Comparison of Physicochemical Properties and Immunomodulatory Activity of Polysaccharides from Fresh and Dried Litchi Pulp

Abstract
:1. Introduction
2. Results and Discussion
2.1. Preliminary Characterization of Litchi Polysaccharides
2.1.1. The Chemical Compositions of Litchi Polysaccharides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | LPF | LPD |
|---|---|---|
| Neutral sugar (W%) | 66.85 ± 1.12 | 65.73 ± 1.76 |
| Uronic acid (W%) | 2.09 ± 0.48 | 4.72 ± 0.27 * |
| Protein (W%) | 3.94 ± 0.16 | 6.48 ± 0.24 * |
| Molecular weight (Da) | 970085 | 370365; 8207 |
| Monosaccharide composition (%) | ||
| Rhamnose | 0.31 | 1.86 |
| Arabinose | 5.44 | 17.62 |
| Xylose | 0.71 | 3.21 |
| Mannose | 15.18 | 10.56 |
| Glucose | 66.1 | 20.82 |
| Galactose | 11.58 | 41.18 |
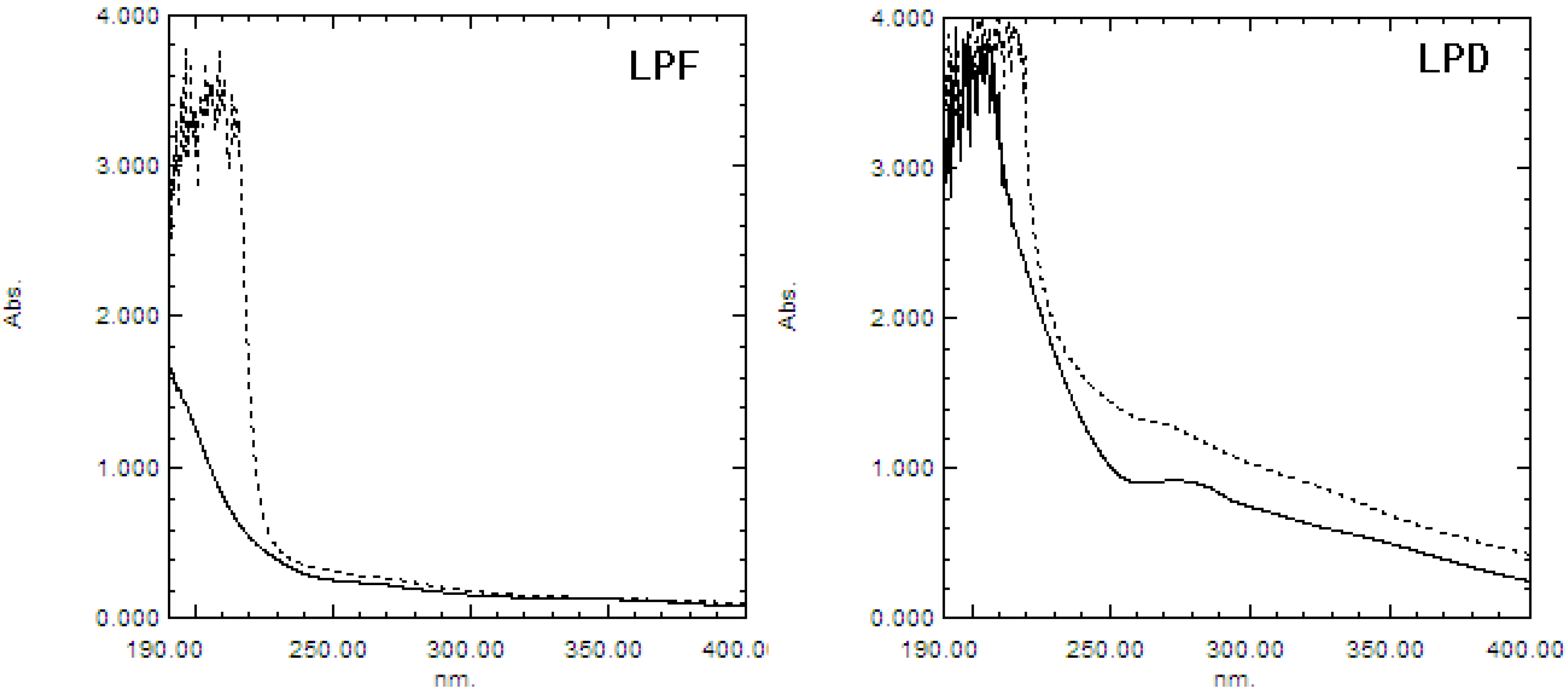
2.1.2. UV and IR Spectra Analysis

| Absorption (cm−1) a | Functional group b | Structural characteristics | |
|---|---|---|---|
| LPF | LPD | ||
| 3397.4 | 3405.6 | hydroxyl group (-OH) | O-H stretching vibration |
| amino group (-NH2) | N-H stretching vibration | ||
| 2928.0 | 2926.1 | alkyl group (-CH2-) | C-H stretching vibration |
| 1654.5 | 1637.6 | carbonyl group (-C=O or -CHO) | C=O stretching vibration |
| amide group (-NH2 or –COR) | N-H bending vibration or C=O stretching vibration | ||
| amino group (-NH2) | N-H bending vibration | ||
| bound water | |||
| 1541.4 | amino group (-NH2) or amide group (-NH2) | N-H bending vibration | |
| carbonyl group (-C=O) | C=O stretching vibration | ||
| 1458.8 | alkyl group (-CH2- or –CH3) | C-H bending vibration | |
| 1420.0 | 1420.8 | carboxyl group (-COOH) | C-O stretching vibration |
| 1364.0 | 1375.8 | carboxyl group (-COOH) | C=O symmetrical stretching vibration |
| 1270.8 | 1275.1 | carboxyl group (-COOH) | O-H bending vibration |
| 1156.0 | ether (-C-O-C-) | C-O stretching vibration | |
| 1017.0 | 1074.7 | hydroxyl group (-OH) | O-H bending vibration |
| 917.9 | D-glucopyranose ring | Antisymmetrical ring vibration | |
| 848.9 | α-type glycosidic linkage | C-H bending vibration | |
| 764.8 | 777.4 | D-glucopyranose ring | symmetrical ring vibration |
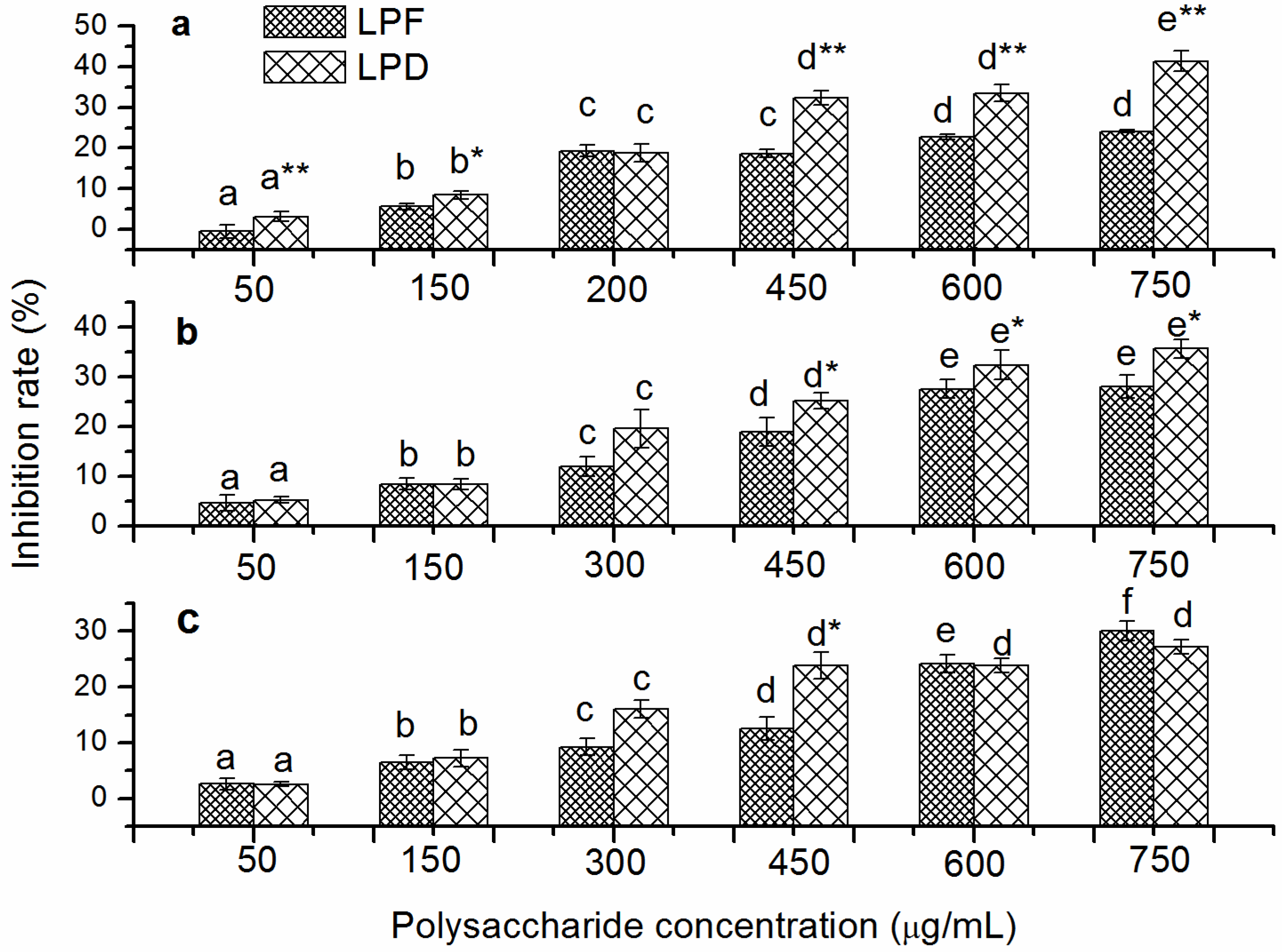
2.2. In Vitro Inhibition Effects of Tumor Cell Proliferation of Litchi Polysaccharides

2.3. In Vitro Immunostimulatory Activity of Litchi Polysaccharides
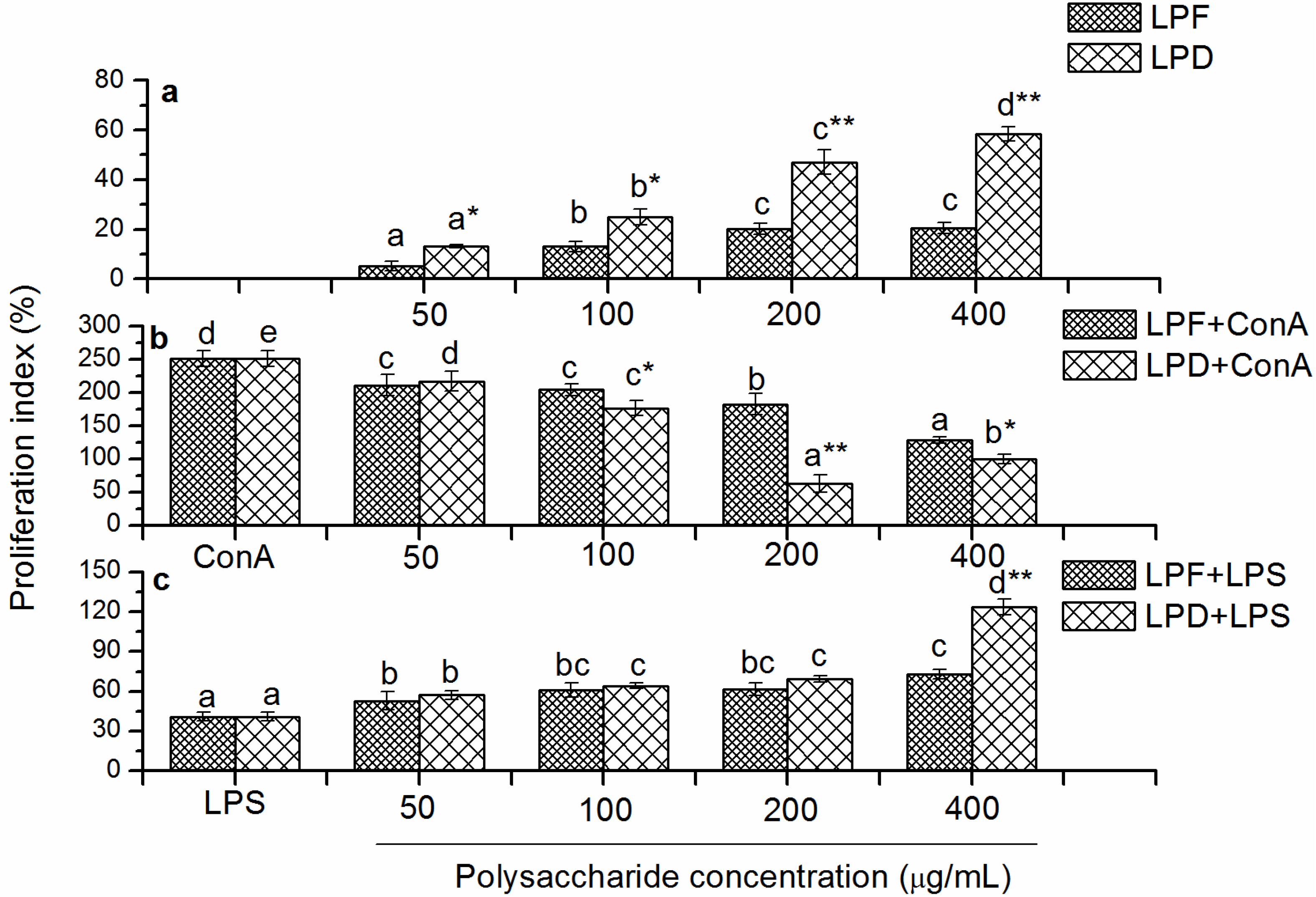
2.3.1. Effects of Litchi Polysaccharides on Splenocyte Proliferation in Vitro

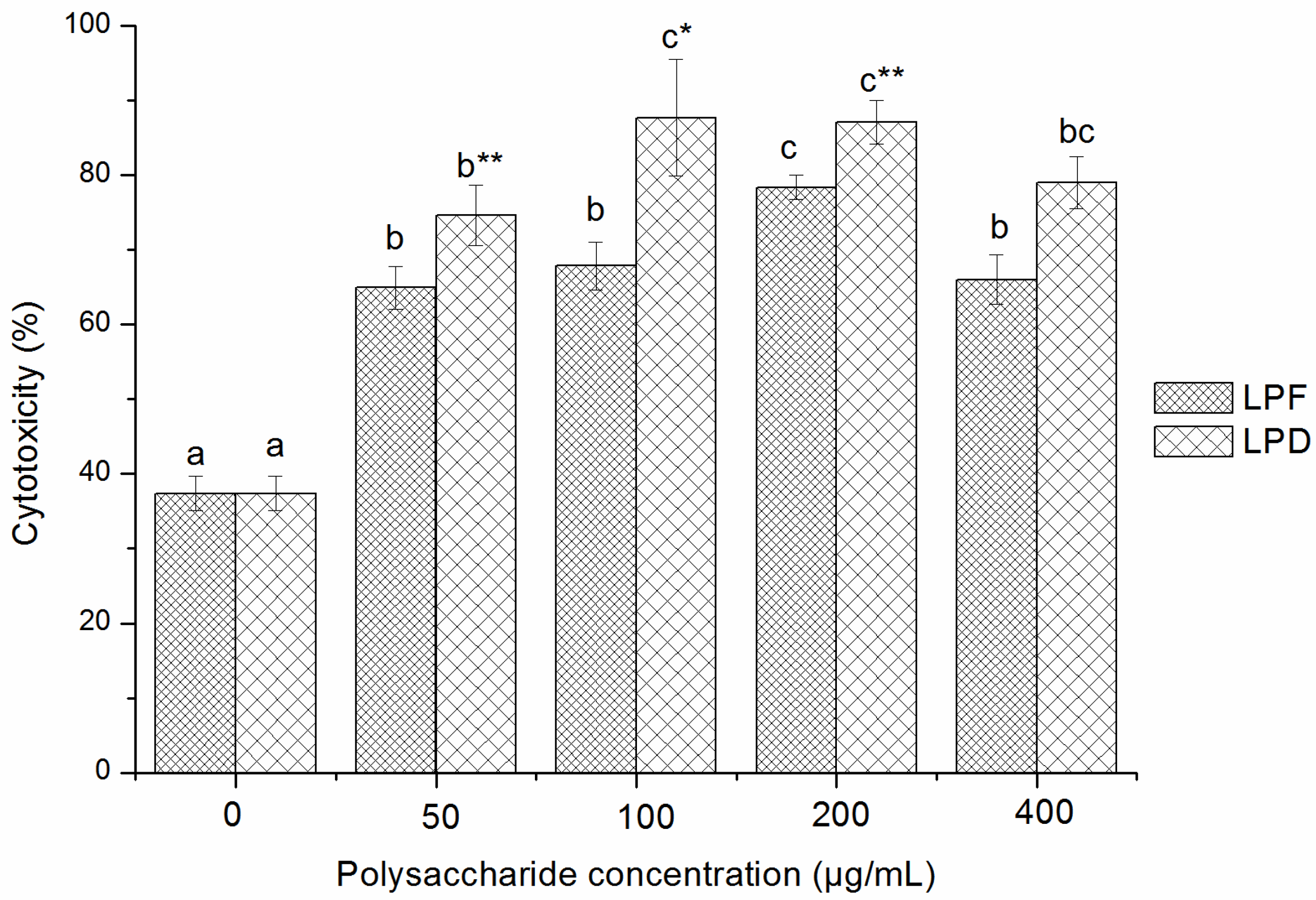
2.3.2. Effects of Litchi Polysaccharides on NK Cells Cytotoxicity
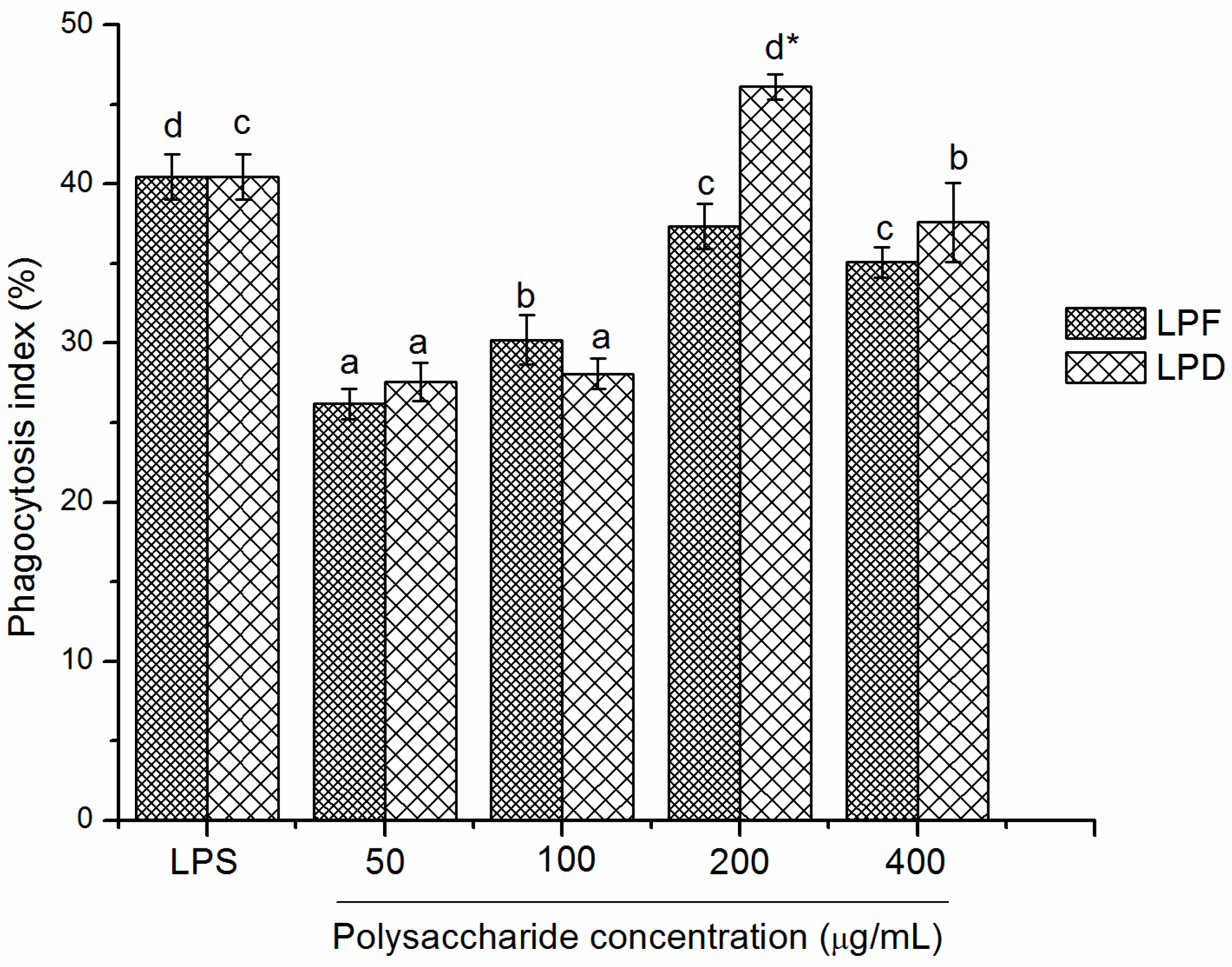
2.3.3. Effects of Litchi Polysaccharides on Phagocytosis of RAW264.7 Mice Macrophages


3. Experimental
3.1. Materials and Chemicals
3.1.1. Chemicals and Reagents
3.1.2. Cells and Animals
3.1.3. Preparation of Litchi Fruit
3.2. Preparation of Crude Litchi Pulp Polysaccharides
3.3. Preliminary Characterization of Litchi Polysaccharides
3.3.1. Analysis of the Chemical Characteristics of Litchi Polysaccharides
3.3.2. Determination of Molecular Weights of Litchi Polysaccharides
3.3.3. UV and IR Spectra Analysis
3.4. In Vitro Inhibition Assay of Tumor Cells Proliferation of Litchi Polysaccharides
3.4.1. Cytotoxicity Assay of Tumor Cells
3.4.2. Inhibition Assay of Tumor Cells Proliferation
3.5. In Vitro Immunostimulatory Activity Assay of Litchi Polysaccharides
3.5.1. Determination of Mouse Splenocyte Proliferation
3.5.2. Cytotoxicity Assay of Natural Killer Cells
3.5.3. Assay of Phagocytosis of RAW264.7 Macrophages
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kong, F.; Zhang, M.; Liao, S.; Yu, S.; Chi, J.; Wei, Z. Antioxidant activity of polysaccharide-enriched fractions extracted from pulp tissue of Litchi Chinensis sonn. Molecules 2010, 15, 2152–2165. [Google Scholar] [CrossRef]
- Jiang, G.; Lin, S.; Wen, L.; Jiang, Y.; Zhao, M.; Chen, F.; Prasad, K.N.; Duan, X.; Yang, B. Identification of a novel phenolic compound in litchi (Litchi chinensis Sonn.) pericarp and bioactivity evaluation. Food Chem. 2013, 136, 563–568. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, H.; Yang, R.; Huang, H.; Sun, Y.; Shen, Y.; Lei, H.; Gao, H. Effects of Litchi chinensis fruit isolates on prostaglandin E(2) and nitric oxide production in J774 murine macrophage cells. BMC Complement. Altern. Med. 2012, 12. [Google Scholar] [CrossRef]
- Wang, X.; Wei, Y.; Yuan, S.; Liu, G.; Zhang, Y.L.J.; Wang, W. Potential anticancer activity of litchi fruit pericarp extract against hepatocellular carcinoma in vitro and in vivo. Cancer Lett. 2006, 239, 144–150. [Google Scholar] [CrossRef]
- Bhoopat, L.; Srichairatanakool, S.; Kanjanapothi, D.; Taesotikul, T.; Thananchai, H.; Bhoopat, T. Hepatoprotective effects of lychee (Litchi chinensis Sonn.): A combination of antioxidant and anti-apoptotic activities. J. Ethnopharmacol. 2011, 136, 55–66. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, B.; Wang, J.; Liu, Y.; Yu, L.; Jiang, Y. Immunomodulatory and anticancer activities of flavonoids extracted from litchi (Litchi chinensis Sonn.) pericarp. Int. Immunopharmacol. 2007, 7, 162–166. [Google Scholar] [CrossRef]
- Hu, X.Q.; Huang, Y.Y.; Dong, Q.F.; Song, L.Y.; Yuan, F.; Yu, R.M. Structure characterization and antioxidant activity of a novel polysaccharide isolated from pulp tissues of Litchi chinensis. J. Agric. Food Chem. 2011, 59, 11548–11552. [Google Scholar]
- Garau, M.C.; Simal, S.; Rosselló, C.; Femenia, A. Effect of air-drying temperature on physico-chemical properties of dietary fibre and antioxidant capacity of orange (Citrus aurantium v. Canoneta) by-products. Food Chem. 2007, 104, 1014–1024. [Google Scholar] [CrossRef]
- Chang, C.-H.; Lin, H.-Y.; Chang, C.-Y.; Liu, Y.-C. Comparisons on the antioxidant properties of fresh, freeze-dried and hot-air-dried tomatoes. J. Food Eng. 2006, 77, 478–485. [Google Scholar] [CrossRef]
- Ferreira, D.; Barros, A.; Coimbra, M.; Delgadillo, I. Use of FT-IR spectroscopy to follow the effect of processing in cell wall polysaccharide extracts of a sun-dried pear. Carbohydr. Polym. 2001, 45, 175–182. [Google Scholar] [CrossRef]
- Ali Asgar, M. Structural features of pectins from fresh and sun-dried Japanese persimmon fruit. Food Chem. 2004, 87, 247–251. [Google Scholar] [CrossRef]
- Thetsrimuang, C.; Khammuang, S.; Chiablaem, K.; Srisomsap, C.; Sarnthima, R. Antioxidant properties and cytotoxicity of crude polysaccharides from Lentinus polychrous Lév. Food Chem. 2011, 128, 634–639. [Google Scholar] [CrossRef]
- Ma, L.; Chen, H.; Zhu, W.; Wang, Z. Effect of different drying methods on physicochemical properties and antioxidant activities of polysaccharides extracted from mushroom Inonotus obliquus. Food Res. Int. 2013, 50, 2152–2165. [Google Scholar]
- Kong, F.-L.; Zhang, M.-W.; Kuang, R.-B.; Yu, S.-J.; Chi, J.-W.; Wei, Z.-C. Antioxidant activities of different fractions of polysaccharide purified from pulp tissue of litchi (Litchi chinensis Sonn.). Carbohydr. Polym. 2010, 81, 612–616. [Google Scholar] [CrossRef]
- Jing, Y.; Huang, L.; Lv, W.; Tong, H.; Song, L.; Hu, X.; Yu, R. Structure Characterization of a Novel Polysaccharide from Pulp Tissues of Litchi chinensis and its Immunomodulatory Activity. J. Agric. Food Chem. 2013, 62, 902–911. [Google Scholar]
- Ducel, V.; Saulnier, P.; Richard, J.; Boury, F. Plant protein-polysaccharide interactions in solutions: Application of soft particle analysis and light scattering measurements. Colloids Surf. 2005, 41, 95–102. [Google Scholar] [CrossRef]
- Satitmanwiwat, S.; Ratanakhanokchai, K.; Laohakunjit, N.; Chao, L.K.; Chen, S.T.; Pason, P.; Tachaapaikoon, C.; Kyu, K.L. Improved purity and immunostimulatory activity of beta-(1–>3) (1–>6)-glucan from Pleurotus sajor-caju using cell wall-degrading enzymes. J. Agric. Food Chem. 2012, 60, 5423–5430. [Google Scholar] [CrossRef]
- Zhang, N.; Chen, H.; Ma, L.; Zhang, Y. Physical modifications of polysaccharide from Inonotus obliquus and the antioxidant properties. Int. J. Biol. Macromol. 2013, 54, 209–215. [Google Scholar] [CrossRef]
- Mirhosseini, H.; Amid, B.T.; Cheong, K.W. Effect of different drying methods on chemical and molecular structure of heteropolysaccharide–protein gum from durian seed. Food Hydrocolloids 2013, 31, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Asgar, M.A.; Yamauchi, R.; Kato, K. Modification of pectin in Japanese persimmon fruit during the sun-drying process. Food Chem. 2003, 81, 555–560. [Google Scholar] [CrossRef]
- Doublier, J.-L.; Garnier, C.; Renard, D.; Sanchez, C. Protein–polysaccharide interactions. Curr. Opin. Colloid Interface Sci. 2000, 5, 202–214. [Google Scholar] [CrossRef]
- Zhang, W. Biochemical Technology of Carbohydrate Complexes; Zhejiang University Press: Hangzhou, China, 1994. [Google Scholar]
- Tian, L.; Zhao, Y.; Guo, C.; Yang, X. A comparative study on the antioxidant activities of an acidic polysaccharide and various solvent extracts derived from herbal Houttuynia cordata. Carbohydr. Polym. 2011, 83, 537–544. [Google Scholar] [CrossRef]
- Xin, T.; Zhang, F.; Jiang, Q.; Chen, C.; Huang, D.; Lv, Y.; Shen, W.; Jin, Y. Purification and antitumor activity of two acidic polysaccharides from the roots of Polygala tenuifolia. Carbohydr. Polym. 2012, 90, 1671–1676. [Google Scholar] [CrossRef]
- Ouchi, T.; Matsumoto, M.; Ihara, K.; Ohya, Y. Synthesis and cytotoxic activity of oxidized galactomannan/ADR conjugate. J. Macromol. Sci. Part A Pure Appl. Chem. 1997, 34, 975–989. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Xiong, Q.; Jiang, C.; Lai, X. Extraction, characterization and biological activities of polysaccharides from Amomum villosum. Carbohydr. Polym. 2013, 95, 114–122. [Google Scholar] [CrossRef]
- Zeng, G.; Ju, Y.; Shen, H.; Zhou, N.; Huang, L. Immunopontentiating activities of the purified polysaccharide from evening primrose in H22 tumor-bearing mice. Int. J. Biol Macromol. 2013, 52, 280–285. [Google Scholar] [CrossRef]
- Yi, Y.; Liao, S.T.; Zhang, M.W.; Shi, J.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C. Physicochemical characteristics and immunomodulatory activities of three polysaccharide-protein complexes of longan pulp. Molecules 2011, 16, 6148–6164. [Google Scholar] [CrossRef]
- Iribe, H.; Koga, T. Augmentation of the proliferative response of thymocytes to phytohemagglutinin by the muramyl dipeptide. Cell Immunol. 1984, 88, 9–15. [Google Scholar]
- Kuang, H.; Xia, Y.; Yang, B.; Wang, Q.; Wang, Y. Screening and comparison of the immunosuppressive activities of polysaccharides from the stems of Ephedra sinica Stapf. Carbohydr. Polym. 2011, 83, 787–795. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, K. Characteristic immunostimulation by MAP, a polysaccharide isolated from the mucus of the loach, Misgurnus anguillicaudatus. Carbohydr. Polym. 2005, 59, 75–82. [Google Scholar] [CrossRef]
- Lin, K.-I.; Kao, Y.-Y.; Kuo, H.-K.; Yang, W.-B.; Chou, A.; Lin, H.-H.; Alice, L.Y.; Wong, C.-H. Reishi polysaccharides induce immunoglobulin production through the TLR4/TLR2-mediated induction of transcription factor Blimp-1. J. Biol. Chem. 2006, 281, 24111–24123. [Google Scholar]
- Sun, Y.; Sun, T.; Wang, F.; Zhang, J.; Li, C.; Chen, X.; Li, Q.; Sun, S. A polysaccharide from the fungi of Huaier exhibits anti-tumor potential and immunomodulatory effects. Carbohydr. Polym. 2013, 92, 577–582. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, H.; Zhang, X.; Tong, H.; Liu, J. Structural elucidation and immunological activity of a polysaccharide from the fruiting body of Armillaria mellea. Bioresour. Technol. 2009, 100, 1860–1863. [Google Scholar] [CrossRef]
- Kralovec, J.; Metera, K.; Kumar, J.; Watson, L.; Girouard, G.; Guan, Y.; Carr, R.; Barrow, C.; Ewart, H. Immunostimulatory principles from Chlorella p yrenoidosa—Part 1: Isolation and biological assessment in vitro. Phytomedicine 2007, 14, 57–64. [Google Scholar] [CrossRef]
- Lo, T.C.; Jiang, Y.H.; Chao, A.L.; Chang, C.A. Use of statistical methods to find the polysaccharide structural characteristics and the relationships between monosaccharide composition ratio and macrophage stimulatory activity of regionally different strains of Lentinula edodes. Anal. Chim. Acta 2007, 584, 50–56. [Google Scholar] [CrossRef]
- Sevag, M.; Lackman, D.B.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 1938, 425–436. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 3, 350–356. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Felice, D.L.; Sun, J.; Liu, R.H. A modified methylene blue assay for accurate cell counting. J. Funct. Foods 2009, 1, 109–118. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, M.-W.; Liao, S.-T.; Zhang, R.-F.; Deng, Y.-Y.; Wei, Z.-C.; Tang, X.-J.; Zhang, Y. Structural features and immunomodulatory activities of polysaccharides of longan pulp. Carbohydr. Polym. 2012, 87, 636–643. [Google Scholar] [CrossRef]
- Sample Availability: Samples of polysaccharides are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, F.; Zhang, R.; Yi, Y.; Tang, X.; Zhang, M.; Su, D.; Deng, Y.; Wei, Z. Comparison of Physicochemical Properties and Immunomodulatory Activity of Polysaccharides from Fresh and Dried Litchi Pulp. Molecules 2014, 19, 3909-3925. https://doi.org/10.3390/molecules19043909
Huang F, Zhang R, Yi Y, Tang X, Zhang M, Su D, Deng Y, Wei Z. Comparison of Physicochemical Properties and Immunomodulatory Activity of Polysaccharides from Fresh and Dried Litchi Pulp. Molecules. 2014; 19(4):3909-3925. https://doi.org/10.3390/molecules19043909
Chicago/Turabian StyleHuang, Fei, Ruifen Zhang, Yang Yi, Xiaojun Tang, Mingwei Zhang, Dongxiao Su, Yuanyuan Deng, and Zhencheng Wei. 2014. "Comparison of Physicochemical Properties and Immunomodulatory Activity of Polysaccharides from Fresh and Dried Litchi Pulp" Molecules 19, no. 4: 3909-3925. https://doi.org/10.3390/molecules19043909