1A). Although PREG conversion by 3βHSD2 was also inhibited by all the flavonoids assayed, rutin exhibited the greatest inhibitory effect, while vitexin had the lowest inhibitory effect, 2-fold lower than that of rutin (
Figure 1B).
Figure 1.
Substrate conversion (1 µM) in transiently transfected COS-1 cells in the absence and presence of rooibos extract (4.3 mg/mL) and selected flavonoids (10 µM). CYP17A1 and CYP21A2 data adapted from [
8].
Figure 1.
Substrate conversion (1 µM) in transiently transfected COS-1 cells in the absence and presence of rooibos extract (4.3 mg/mL) and selected flavonoids (10 µM). CYP17A1 and CYP21A2 data adapted from [
8].
CYP21A2 catalyzes the conversion of PROG and 17OH-PROG to DOC and deoxycortisol, respectively. In contrast to CYP17A1 and 3βHSD2, only the dihydrochalcones inhibited the activity of CYP21A2 significantly, with the flavones and rutin not influencing the conversion of PROG by CYP21A2 (
Figure 1C). In the conversion of 17OH-PROG, however, these compounds displayed inhibitory effects below 20% (
Figure 1D). Neither aspalathin nor nothofagin inhibited the conversion of DOC and deoxycortisol by CYP11B1 to their corresponding products, CORT and cortisol, respectively. The main structural difference between the dihydrochalcones and the other flavonoids assayed in this study is the structural flexibility of the molecules and the glucose moiety at C3' on ring A of the dihydrochalcones, which could interfere with the binding of the compounds to CYP11B1. Inhibition of CYP11B1 by vitexin, orientin and rutin, however, was substrate specific. While inhibition of DOC conversion was negligible (
Figure 1E), inhibition of deoxycortisol conversion was significantly higher in the presence of rutin, orientin and vitexin (
Figure 1F). The higher binding affinity of CYP11B1 for DOC compared to deoxycortisol may account for the negligible inhibition of the flavonoids on the conversion of DOC. CYP11B1 has been shown to exhibit a lower Km for DOC and its catalytic conversion is also characterized by a lower K
cat, 2-fold less than that of deoxycortisol conversion, clearly indicating that a single hydroxyl group influences the enzyme’s affinity for its substrate, as well as the substrate turnover. It was suggested that the presence of the C17 hydroxyl group of deoxycortisol obstructs the entrance of deoxycortisol into the enzyme’s active pocket [
12].
While the rooibos extract exhibited the greatest inhibitory effect on CYP17A1 and CYP21A2, inhibition of the other enzymes was negligible. The inhibition of PREG conversion by CYP17A1 (
Figure 1A) as well as the conversion of PROG (
Figure 1C) and 17OH-PROG (
Figure 1D) by CYP21A2 was similar (±50%) in the presence of the extract. However, the inhibitory effect of rooibos on PREG conversion by 3βHSD2 (
Figure 1B) and DOC (
Figure 1E) and deoxycortisol (
Figure 1F) conversion by CYP11B1, was less than 20%. The inhibitory effects brought about by the individual flavonoid compounds (10 µM) was not reflected in the inhibitory effects of the whole extract in all instances, even though the flavonoid concentrations within the rooibos extract ranged between 27 µM and 1.4 mM. Although we assayed five of the major flavonoids present in rooibos, it is important to note that, to date, 46 flavonoid compounds have been identified in rooibos [
13,
14] which may possibly contribute to the data obtained in the presence of the extract. From the data it is clear, however, that the rooibos extract preferentially inhibits CYP17A1 and CYP21A2, two key enzymes at the branch point of adrenal steroidogenesis.
2.2. The Influence of Selected Flavonoids on Steroid Metabolism in Adrenal H295R Cells under Basal and Forskolin Stimulated Conditions
In H295R cells, the dihydrochalcones and the flavones decreased the total steroid output under both basal and stimulated conditions, while rutin’s effect was only detected in the presence of forskolin. Forskolin mimics the effects of ACTH by activating cAMP pathways in adrenal cells [
15], stimulating steroidogenic enzymes and increasing steroid production. As expected, forskolin significantly increased the total steroid output to 11.1 µM (2.8 fold), with the greatest inhibition being detected in the presence of rutin (
Table 2). Both aspalathin and nothofagin significantly reduced basal 17OH-PROG and 16OH-PROG levels due to the inhibition of CYP17A1 and 3βHSD2. Nothofagin also reduced deoxycortisol levels under basal conditions, possibly due to inhibition of CYP21A2 or due to lower levels of 17OH-PROG being available as the precursor substrate. Both dihydrochalcones reduced A4 levels significantly under basal and forskolin stimulated conditions, indicative of 3βHSD2 inhibition. Interestingly, while nothofagin reduced basal levels of 11β-hydroxyandrostenedione (11OHA4), the product of A4, it did not reduce basal production of cortisol, which is catalysed by the same enzyme, CYP11B1. Although this may suggest possible inhibition of CYP11B1, neither aspalathin nor nothofagin were shown to inhibit the enzyme at 10 µM in COS-1 cells, suggesting that the reduction in 11OHA4 was due to upstream inhibition. Under stimulated conditions, the production of both cortisol and 11OHA4 was inhibited in the presence of aspalathin and nothofagin, indicating upstream inhibition, resulting in reduced levels of precursor steroid metabolites. It should, however, also be noted that when comparing inhibitory effects of the flavonoid compounds under basal and stimulated conditions, the expression levels of steroidogenic enzymes are altered when cells are stimulated with forskolin, which would in turn alter steroid metabolite/precursor levels. From these results it is clear that the inhibitory effects of both dihydrochalcones are very similar, however, the inhibitory effect of nothofagin was notably greater than that of aspalathin.
Table 2.
Steroid metabolites produced in adrenal H295R cells under basal and forskolin (10 µM) stimulated conditions in the absence and presence of selected flavonoids (10 µM) after 48 h.
Table 2.
Steroid metabolites produced in adrenal H295R cells under basal and forskolin (10 µM) stimulated conditions in the absence and presence of selected flavonoids (10 µM) after 48 h.
| Steroid metabolites | Basal | + Aspalathin a | + Nothofagin a | + Orientin | + Vitexin | + Rutin b |
|---|
| Total ± SEM (nM) | Fold change | Fold change | Fold change | Fold change | Fold change |
|---|
| PREG | 35.2 | ± | 2.2 | | | | | | | | | | ↑ | 1.4 | ** | | | |
| PROG | 5.8 | ± | 1.7 | | | | | | | | | | | | | | | |
| DOC | 107.4 | ± | 13.0 | | | | | | | ↑ | 1.2 | * | ↑ | 1.2 | ** | | | |
| CORT | 241.1 | ± | 31.6 | | | | | | | ↑ | 1.2 | * | | | | | | |
| 18OH-CORT | 9.3 | ± | 1.3 | | | | | | | ↑ | 2.6 | * | | | | | | |
| ALDO | 2.3 | ± | 0.5 | | | | | | | | | | | | | | | |
| 11-DHC | 9.9 | ± | 0.3 | | | | | | | | | | | | | | | |
| 16OH-PROG | 66.4 | ± | 10.3 | ↓ | 2.3 | *** | ↓ | 3.6 | *** | ↓ | 1.2 | *** | ↓ | 1.8 | *** | | | |
| 17OH-PROG | 56.8 | ± | 6.3 | ↓ | 3.6 | ** | ↓ | 4.8 | ** | | | | ↓ | 1.4 | * | | | |
| Deoxycortisol | 1741.0 | ± | 234.1 | | | | ↓ | 2.3 | * | | | | ↓ | 1.3 | *** | | | |
| Cortisol | 670.2 | ± | 39.3 | | | | | | | | | | ↓ | 1.3 | ** | | | |
| Cortisone | 6.6 | ± | 2.2 | | | | | | | ↓ | 1.5 | *** | ↓ | 1.8 | *** | | | |
| A4 | 806.5 | ± | 115.6 | ↓ | 2.4 | ** | ↓ | 3.5 | *** | ↓ | 1.1 | * | ↓ | 1.5 | *** | | | |
| 11OHA4 | 90.1 | ± | 11.1 | | | | ↓ | 3.0 | * | | | | ↓ | 1.5 | *** | | | |
| Testosterone | 39.22 | ± | 3.60 | ↓ | 4.3 | *** | ↓ | 7.1 | *** | ↓ | 1.3 | ** | ↓ | 2.5 | *** | | | |
| Total steroid (nM) | 3947.0 | ↓ | 1.6 | | ↓ | 2.0 | * | ↓ | 1.0 | | ↓ | 1.3 | ** | ↓ | 1.0 | |
| Steroid metabolites | Forskolin | + Aspalathin a | + Nothofagin a | + Orientin | + Vitexin | + Rutin b |
| Total ± SEM (nM) | Fold change | Fold change | Fold change | Fold change | Fold change |
| PREG | 47.5 | ± | 3.4 | | | | | | | ↑ | 1.4 | ** | ↑ | 1.3 | ** | | | |
| PROG | 5.1 | ± | 0.5 | | | | | | | ↑ | 1.4 | ** | ↑ | 1.5 | ** | | | |
| DOC | 343.2 | ± | 61.1 | | | | | | | ↓ | 1.2 | * | ↓ | 1.1 | * | | | |
| CORT | 2062.0 | ± | 170.5 | | | | | | | | | | ↓ | 1.3 | ** | ↓ | 1.9 | * |
| 18OH-CORT | 104.8 | ± | 15.1 | | | | | | | | | | | | | ↓ | 2.0 | ** |
| ALDO | 27.4 | ± | 3.4 | | | | | | | ↓ | 1.1 | *** | ↓ | 1.4 | * | | | |
| 11-DHC | 11.2 | ± | 1.4 | | | | | | | | | | | | | | | |
| 16OH-PROG | 66.0 | ± | 4.7 | | | | | | | | | | ↓ | 1.4 | ** | | | |
| 17OH-PROG | 36.0 | ± | 2.4 | | | | | | | | | | | | | ↓ | 3.1 | * |
| Deoxycortisol | 2757.0 | ± | 393.1 | | | | | | | | | | ↓ | 1.3 | * | ↓ | 2.0 | ** |
| Cortisol | 3793.0 | ± | 285.1 | ↓ | 1.3 | *** | ↓ | 1.7 | *** | ↓ | 1.3 | *** | ↓ | 1.8 | *** | ↓ | 3.0 | *** |
| Cortisone | 11.6 | ± | 2.7 | | | | ↓ | 2.6 | * | | | | | | | | | |
| A4 | 1402.0 | ± | 180.0 | ↓ | 1.7 | *** | ↓ | 2.0 | *** | ↓ | 1.4 | *** | ↓ | 1.5 | *** | ↓ | 4.4 | *** |
| 11OHA4 | 388.3 | ± | 16.3 | ↓ | 1.3 | ** | ↓ | 2.5 | *** | ↓ | 1.5 | *** | ↓ | 1.7 | *** | ↓ | 6.9 | *** |
| Testosterone | 50.5 | ± | 6.1 | ↓ | 1.7 | *** | ↓ | 2.4 | *** | ↓ | 1.7 | *** | ↓ | 2.3 | *** | ↓ | 5.8 | *** |
| Total steroid (nM) | 11,141.0 | ↓ | 1.4 | * | ↓ | 1.4 | * | ↓ | 1.1 | | ↓ | 1.3 | ** | ↓ | 2.3 | ** |
Orientin, the flavone analogue of aspalathin, significantly increased the steroid flux in the mineralocorticoid pathway, increasing basal levels of DOC, CORT and 18OH-CORT, suggesting that inhibition of CYP17A1 resulted in more metabolites being channeled into the mineralocorticoid pathway. The data obtained clearly shows that the flavones inhibit CYP17A1, since PREG and PROG levels increased, while 16OH-PROG and 17OH-PROG levels decreased (
Table 2).
The inhibitory profile of vitexin was very similar to that of orientin, however, vitexin exhibited a greater inhibitory effect, reducing the total steroid output 1.3-fold under both basal and stimulated conditions, while the effect of orientin on total steroid production was negligible. Interestingly, under stimulated conditions, during which aldosterone synthase (CYP11B2) expression is upregulated [
16], the effect of vitexin on the mineralocorticoid pathway was much more prominent compared to orientin, with a greater reduction in ALDO levels being observed, suggesting inhibition of either CYP11B2 or upstream enzymes. Under both basal and stimulated conditions, vitexin also had a much greater effect on the glucocorticoid pathway compared to orientin, reducing the levels of cortisol and its precursors significantly. In COS-1 cells, rutin as well as the flavones inhibited the biosynthesis of cortisol by CYP11B1, while the dihydrochalcones did not, suggesting that the reduced cortisol levels observed in H295R cells in the presence of aspalathin and nothofagin may be indicative of upstream inhibition only.
While the dihydrochalcones and their flavone analogues reduced basal testosterone levels significantly, it was the former compounds which had the greatest effect, suggesting inhibition of 17βHSD. However, the adrenal is not the primary site for 17βHSD expression, as is also the case for 11βHSD2, which catalyses the formation of cortisone and 11-dehydrocorticosterone (11-DHC). The detected levels of these metabolites were very low, with the flavones reducing cortisone levels under basal conditions, while under stimulated conditions, it was inhibited in the presence of nothofagin only. Although 17βHSD and 11βHSD2 are expressed at very low levels in the adrenal [
17], it would appear that these hydroxysteroid dehydrogenases, together with 3βHSD2, are sensitive to the flavonoid compounds present in Rooibos.
In a recent pharmacophore-based virtual screening study investigating inhibitors of 17βHSD type 3 and 5, key enzymes involved in adrenal androgen production, it was shown that the ligand binding domain of these enzymes is able to accommodate structurally highly diverse ligands which bind to different regions of the active site. An enlargement of the binding cavity was observed in the crystal structure of 17βHSD3/5 when rutin was bound to these enzymes. Furthermore, the X-ray structure of rutin bound to 17βHSD5 showed that water molecules formed a hydrogen bonding network with rutin bound at the base of the ligand binding domain [
18].
Rutin, one of the more stable flavonoid compounds in both fermented and unfermented Rooibos, had no significant effect on any of the steroid hormones under basal conditions. However, upon forskolin stimulation, rutin had the greatest inhibitory effect of all the flavonoid compounds assayed, reducing the total steroid output 2.3-fold. In the mineralocorticoid pathway, CORT and 18OH-CORT levels were significantly lower, suggesting inhibition of CYP11B2. In the glucocorticoid pathway, significant reductions in the levels of 17OH-PROG, deoxycortisol and cortisol indicate inhibition of CYP17A1 and CYP11B1, as was also shown in COS-1 cells (
Figure 1). Upstream inhibition also contributed to the reduced deoxycortisol levels, since rutin did not inhibit CYP21A2. In the adrenal androgen precursor pathway, rutin reduced the levels of A4, testosterone and 11OHA4, suggesting inhibition of CYP17A1 and/or 3βHSD2.
The data obtained in the H295R cell line did not in all cases reflect results obtained in COS-1 cells. The expression levels of the individual enzymes assayed in COS-1 cells are comparable, as the cells were transiently transfected with equal concentrations of the appropriate cDNA. Furthermore, the conversion of substrates in COS-1 cells allowed assays to be conducted under very specific conditions. A fixed substrate concentration and no competition from other enzymes for the same substrates, as is the case in the adrenal H295R cell model, allowed us to investigate the effect of flavonoids on the catalytic activity of specific steroidogenic enzymes. In contrast, enzymes are expressed at different levels in adrenal H295R cells and vary from enzyme to enzyme under both basal and forskolin stimulated conditions, with specific enzymes being upregulated when cells are stimulated. It was recently shown that under basal conditions in H295R cells, CYP11A1 is expressed at the highest levels, followed by CYP21A2, CYP17A1, SULT2A1, 3βHSD2, CYP11B2 and CYP11B1, while under forskolin stimulated conditions, CYP17A1 is expressed at the highest levels, followed by CYP21A2, CYP11A1, 3βHSD2, SULT2A1, CYP11B2, and CYP11B1 [
16].
Overall, from these results it is clear that orientin and vitexin had the greatest effect on the mineralocorticoid pathway under both basal and forskolin stimulated conditions. The dihydrochalcones had no significant effect on any of the metabolites in the mineralocorticoid pathway, while rutin’s effect was only detected under stimulated conditions, with significantly reduced levels of ALDO precursors, CORT and 18OH-CORT. The influence of rutin was most evident in the glucocorticoid and adrenal androgen pathways under stimulated conditions, with the other flavonoids decreasing metabolites under both basal and stimulated conditions in these pathways.
2.3. Structure Activity Analyses
Analyses regarding the influence of structural differences of flavonoid compounds on their ability to inhibit steroidogenic enzymes are hampered due to the uncertainty of the manner in which these compounds would interact with the active site of these enzymes. As the type of inhibition of the steroidogenic cytochromes P450 by the flavonoids is not known, it is possible that binding could also occur at sites other than the active pocket. Furthermore, molecular docking studies reporting on structural aspects of flavonoid compounds impacting on substrate inhibition in terms of steroidogenic enzymes are limited. It has, however, been shown by Ohno
et al. that the isoflavone, diadzein, was able to competitively inhibit the binding of DHEA and PROG to 3βHSD2 and CYP21A2, respectively. The authors suggested, having assayed a range of flavone compounds, that it was the hydroxyl groups at C6 on ring A and C4' on ring B which played an important role in the inhibition of steroidogenic P450 enzymes. They showed, in cAMP-stimulated H295R cells, at a concentration of 12.5 µM, that 6-hydroxyflavone, 4'-hydroxyflavone and apigenin, which contains hydroxyl groups at C5, C7 and C4', inhibited cortisol production significantly [
7]. In the present study, vitexin, with a C4' hydroxyl group and orientin with C3' and C4' hydroxyl groups, both also containing a glucose moiety at C8, significantly inhibited cortisol biosynthesis at a concentration of 10 µM. In COS-1 cells, it was the flavones and rutin that inhibited CYP11B1 significantly, while the dihydrochalcones did not inhibit the activity of CYP11B1 towards DOC or deoxycortisol. It is interesting to note that the flavones as well as rutin, containing glucose moieties on ring A and ring C respectively, showed negligible inhibition of CYP21A2. In contrast, while aspalathin and nothofagin significantly inhibited substrate conversion by CYP21A2, these compounds had no influence on the conversion of either DOC or deoxycortisol by CYP11B1, possibly due to the glucosyl moiety at C3' on ring A. Our data thus far suggests that a glucosyl moiety at this position may prevent compounds inhibiting the catalytic activity of CYP11B1, while the C8 glucosyl and C3 rutinosyl moieties on the pyran ring of the other flavonoids do not. Substitutions at C3 and C4 (dihydrochalcones) and C3' and C4' (flavones and flavonol) on the B ring may, however, also play a role. The type of inhibition that these compounds elicit need to be determined, while homology models may pinpoint the effect of functional groups on the binding of the compounds in the active pocket of the relevant enzymes.
In a more recent study by Hasegawa
et al. [
5], it was also shown in H295R cells that a range of flavones with hydroxyl groups at positions C4' or C3' and C4' on ring B significantly reduced the levels of DOC and A4, suggesting inhibition of 3βHSD2, while apigenin was more potent than the other polyphenols assayed, increasing the levels of PREG and 17OH-PROG, suggesting inhibition of CYP17A1, CYP21A2 and 3βHSD2. Apigenin was also shown to downregulate the expression levels of CYP17A1, CYP21A2 and 3βHSD2 mRNA significantly [
5].
From our data it appears that the enzymes 3βHSD2 and CYP17A1 are most susceptible to inhibition by the rooibos flavonoids assayed in this study. The natural substrates for these two enzymes, PREG, 17OH-PREG and DHEA contains a hydroxyl group on C3 of the steroid backbone structure involved with substrate binding in the active pocket [
19,
20]. It is possible that the hydroxyl group on position C4' of the flavonoid is involved in the binding of these compounds to the active pocket of steroidogenic P450 enzymes, with hydrogen bonds stabilizing the orientation of flavonoid compounds in the active site. However, since limited data is available on the molecular docking of flavonoids, no conclusions can be drawn regarding the orientation of these compounds within the active pocket. In addition, as the mechanism of inhibition is uncertain, the binding of these compounds to the same site that the substrate would occupy cannot be assumed. A recent study by Androutsopoulos
et al. [
21], investigating the binding mode of selected flavonoids to the heme group using the CYP1A2 crystal structure as a template, predicted that flavonoids bind in the active pocket with the B-ring orientated towards the heme group. In an earlier study, Shimada
et al. [
22] investigated the inhibition of a range flavonoid compounds towards five human P450 enzymes by assessing inhibitory activity together with molecular docking studies. These studies clearly showed that the position and number of hydroxy and methoxy groups impacted on the orientation of the compounds in the active pocket of the different enzymes. It was shown that ring B of the compounds was not, in all cases, oriented towards the heme group and that hydroxy/methoxy substitutions influenced positioning of the flavonoid compounds and thus also affected the mechanisms of inhibition for the enzymes assayed.
The compounds assayed in this study all contained glucose moieties on either ring A or C, which would, if bound in the active site of the enzyme, affect the orientation of the molecule, impacting on the mechanism of inhibition. It would seem that while a hydroxy group on B ring plays a role in flavonoid inhibitory activity, a second hydroxy group does not significantly affect inhibition or contribute towards increasing inhibition, possibly due to rotation between rings A and B resulting in a degree of flexibility. It should, however, also be noted that aspalathin and nothofagin have a more flexible structure compared to rutin, orientin and vitexin. It is interesting to note that the dihydrochalcones inhibit the catalytic activity of CYP21A2 towards both substrates, while exhibiting no inhibitory effect on CYP11B1, regardless of the substrate. In contrast, the inhibition of the activity of these enzymes towards their substrates differs in the presence of flavone/flavonol compounds, with the inhibitory effect of these compounds being greater when the substrate is hydroxylated at the C17 position.


{kind=link}
{kind=link}




