β-Asarone Reverses Chronic Unpredictable Mild Stress-Induced Depression-Like Behavior and Promotes Hippocampal Neurogenesis in Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
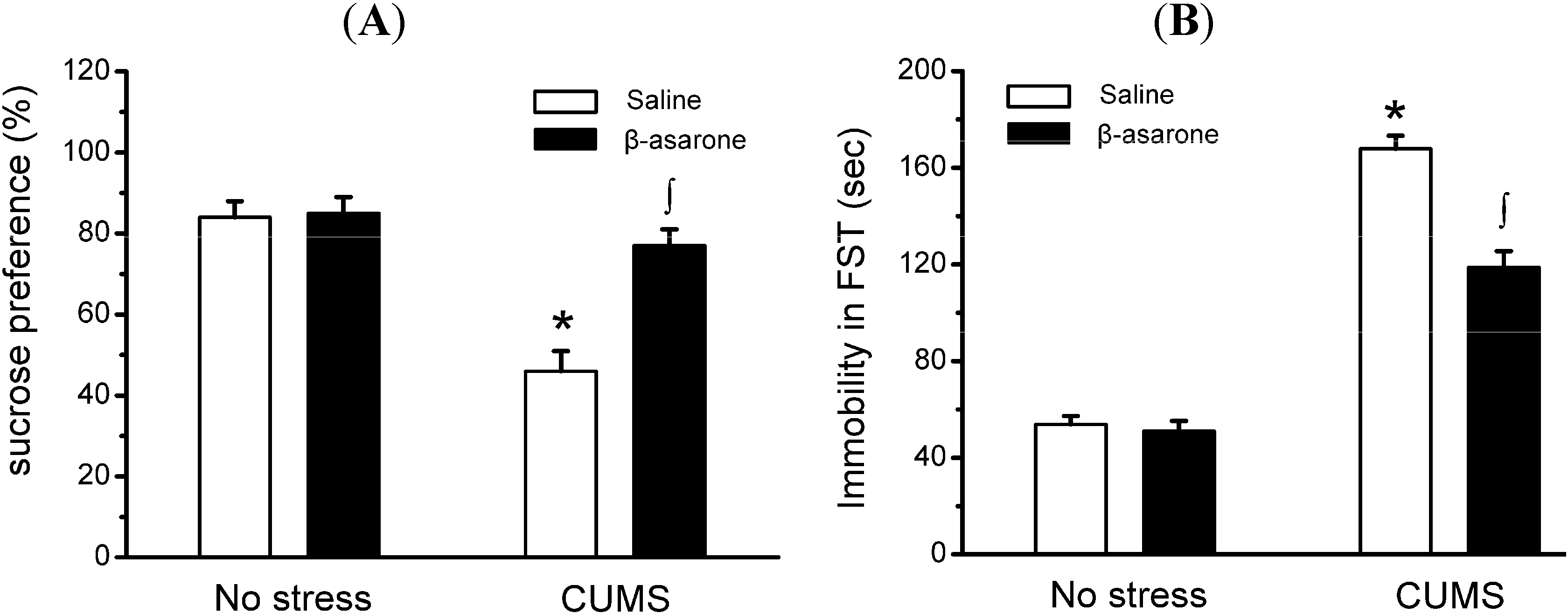
2.1. β-Asarone Treatment Produces Antidepressant-Like Behavioral Responses in CUMS-Exposed Rats
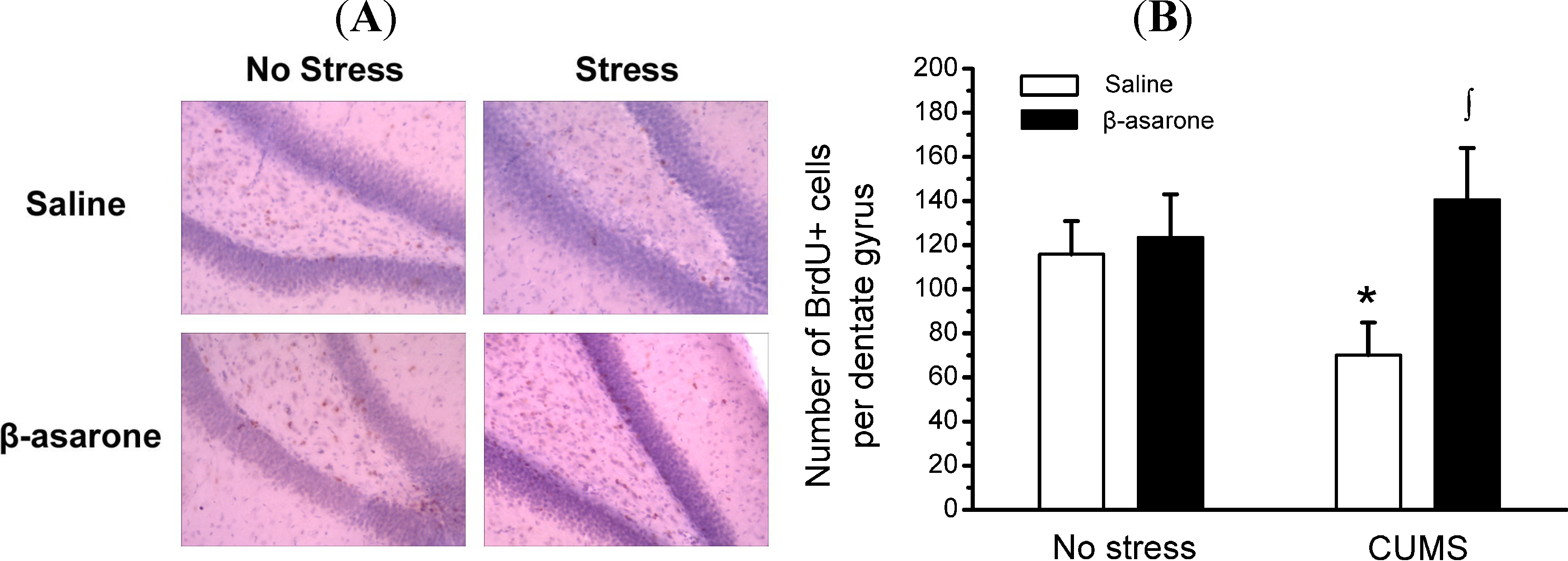
2.2. β-Asarone Increases Hippocampal Neurons Neurogenesis in CUMS-Exposed Rats


2.3. Involvement of ERK1/2 and CREB Phosphorylation in Neuroprotection Effects of β-Asarone in CUMS-Exposed Rats
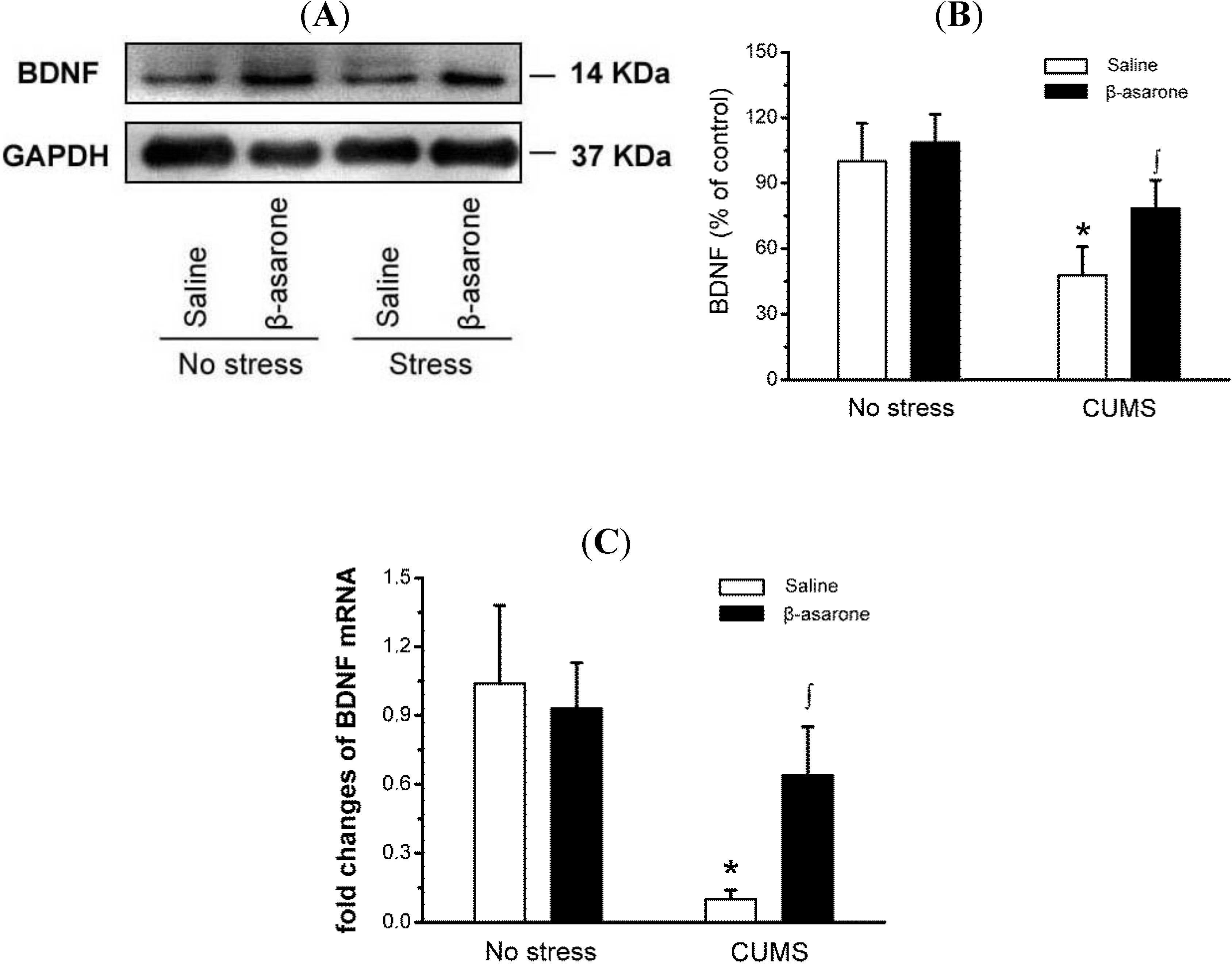
2.4. β-Asarone Increased BDNF Expression in CUMS-Exposed Rats
2.5. Discussion


3. Experimental Section
3.1. Animals
3.2. CUMS Procedure
3.3. β-Asarone Preparation and Treatment Protocols
3.4. Sucrose Preference Test (SPT)
3.5. Forced Swim Test (FST)
3.6. BrdU Labeling and Immunohistochemical Staining
3.7. RNA Isolation and Real-Time Polymerase Chain Reaction (PCR)
3.8. Western Blot
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barcelos-Ferreira, R.; Nakano, E.Y.; Steffens, D.C.; Bottino, C.M. Quality of life and physical activity associated to lower prevalence of depression in community-dwelling elderly subjects from Sao Paulo. J. Affect. Disord. 2013, 150, 616–622. [Google Scholar] [CrossRef]
- Michl, L.C.; McLaughlin, K.A.; Shepherd, K.; Nolen-Hoeksema, S. Rumination as a mechanism linking stressful life events to symptoms of depression and anxiety: Longitudinal evidence in early adolescents and adults. J. Abnorm. Psychol. 2013, 122, 339–352. [Google Scholar] [CrossRef]
- Yadid, G.; Friedman, A. Dynamics of the dopaminergic system as a key component to the understanding of depression. Prog. Brain Res. 2008, 172, 265–286. [Google Scholar] [CrossRef]
- Voleti, B.; Duman, R.S. The roles of neurotrophic factor and Wnt signaling in depression. Clin. Pharmacol. Ther. 2012, 91, 333–338. [Google Scholar] [CrossRef]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Fries, G.R.; Kapczinski, F.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Quevedo, J. Chronic administration of harmine elicits antidepressant-like effects and increases BDNF levels in rat hippocampus. J. Neural. Transm. 2010, 117, 1131–1137. [Google Scholar] [CrossRef]
- Malki, K.; Lourdusamy, A.; Binder, E.; Payá-Cano, J.; Sluyter, F.; Craig, I.; Keers, R.; McGuffin, P.; Uher, R.; Schalkwyk, L.C. Antidepressant-dependent mRNA changes in mouse associated with hippocampal neurogenesis in a mouse model of depression. Pharmacogenet. Genom. 2012, 22, 765–776. [Google Scholar] [CrossRef]
- Guilloux, J.P.; Mendez-David, I.; Pehrson, A.; Guiard, B.P.; Repérant, C.; Orvoën, S.; Gardier, A.M.; Hen, R.; Ebert, B.; Miller, S.; et al. Antidepressant and anxiolytic potential of the multimodal antidepressant vortioxetine (Lu AA21004) assessed by behavioural and neurogenesis outcomes in mice. Neuropharmacology 2013, 73, 147–159. [Google Scholar] [CrossRef]
- Den Heijer, T.; Tiemeier, H.; Luijendijk, H.J.; van der Lijn, F.; Koudstaal, P.J.; Hofman, A.; Breteler, M.M. A study of the bidirectional association between hippocampal volume on magnetic resonance imaging and depression in the elderly. Biol. Psychiat. 2011, 70, 191–197. [Google Scholar] [CrossRef]
- Kaae, S.S.; Chen, F.; Wegener, G.; Madsen, T.M.; Nyengaard, J.R. Quantitative hippocampal structural changes following electroconvulsive seizure treatment in a rat model of depression. Synapse 2012, 66, 667–676. [Google Scholar] [CrossRef]
- Joëls, M.; Karst, H.; Alfarez, D.; Heine, V.M.; Qin, Y.; van Riel, E.; Verkuyl, M.; Lucassen, P.J.; Krugers, H.J. Effects of chronic stress on structure and cell function in rat hippocampus and hypothalamus. Stress 2004, 7, 221–231. [Google Scholar] [CrossRef]
- Bianchi, M.; Hagan, J.J.; Heidbreder, C.A. Neuronal plasticity, stress and depression: Involvement of the cytoskeletal microtubular system? Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 597–611. [Google Scholar] [CrossRef]
- Boldrini, M.; Underwood, M.D.; Hen, R.; Rosoklija, G.B.; Dwork, A.J.; John Mann, J.; Arango, V. Antidepressants increase neural progenitor cells in the human hippocampus. Neuropsychopharmacology 2009, 34, 2376–2389. [Google Scholar] [CrossRef]
- Chen, F.; Madsen, T.M.; Wegener, G.; Nyengaard, J.R. Imipramine treatment increases the number of hippocampal synapses and neurons in a genetic animal model of depression. Hippocampus 2010, 20, 1376–1384. [Google Scholar] [CrossRef]
- Chen, B.; Dowlatshahi, D.; MacQueen, G.M.; Wang, J.F.; Young, L.T. Increased hippocampal BDNF immunoreactivity in subjects treated with antidepressant medication. Biol. Psychiat. 2001, 50, 260–265. [Google Scholar] [CrossRef]
- Lindholm, J.S.; Autio, H.; Vesa, L.; Antila, H.; Lindemann, L.; Hoener, M.C.; Skolnick, P.; Rantamäki, T.; Castrén, E. The antidepressant-like effects of glutamatergic drugs ketamine and AMPA receptor potentiator LY 451646 are preserved in bdnf+/− heterozygous null mice. Neuropharmacology 2012, 62, 391–397. [Google Scholar] [CrossRef]
- Alboni, S.; Tascedda, F.; Corsini, D.; Benatti, C.; Caggia, F.; Capone, G.; Barden, N.; Blom, J.M.; Brunello, N. Stress induces altered CRE/CREB pathway activity and BDNF expression in the hippocampus of glucocorticoid receptor-impaired mice. Neuropharmacology 2011, 60, 1337–1346. [Google Scholar] [CrossRef]
- Einat, H.; Yuan, P.; Gould, T.D.; Li, J.; Du, J.; Zhang, L.; Manji, H.K.; Chen, G. The role of the extracellular signal-regulated kinase signaling pathway in mood modulation. J. Neurosci. 2003, 23, 7311–7316. [Google Scholar]
- Gourley, S.L.; Wu, F.J.; Kiraly, D.D.; Ploski, J.E.; Kedves, A.T.; Duman, R.S.; Taylor, J.R. Regionally specific regulation of ERK MAP kinase in a model of antidepressant-sensitive chronic depression. Biol. Psychiat. 2008, 63, 353–359. [Google Scholar] [CrossRef]
- Jeon, S.J.; Rhee, S.Y.; Seo, J.E.; Bak, H.R.; Lee, S.H.; Ryu, J.H.; Cheong, J.H.; Shin, C.Y.; Kim, G.H.; Lee, Y.S. Oroxylin A increases BDNF production by activation of MAPK-CREB pathway in rat primary cortical neuronal culture. Neurosci. Res. 2011, 69, 214–222. [Google Scholar] [CrossRef]
- Stahl, S.M.; Felker, A. Monoamine oxidase inhibitors: A modern guide to an unrequited class of antidepressants. CNS Spectr. 2008, 13, 855–870. [Google Scholar]
- Wu, H.B.; Fang, Y.Q. Pharmacokinetics of beta-asarone in rats. Yao Xue Xue Bao 2004, 39, 836–838. [Google Scholar]
- Li, Z.; Zhao, G.; Qian, S.; Yang, Z.; Chen, X.; Chen, J.; Cai, C.; Liang, X.; Guo, J. Cerebrovascular protection of β-asarone in Alzheimer’s disease rats: A behavioral, cerebral blood flow, biochemical and genic study. J. Ethnopharmacol. 2012, 144, 305–312. [Google Scholar] [CrossRef]
- Zou, D.J.; Wang, G.; Liu, J.C.; Dong, M.X.; Li, X.M.; Zhang, C.; Zhou, L.; Wang, R.; Niu, Y.C. Beta-asarone attenuates beta-amyloid-induced apoptosis through the inhibition of the activation of apoptosis signal-regulating kinase 1 in SH-SY5Y cells. Pharmazie 2011, 66, 44–51. [Google Scholar]
- Ossowska, G.; Danilczuk, Z.; Klenk-Majewska, B.; Czajkowski, L.; Zebrowska-Łupina, I. Antidepressants in chronic unpredictable mild stress (CUMS)-induced deficit of fighting behavior. Pol. J. Pharmacol. 2004, 56, 305–311. [Google Scholar]
- Yazir, Y.; Utkan, T.; Aricioglu, F. Inhibition of neuronal nitric oxide synthase and soluble guanylate cyclase prevents depression-like behaviour in rats exposed to chronic unpredictable mild stress. Basic Clin. Pharmacol. Toxicol. 2012, 111, 154–160. [Google Scholar]
- Banasr, M.; Valentine, G.W.; Li, X.Y.; Gourley, S.L.; Taylor, J.R.; Duman, R.S. Chronic unpredictable stress decreases cell proliferation in the cerebral cortex of the adult rat. Biol. Psychiat. 2007, 62, 496–504. [Google Scholar] [CrossRef]
- Qi, X.; Lin, W.; Li, J.; Li, H.; Wang, W.; Wang, D.; Sun, M. Fluoxetine increases the activity of the ERK-CREB signal system and alleviates the depressive-like behavior in rats exposed to chronic forced swim stress. Neurobiol. Dis. 2008, 31, 278–285. [Google Scholar] [CrossRef]
- Carlini, V.P.; Poretti, M.B.; Rask-Andersen, M.; Chavan, R.A.; Ponzio, M.F.; Sawant, R.S.; de Barioglio, S.R.; Schiöth, H.B.; de Cuneo, M.F. Differential effects of fluoxetine and venlafaxine on memory recognition: Possible mechanisms of action. Prog. Neuropsychopharmacol. Biol. Psychiat. 2012, 38, 159–167. [Google Scholar]
- Cazzin, C.; Mion, S.; Caldara, F.; Rimland, J.M.; Domenici, E. Microarray analysis of cultured rat hippocampal neurons treated with brain derived neurotrophic factor. Mol. Biol. Rep. 2011, 38, 983–990. [Google Scholar] [CrossRef]
- Zoon, H.F.; Veth, C.P.; Arns, M.; Drinkenburg, W.H.; Talloen, W.; Peeters, P.J.; Kenemans, J.L. EEG alpha power as an intermediate measure between brain-derived neurotrophic factor Val66Met and depression severity in patients with major depressive disorder. J. Clin. Neurophysiol. 2011, 30, 261–267. [Google Scholar]
- Ye, Y.; Wang, G.; Wang, H.; Wang, X. Brain-derived neurotrophic factor (BDNF) infusion restored astrocytic plasticity in the hippocampus of a rat model of depression. Neurosci. Lett. 2011, 503, 15–19. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xian, Y.F.; Ip, S.P.; Tsai, S.H.; Che, C.T. Long-term treatment with peony glycosides reverses chronic unpredictable mild stress-induced depressive-like behavior via increasing expression of neurotrophins in rat brain. Behav. Brain Res. 2010, 210, 171–177. [Google Scholar] [CrossRef]
- Papp, M.; Willner, P.; Muscat, R. An animal model of anhedonia: Attenuation of sucrose consumption and place preference conditioning by chronic unpredictable mild stress. Psychopharmacology (Berl) 1991, 104, 255–259. [Google Scholar] [CrossRef]
- Chourbaji, S.; Zacher, C.; Sanchis-Segura, C.; Dormann, C.; Vollmayr, B.; Gass, P. Learned helplessness: Validity and reliability of depressive-like states in mice. Brain Res. Protoc. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Matthews, K.; Forbes, N.; Reid, I.C. Sucrose consumption as an hedonic measure following chronic unpredictable mild stress. Physiol. Behav. 1995, 57, 241–248. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhou, Y.Z.; Li, Z.F.; Cui, J.; Li, Z.Y.; Gao, X.X.; Sun, H.F.; Zhang, L.Z.; Du, G.H.; Qin, X.M. Anti-depressant effects of Xiaoyaosan on rat model of chronic unpredictable mild stress: A plasma metabonomics study based on NMR spectroscopy. J. Pharm. Pharmacol. 2012, 64, 578–588. [Google Scholar] [CrossRef]
- Han, P.; Han, T.; Peng, W.; Wang, X.R. Antidepressant-like effects of essential oil and asarone, a major essential oil component from the rhizome of Acorus tatarinowii. Pharm. Biol. 2013, 51, 589–594. [Google Scholar] [CrossRef]
- King, C.A.; Knox, M.S.; Henninger, N.; Nguyen, T.A.; Ghaziuddin, N.; Maker, A.; Hanna, G.L. Major depressive disorder in adolescents: Family psychiatric history predicts severe behavioral disinhibition. J. Affect. Disord. 2006, 90, 111–121. [Google Scholar] [CrossRef]
- Farin, A.; Liu, C.Y.; Langmoen, I.A.; Apuzzo, M.L. The biological restoration of central nervous system architecture and function: Part 2-emergence of the realization of adult neurogenesis. Neurosurgery 2009, 64, 581–601. [Google Scholar] [CrossRef]
- Massart, R.; Mongeau, R.; Lanfumey, L. Beyond the monoaminergic hypothesis: Neuroplasticity and epigenetic changes in a transgenic mouse model of depression. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2485–2494. [Google Scholar] [CrossRef]
- Lee, B.H.; Kim, Y.K. The roles of BDNF in the pathophysiology of major depression and in antidepressant treatment. Psychiat. Investig. 2010, 7, 231–235. [Google Scholar] [CrossRef]
- Perera, T.D.; Dwork, A.J.; Keegan, K.A.; Thirumangalakudi, L.; Lipira, C.M.; Joyce, N.; Lange, C.; Higley, J.D.; Rosoklija, G.; Hen, R.; et al. Necessity of hippocampal neurogenesis for the therapeutic action of antidepressants in adult nonhuman primates. PLoS One 2011, 6, e17600. [Google Scholar] [CrossRef]
- Viveros, M.P.; Marco, E.M.; Llorente, R.; Lamota, L. The role of the hippocampus in mediating emotional responses to nicotine and cannabinoids: A possible neural substrate for functional interactions. Behav. Pharmacol. 2007, 18, 375–389. [Google Scholar]
- Tanti, A.; Belzung, C. Hippocampal neurogenesis: A biomarker for depression or antidepressant effects? Methodological considerations and perspectives for future research. Cell Tissue Res. 2013, 354, 203–219. [Google Scholar] [CrossRef]
- Mueller, T.; Wullimann, M.F. BrdU-, neuroD (nrd)- and Hu-studies reveal unusual non-ventricular neurogenesis in the postembryonic zebrafish forebrain. Mech. Dev. 2002, 117, 123–135. [Google Scholar]
- Zhou, X.; Moon, C.; Zheng, F.; Luo, Y.; Soellner, D.; Nuñez, J.L.; Wang, H. N-methyl-d-aspartate-stimulated ERK1/2 signaling and the transcriptional up-regulation of plasticity-related genes are developmentally regulated following in vitro neuronal maturation. J. Neurosci. Res. 2009, 87, 2632–2644. [Google Scholar] [CrossRef]
- Suzuki, S.; al-Noori, S.; Butt, S.A.; Pham, T.A. Regulation of the CREB signaling cascade in the visual cortex by visual experience and neuronal activity. J. Comp. Neurol. 2004, 479, 70–83. [Google Scholar]
- Ha, S.; Redmond, L. ERK mediates activity dependent neuronal complexity via sustained activity and CREB-mediated signaling. Dev. Neurobiol. 2008, 68, 1565–1579. [Google Scholar] [CrossRef]
- Ring, R.M.; Regan, C.M. Captodiamine, a putative antidepressant, enhances hypothalamic BDNF expression in vivo by synergistic 5-HT2c receptor antagonism and sigma-1 receptor agonism. J. Psychopharmacol. 2013, 27, 930–939. [Google Scholar] [CrossRef]
- Xiao, Z.; Liu, W.; Gao, K.; Wan, Q.; Yang, C.; Wang, H.; Wang, X.; Wang, G.; Liu, Z. Interaction between CRHR1 and BDNF genes increases the risk of recurrent major depressive disorder in Chinese population. PLoS One 2011, 6, e28733. [Google Scholar]
- Castrén, E.; Rantamäki, T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Dev. Neurobiol. 2010, 70, 289–297. [Google Scholar] [CrossRef]
- Yeh, C.M.; Huang, C.C.; Hsu, K.S. Prenatal stress alters hippocampal synaptic plasticity in young rat offspring through preventing the proteolytic conversion of pro-brain-derived neurotrophic factor (BDNF) to mature BDNF. J. Physiol. 2012, 590, 991–1010. [Google Scholar]
- Satomura, E.; Baba, H.; Nakano, Y.; Maeshima, H.; Suzuki, T.; Arai, H. Correlations between brain-derived neurotrophic factor and clinical symptoms in medicated patients with major depression. J. Affect. Disord. 2011, 135, 332–335. [Google Scholar] [CrossRef]
- Sarchiapone, M.; Carli, V.; Roy, A.; Iacoviello, L.; Cuomo, C.; Latella, M.C.; di Giannantonio, M.; Janiri, L.; de Gaetano, M.; Janal, M.N. Association of polymorphism (Val66Met) of brain-derived neurotrophic factor with suicide attempts in depressed patients. Neuropsychobiology 2008, 57, 139–145. [Google Scholar] [CrossRef]
- Hill, M.N.; Hellemans, K.G.; Verma, P.; Gorzalka, B.B.; Weinberg, J. Neurobiology of chronic mild stress: Parallels to major depression. Neurosci. Biobehav. Rev. 2012, 36, 2085–2117. [Google Scholar] [CrossRef]
- Dai, J.; Ha, C.; Shen, M. Systematic study of beta-asarone-rich volatile oil from Acori graminei rhizoma by off-line supercritical CO2 extraction-gas chromatography-mass spectrometry. J. Sep. Sci. 2008, 31, 714–720. [Google Scholar] [CrossRef]
- Adnan, M.; Morton, G.; Hadi, S. Analysis of rpoS and bolA gene expression under various stress-induced environments in planktonic and biofilm phase using 2(−ΔΔCT) method. Mol. Cell. Biochem. 2011, 357, 275–282. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds (beta-Asarone) are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dong, H.; Gao, Z.; Rong, H.; Jin, M.; Zhang, X. β-Asarone Reverses Chronic Unpredictable Mild Stress-Induced Depression-Like Behavior and Promotes Hippocampal Neurogenesis in Rats. Molecules 2014, 19, 5634-5649. https://doi.org/10.3390/molecules19055634
Dong H, Gao Z, Rong H, Jin M, Zhang X. β-Asarone Reverses Chronic Unpredictable Mild Stress-Induced Depression-Like Behavior and Promotes Hippocampal Neurogenesis in Rats. Molecules. 2014; 19(5):5634-5649. https://doi.org/10.3390/molecules19055634
Chicago/Turabian StyleDong, Haiying, Zhiying Gao, Hua Rong, Ming Jin, and Xiaojie Zhang. 2014. "β-Asarone Reverses Chronic Unpredictable Mild Stress-Induced Depression-Like Behavior and Promotes Hippocampal Neurogenesis in Rats" Molecules 19, no. 5: 5634-5649. https://doi.org/10.3390/molecules19055634