An Isoflavone from Dipteryx alata Vogel is Active against the in Vitro Neuromuscular Paralysis of Bothrops jararacussu Snake Venom and Bothropstoxin I, and Prevents Venom-Induced Myonecrosis

 and
and
Abstract
:1. Introduction
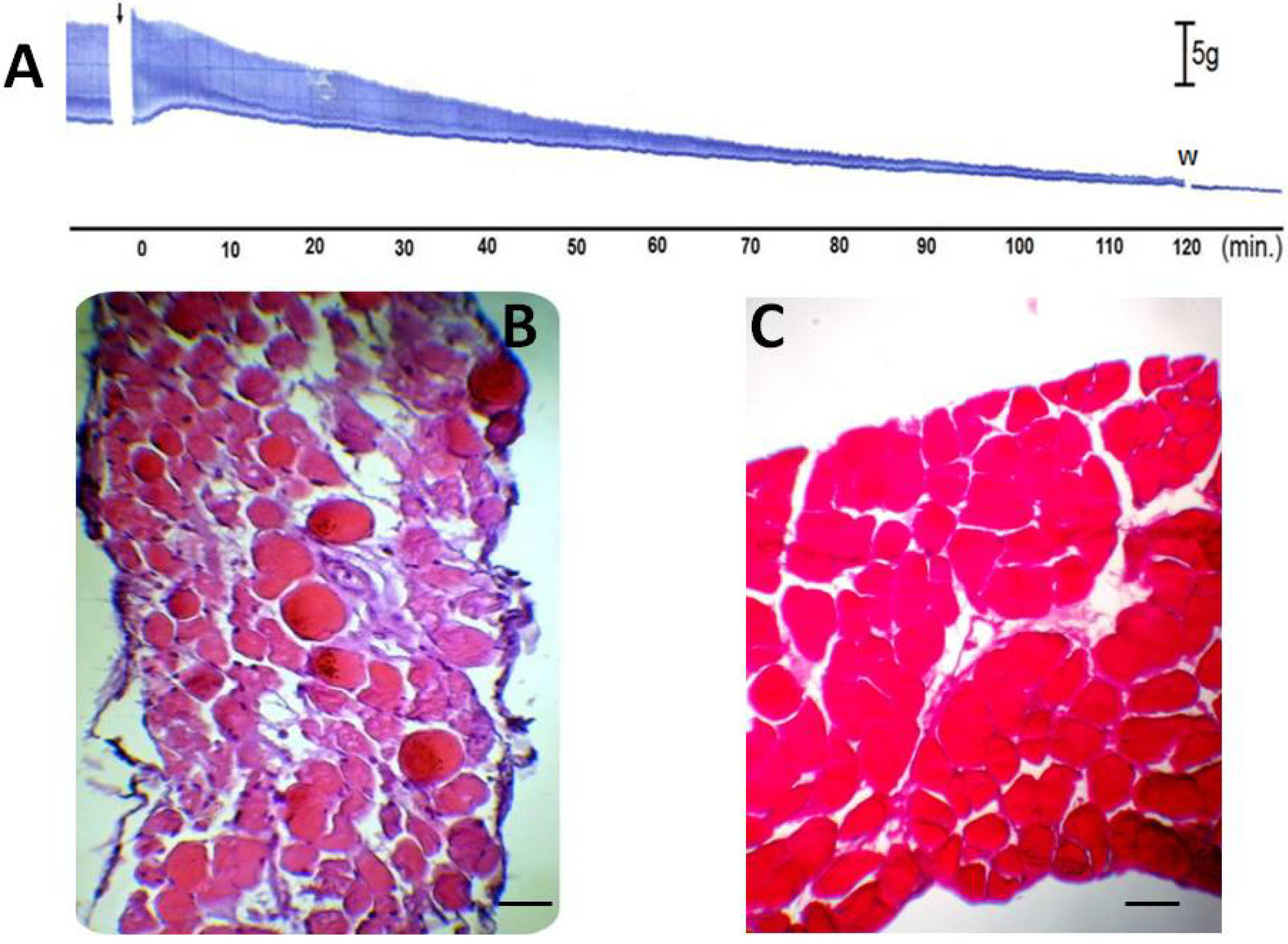
2. Results and Discussion
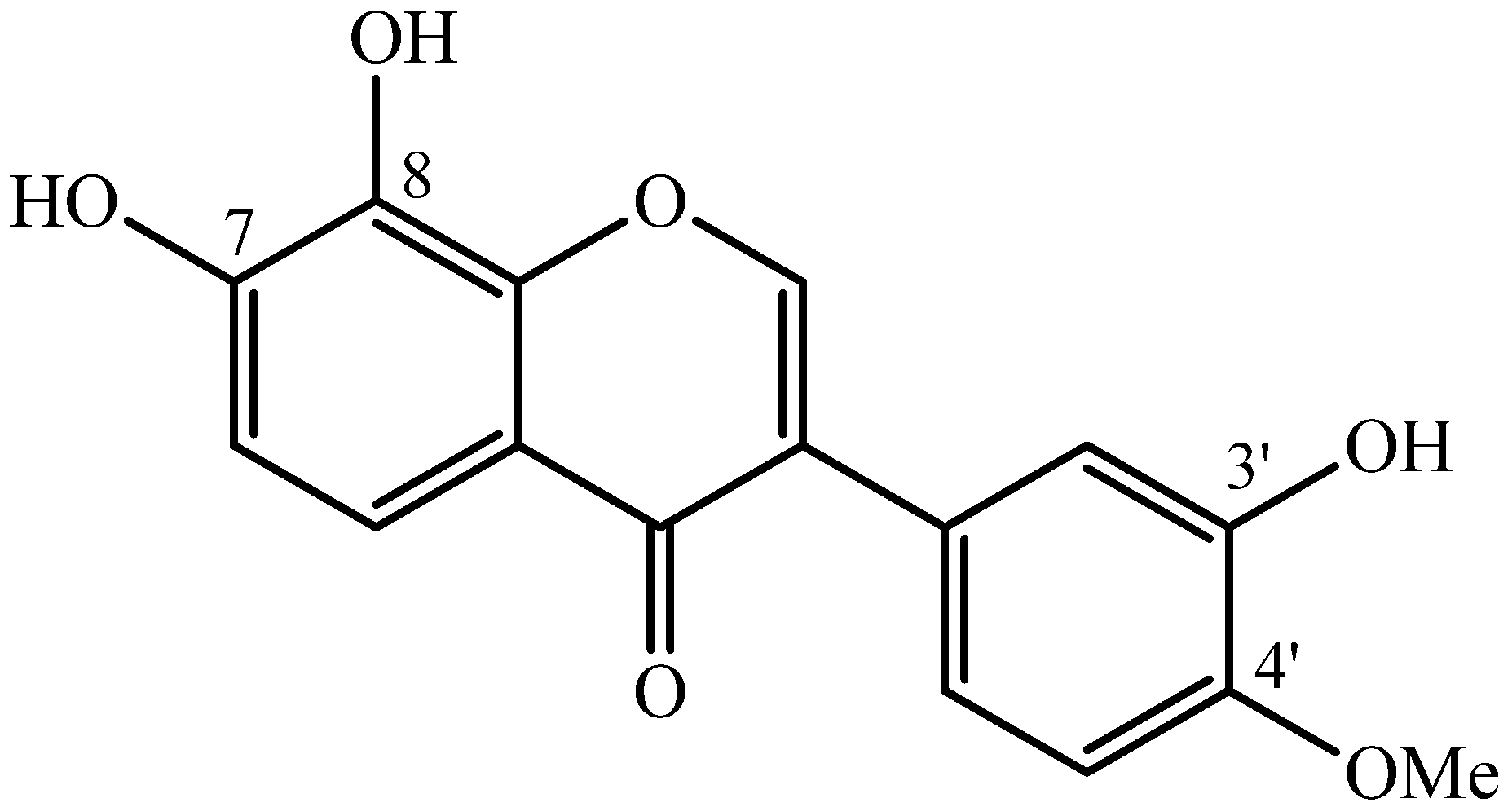
2.1. Molecule

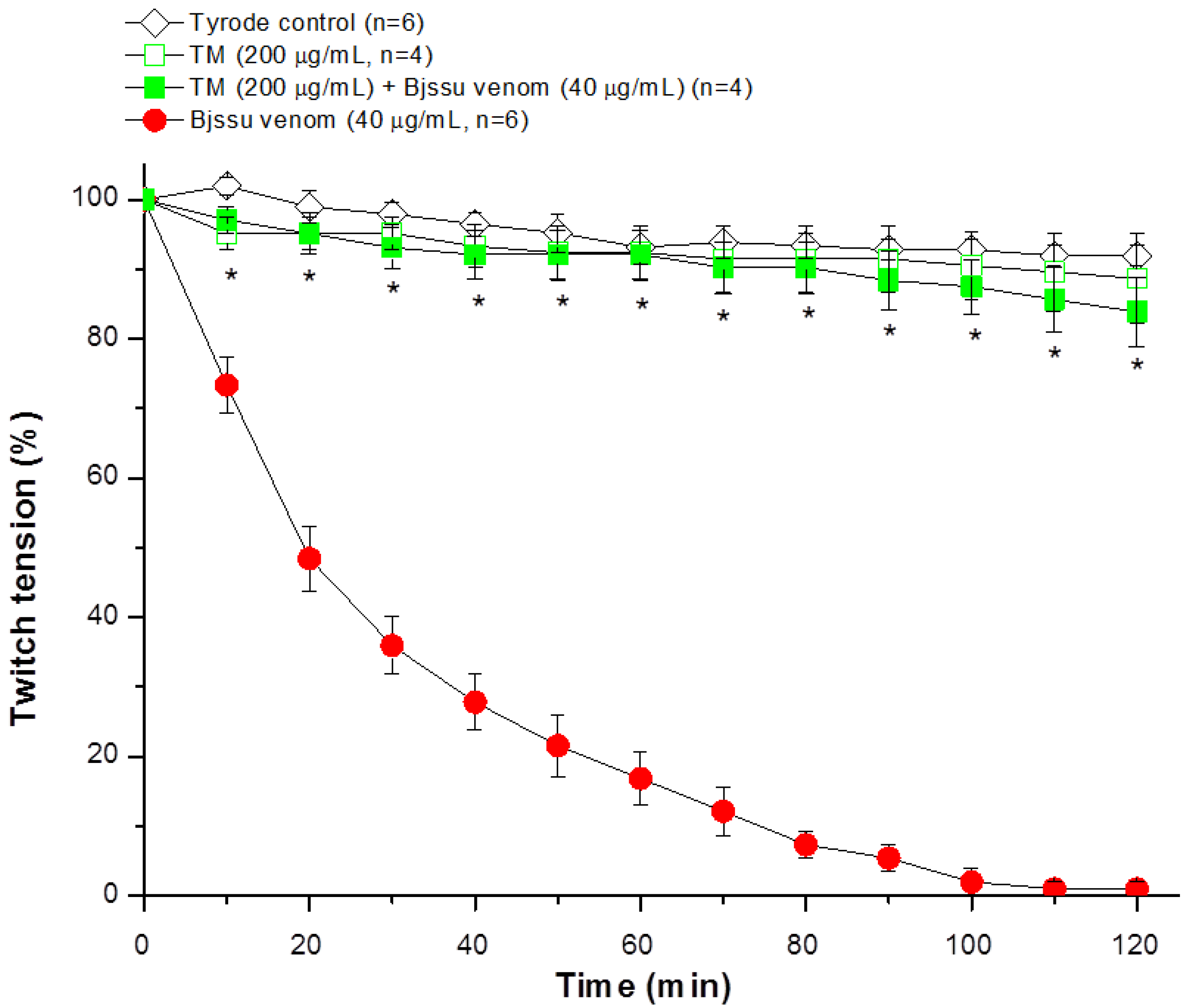
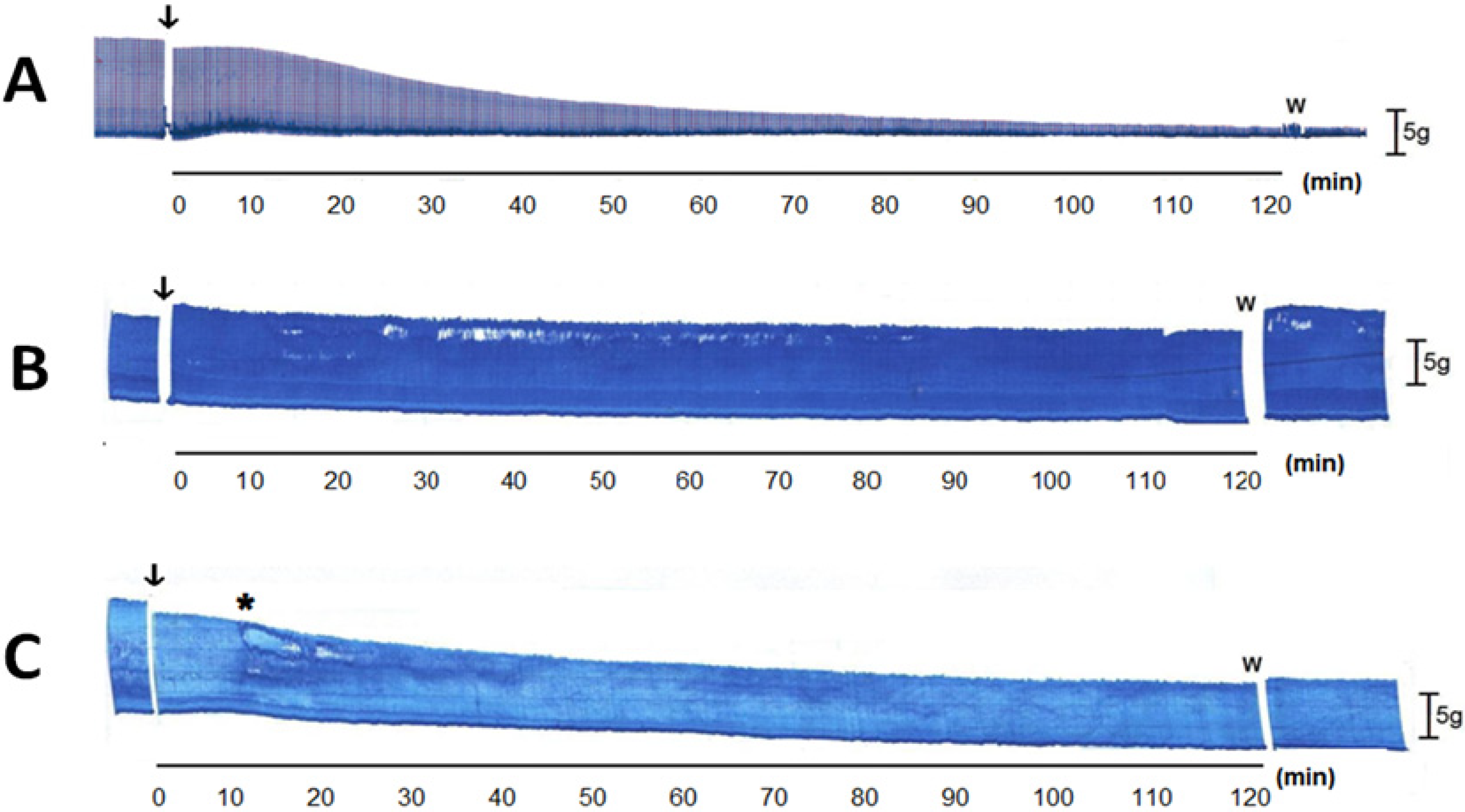
2.2. Pharmacological Assays


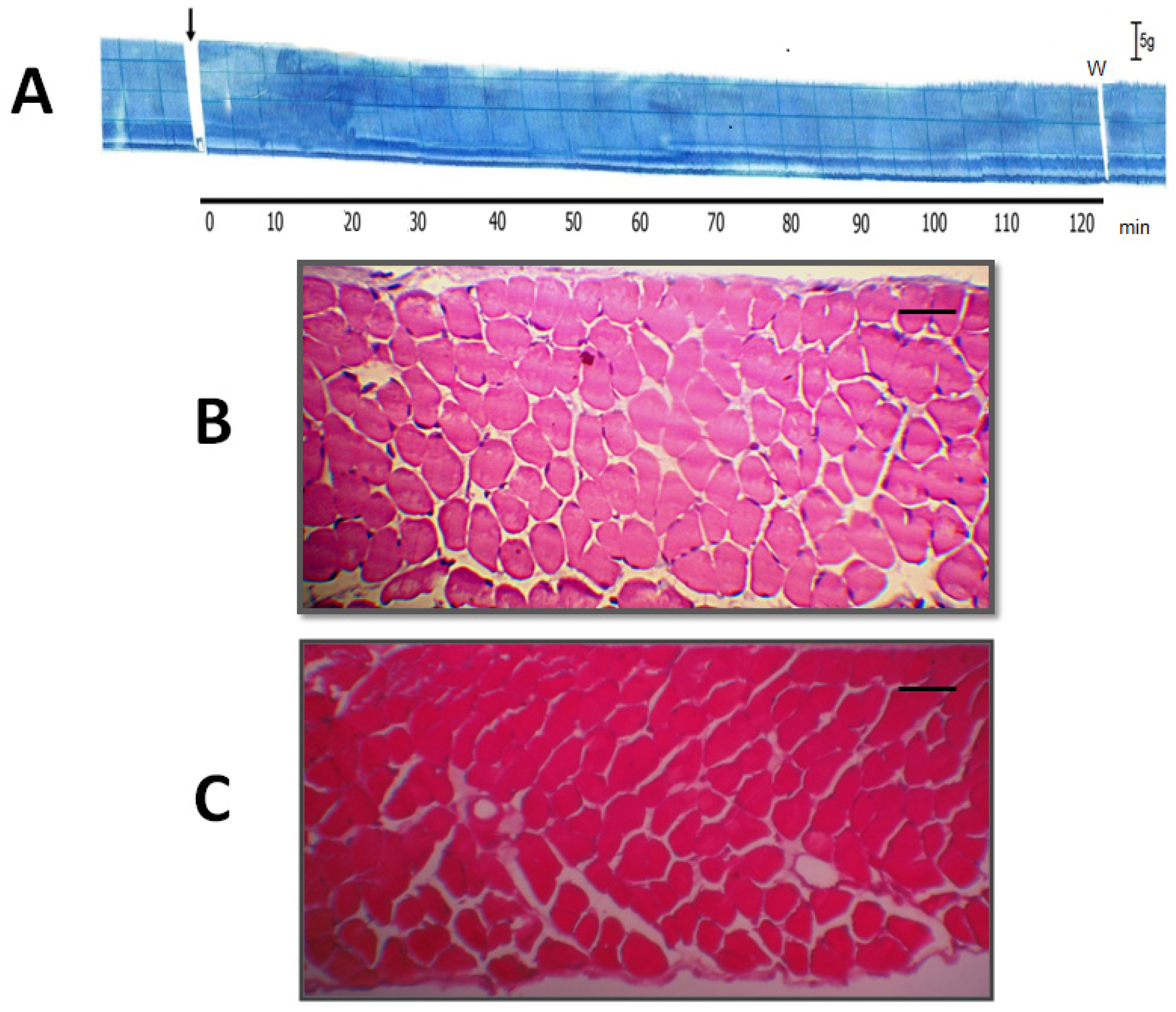
2.3. Quantitative Histological Analysis


2.4. Salmonella Mutagenicity Assay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TM | ||||
|---|---|---|---|---|
| Treatment | TA 98 | TA 97a | ||
| mg/plate | −S9 | +S9 | −S9 | +S9 |
| DMSO | 20 ± 2 | 30 ± 2 | 151 ± 8 | 143 ± 8 |
| 0.19 | 20 ± 4 (1.0) | 34 ± 3 (1.1) | 194 ± 4 (1.3) | 172 ± 4 (1.2) |
| Control+ | 1319 ± 41 a | 1696 ± 41 b | 1875 ± 62 a | 1623 ± 48 b |
3. Experimental
3.1. Plant Material and Extraction
3.1.1. Isolation of 7,8,3'-trihydroxy-4'-methoxyisoflavone
3.1.2. Isoflavone Solubilization
3.2. Pharmacological Assays
3.2.1. Venom and Purification of BthTX-I
3.2.2. Animals
3.2.3. Mouse Phrenic Nerve-Diaphragm Muscle (PND) Preparation
3.2.4. Quantitative Histological Analysis
3.2.5. In vitro Mutagenicity Assay
3.2.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Warrell, D.A. Snakebites in central and south america: Epidemiology, clinical features and clinical management. In The Venomous Reptiles of the Western Hemisphere; Campbell, J.A., Lamar, W.W., Eds.; Comstock Publishing Associates/Cornell University Press: Ithaca, NY, USA, 2004; Volume 2, pp. 709–761. [Google Scholar]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of the Western Hemisphere; Comstock Publishing Associates/Cornell University Press: Ithaca, NY, USA, 2004; p. 870. [Google Scholar]
- Milani Junior, R.; Jorge, M.T.; de Campos, F.P.; Martins, F.P.; Bousso, A.; Cardoso, J.L.C.; Ribeiro, L.A.; Fan, H.W.; França, F.O.S.; Sano-Martins, I.S.; et al. Snake bites by the jararacuçu (Bothrops jararacussu): Clinicopathological studies of 29 proven cases in São Paulo State, Brazil. Q. J. Med. 1997, 90, 323–334. [Google Scholar] [CrossRef]
- Kashima, S.; Robert, P.G.; Soares, A.M.; Astolfi-Filho, S.; Pereira, J.O.; Giuliati, S.; Faria Junior, M.; Xavier, M.A.S.; Fontes, M.R.M.; Giglio, J.R.; et al. Analysis of Bothrops jararacussu venomous gland transcriptome focusing on structural and functional aspects: I—gene expression profile of highly expressed phospholipases A2. Biochimie 2004, 86, 211–219. [Google Scholar] [CrossRef]
- Brazil, V. Do envenamento ophidico e seu tratamento. Collet. Trab. Inst. Butantan 1901, 1, 31–55. [Google Scholar]
- Vellard, J.A. Serpentes venenosas. In Terapêutica Clínica; Cardini, C., Beretervide, J.J., Eds.; Libreria y Editorial “El Ateneo”: Buenos Aires, Argentina, 1945; ' Volume 4, pp. 265–273. [Google Scholar]
- Alves, E. Medicina de Urgência, 3rd ed.; Livraria Atheneu: Rio de Janeiro, Brazil, 1956; p. 562. [Google Scholar]
- Teixeira, R. Forma grave do acidente por ofídios da sub família Crotalinae. Ann. Acad. Med. Bahia 1979, 2, 109–135. [Google Scholar]
- Rodrigues-Simioni, L.; Borgese, N.; Ceccarelli, B. The effects of Bothrops jararacussu venom and its components on frog nerve-muscle preparation. Neuroscience 1983, 10, 475–489. [Google Scholar] [CrossRef]
- Zamunér, S.R.; Cruz-Höfling, M.A.; Corrado, A.P.; Hyslop, S.; Rodrigues-Simioni, L. Comparison of the neurotoxic and myotoxic effects of Brazilian Bothrops venoms and their neutralization by commercial antivenom. Toxicon 2004, 44, 259–271. [Google Scholar] [CrossRef]
- Homsi-Brandeburgo, M.I.; Queiroz, L.S.; Santo-Neto, H.; Rodrigues-Simioni, L.; Giglio, J.R. Fractionation of Bothrops jararacussu snake venom: Partial chemical characterization and biological activity of bothropstoxin. Toxicon 1988, 26, 615–627. [Google Scholar] [CrossRef]
- Heluany, N.F.; Homsi-Brandeburgo, M.I.; Giglio, J.R.; Prado-Franceschi, J.; Rodrigues-Simioni, L. Effects induced by bothropstoxin, a component from Bothrops jararacussu snake venom, on mouse and chick muscle preparations. Toxicon 1992, 30, 1203–1210. [Google Scholar] [CrossRef]
- Andrião-Escarso, S.H.; Soares, A.M.; Rodrigues, V.M.; Ângulo, Y.; Díaz, C.; Lomonte, B.; Gutiérrez, J.M.; Giglio, J.R. Myotoxic phospholipases A2 in Bothrops snake venoms: Effects of chemical modifications on the enzymatic and pharmacological properties of bothropstoxins from Bothrops jararacussu. Biochimie 2000, 82, 755–763. [Google Scholar] [CrossRef]
- Bonfin, V.L.; Toyama, M.H.; Novello, J.C.; Hyslop, S.; Oliveira, C.R.; Rodrigues-Simioni, L.; Marangoni, S. Isolation and enzymatic characterization of a basic phospholipase A2 from Bothrops jararacussu snake venom. J. Protein Chem. 2001, 20, 239–245. [Google Scholar] [CrossRef]
- Oshima-Franco, Y.; Leite, G.B.; Dal Belo, C.A.; Hyslop, S.; Prado-Franceschi, J.; Cintra, A.C.O.; Giglio, J.R.; Cruz-Höfling, M.A.; Rodrigues-Simioni, L. The presynaptic activity of bothropstoxin-I, a myotoxin from Bothrops jararacussu snake venom. Basic Clin. Pharmacol. Toxicol. 2004, 95, 175–182. [Google Scholar]
- Ponce-Soto, L.A.; Bonfim, V.L.; Rodrigues-Simioni, L.; Novello, J.C.; Marangoni, S. Determination of primary structure of two isoforms 6–1 and 6–2 PLA2 D49 from Bothrops jararacussu snake venom and neurotoxic characterization using in vitro neuromuscular preparation. Protein J. 2006, 25, 147–155. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B.; León, G.; Rucavado, A.; Chaves, F.; Angulo, Y. Trends in snakebite envenomation therapy: Scientific, technological and public health considerations. Curr. Pharm. Des. 2007, 13, 2935–2950. [Google Scholar] [CrossRef]
- Da Silva, N.M.V.; Arruda, E.Z.; Murakami, Y.L.B.; Moraes, R.A.M.; El-Kik, C.Z.; Tomaz, M.A.; Fernandes, F.F.A.; Oliveira, C.Z.; Soares, A.M.; Gigliio, J.R.; et al. Evaluation of three Brazilian antivenom ability to antagonize myonecrosis and hemorrhage induced by Bothrops snake venom in mouse model. Toxicon 2007, 50, 196–205. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B.; León, G.; Alape-Girón, A.; Flores-Díaz, M.; Sanz, L.; Angulo, Y.; Calvete, J.J. Snake venomics and antivenomics: Proteomic tools in the design and control of antivenoms for the treatment of snakebite envenoming. J. Proteomics 2009, 72, 165–182. [Google Scholar] [CrossRef]
- Mors, W.B.; Nascimento, M.C.; Pereira, B.M.; Pereira, N.A. Plant natural products active against snake bite—The molecular approach. Phytochemistry 2000, 55, 627–642. [Google Scholar] [CrossRef]
- Melo, R.F.; Farrapo, N.M.; Rocha Junior, D.S.; Silva, M.G.; Cogo, J.C.; Dal Belo, C.A.; Rodrigues Simioni, L.; Groppo, F.C.; Oshima-Franco, Y. Antiophidian mechanisms of medicinal plants. In Flavonoids: Biosynthesis, Biological Effects and Dietary Sources; Keller, R.B., Ed.; Nova Science: New York, NY, USA, 2009; pp. 249–262. [Google Scholar]
- Melo, P.A.; Nascimento, M.C.; Mors, W.B.; Suarez-Kurtz, G. Inhibition of the myotoxic and hemorrhagic activities of crotalid venoms by Eclipta prostrata (Asteraceae) extracts and constituents. Toxicon 1994, 32, 595–603. [Google Scholar] [CrossRef]
- Oshima-Franco, Y.; Alves, C.M.V.; Andréo Filho, N.; Gerenutti, M.; Cintra, A.C.O.; Leite, G.B.; Rodrigues-Simioni, L.; Silva, M.G. Neutralization of the neuromuscular activity of bothropstoxin-I, a myotoxin from Bothrops jararacussu snake venom, by a hydroalcoholic extract of Casearia sylvestris sw (guaçatonga). J. Venom. Anim. Toxins Incl. Trop. Dis. 2005, 11, 465–478. [Google Scholar]
- Cavalcante, W.L.; Campos, T.O.; Dal Pai-Silva, M.; Pereira, P.S.; Oliveira, C.Z.; Soares, A.M.; Gallacci, M. Neutralization of snake venom phospholipase A2 toxins by aqueous extract of Casearia sylvestris (Flacourtiaceae) in mouse neuromuscular preparation. J. Ethnopharmacol. 2007, 112, 490–497. [Google Scholar] [CrossRef]
- Cintra-Francischinelli, M.; Silva, M.G.; Andréo-Filho, N.; Gerenutti, M.; Cintra, A.C.O.; Giglio, J.R.; Leite, G.B.; Cruz-Höfling, M.A.; Rodrigues-Simioni, L.; Oshima-Franco, Y. Antibothropic action of Casearia sylvestris Sw. (Flacourtiaceae) extracts. Phytother. Res. 2008, 22, 784–790. [Google Scholar] [CrossRef]
- Camargo, T.M.; Nazato, V.S.; Silva, M.G.; Cogo, J.C.; Groppo, F.C.; Oshima-Franco, Y. Bothrops jararacussu venom-induced neuromuscular blockade inhibited by Casearia gossypiosperma Briquet hydroalcoholic extract. J. Venom. Anim. Toxins Incl. Trop. Dis. 2010, 16, 432–441. [Google Scholar]
- Farrapo, N.M.; Silva, G.A.A.; Costa, K.N.; Silva, M.G.; Cogo, J.C.; Dal Belo, C.A.; Santos, M.G.; Groppo, F.C.; Oshima-Franco, Y. Inhibition of Bothrops jararacussu venom activities by Plathymenia reticulata Benth extracts. J. Venom Res. 2010, 2, 52–58. [Google Scholar]
- Oshima-Franco, Y.; Rosa, L.J.R.; Silva, G.A.A.; Amaral Filho, J.; Silva, M.G.; Lopes, P.S.; Cogo, J.C.; Cintra, A.C.O.; da Cruz-Höfling, M.A. Antibothropic action of Camellia sinensis extract against the neuromuscular blockade by Bothrops jararacussu snake venom and its main toxin, bothropstoxin-I. In Pharmacology; Gallelli, L., Ed.; Intechopen: Croatia, Croatia, 2012; pp. 469–489. [Google Scholar]
- Tribuiani, N.; da Silva, A.M.; Ferraz, M.C.; Silva, M.G.; Bentes, A.P.G.; Graziano, T.S.; dos Santos, M.G.; Cogo, J.C.; Varanda, E.A.; Groppo, F.C.; et al. Vellozia flavicans Mart. ex Schult. hydroalcoholic extract inhibits the neuromuscular blockade induced by Bothrops jararacussu venom. BMC Complement. Altern. Med. 2014, 14, 48. [Google Scholar] [CrossRef]
- Veronese, E.L.; Esmeraldino, L.E.; Trombone, A.P.; Santana, A.E.; Bechara, G.H.; Kettelhut, I.; Cintra, A.C.; Giglio, J.R.; Sampaio, S.V. Inhibition of the myotoxic activity of Bothrops jararacussu venom and its two major myotoxins, BthTX-I and BthTX-II, by the aqueous extract of Tabernaemontana catharinensis A. DC. (Apocynaceae). Phytomedicine 2005, 12, 123–130. [Google Scholar] [CrossRef]
- Nazato, V.S.; Rubem-Mauro, L.; Vieira, N.A.G.; Rocha-Junior, D.S.; Silva, M.G.; Lopes, P.S.; Dal-Belo, C.A.; Cogo, J.C.; Santos, M.G.; Cruz-Höfling, M.A.; et al. In vitro antiophidian properties of Dipteryx alata Vogel bark extracts. Molecules 2010, 15, 5956–5970. [Google Scholar] [CrossRef]
- Puebla, P.; Oshima-Franco, Y.; Franco, L.M.; dos Santos, M.G.; da Silva, R.V.; Rubem-Mauro, L.; San Feliciano, A. Chemical constituents of the bark of Dipteryx alata Vogel, an active species against Bothrops jararacussu venom. Molecules 2010, 15, 8193–8204. [Google Scholar] [CrossRef]
- Ferraz, M.C.; Parrilha, L.A.C.; Moraes, M.S.D.; Amaral Filho, J.; Cogo, J.C.; dos Santos, M.G.; Franco, L.M.; Groppo, F.C.; Puebla, P.; San Feliciano, A.; et al. The effect of lupane triterpenoids (Dipteryx alata Vogel) in the in vitro neuromuscular blockade and myotoxicity of two snake venoms. Curr. Org. Chem. 2012, 16, 2717–2723. [Google Scholar] [CrossRef]
- Bezuidenhout, S.C.; Bezuidenhout, B.C.B.; Ferreira, D. α-Hydroxydihydrochalcones and related 1,3-diarylpropan-2-ones from Xanthocercis zambesiaca. Phytochemistry 1988, 27, 2329–2334. [Google Scholar] [CrossRef]
- Dewick, P.M. Isoflavonoids. In The Flavonoids Advances in Research Since 1986; Harborne, J.B., Ed.; Chapman & Hall/CRC: London, UK, 1994; pp. 117–386. [Google Scholar]
- Lampe, J.W. Isoflavonoid and lignan phytoestrogens as dietary biomarkers. J. Nutr. 2003, 133 Suppl., 956s–964s. [Google Scholar]
- Correia-de-Sá, P.; Noronha-Matos, J.B.; Ferreirinha, F.; Marques, P.; Soares, A.M.; Carvalho, C.; Cavalcante, W.L.; Gallacci, M. Bothropstoxin-Ireduces evoked acetylcholine release from rat motor nerve terminals: Radiochemical and real-time video-microscopy studies. Toxicon 2013, 61, 16–25. [Google Scholar] [CrossRef]
- Queiroz, L.S.; Santo Neto, H.; Rodrigues-Simioni, L.; Prado-Franceschi, J. Muscle necrosis and regeneration after envenomation by Bothrops jararacussu snake venom. Toxicon 1984, 22, 339–346. [Google Scholar] [CrossRef]
- Chaudhry, I.A.; Nitahara, K.; Nagashima, H.; Vizi, E.S. Neurochemical evidence that [Ca2+]o antagonizes the effect of neomycin on acetylcholine release from mouse hemidiaphragm preparation: An attempt to assess the margin to safety. Acta Anaesthesiol. Scand. 1995, 39, 494–497. [Google Scholar] [CrossRef]
- Harborne, J.B. Phytochemical Methods. A Guide to Modern Techniques of Plant Analysis, 3rd ed.; Chapman & Hall: London, UK, 1998. [Google Scholar]
- OECD Guideline for Testing of Chemicals. Bacterial Reverse Mutation Test. Adopted: 21st July 1997. Available online: http://www.oecd.org/chemicalsafety/risk-assessment/1948418.pdf (accessed on 27 January 2014).
- Kaur, K.; Mathur, N.; Bhatnagar, P. Comparative study of usage of microbial strains for monitoring waste water treatment plants. Univers. J. Environ. Res. Technol. 2012, 2, 26–37. [Google Scholar]
- Vedmaurthy, R.B.; Padmanabhan, S.; Vijayan, M.; Jamal, Z.A.; Kunjumman, J.; Narayanan, M.L. Compatibility of different solvents with Salmonella typhimurium mutant strains in bacterial reverse mutation assay. Int. J. Pharm. Pharm. Sci. 2012, 4, 283–284. [Google Scholar]
- Swartz, C.; Parks, N.; Schaaper, R.M.; Demarini, M. General enhancement of mutagenic potency of various mutagens due to deleted genes in the uvrB strains TA 98 and TA 100 of Salmonella compared with strains containing only a point mutation in uvr B. U.S. Environmental Protection Agency. Presented at The 9th International Conference on Environmental Mutagens, and 36th Annual Meeting of the International Conference on Environmental Mutagen Society. San Francisco, CA, USA, 3–8 September 2005; Available online: http://www.sciencedirect.com/science/journal/00275107/577/supp/S (accessed on 28 January 2014).
- Mortelmans, K.; Zeiger, E. The Ames Salmonella/microsome mutagenicity assay. Mutat. Res. 2009, 455, 29–36. [Google Scholar] [CrossRef]
- Esteves-Pedro, N.M.; Borim, T.; Nazato, V.S.; Silva, M.G.; Lopes, P.S.; dos Santos, M.G.; Dal Belo, C.A.; Cardoso, C.R.P.; Varanda, E.A.; Groppo, F.C.; et al. In vitro and in vivo safety evaluation of Dipteryx alata Vogel extract. BMC Complement. Altern. Med. 2012, 12, 9. [Google Scholar] [CrossRef]
- Cintra-Francischinelli, M.; Silva, M.G.; Andreo-Filho, N.; Cintra, A.C.O.; Leite, G.B.; Cruz-Höfling, M.A.; Rodrigues-Simioni, L.; Oshima-Franco, Y. Effects of commonly used solubilizing agents on a model nerve-muscle synapse. Lat. Am. J. Pharm. 2008, 27, 721–726. [Google Scholar]
- Oshima, M.; Leite, G.B.; Rostelato-Ferreira, S.; Cruz-Höfling, M.A.; Rodrigues-Simioni, L.; Oshima-Franco, Y. Insights of the effects of polyethylene glycol 400 on mammalian and avian nerve terminals. Muscle Nerve 2010, 41, 540–546. [Google Scholar] [CrossRef]
- Bülbring, E. Observation on the isolated phrenic nerve diaphragm preparation of the rat. Br. J. Pharmacol. 1946, 1, 38–61. [Google Scholar]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Bernstein, L.; Kaldor, J.; McCann, J.; Pike, M.C. An empirical approach to the statistical analysis of mutagenesis data from the Salmonella test. Mutat. Res. 1982, 97, 267–281. [Google Scholar]
- Varella, S.D.; Pozetti, G.L.; Vilegas, W.; Varanda, E.A. Mutagenic activity of sweepings and pigments from a household-wax factory assayed with Salmonella typhimurium. Food Chem. Toxicol. 2004, 42, 2029–2035. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ferraz, M.C.; Yoshida, E.H.; Tavares, R.V.S.; Cogo, J.C.; Cintra, A.C.O.; Dal Belo, C.A.; Franco, L.M.; Dos Santos, M.G.; Resende, F.A.; Varanda, E.A.; et al. An Isoflavone from Dipteryx alata Vogel is Active against the in Vitro Neuromuscular Paralysis of Bothrops jararacussu Snake Venom and Bothropstoxin I, and Prevents Venom-Induced Myonecrosis. Molecules 2014, 19, 5790-5805. https://doi.org/10.3390/molecules19055790
Ferraz MC, Yoshida EH, Tavares RVS, Cogo JC, Cintra ACO, Dal Belo CA, Franco LM, Dos Santos MG, Resende FA, Varanda EA, et al. An Isoflavone from Dipteryx alata Vogel is Active against the in Vitro Neuromuscular Paralysis of Bothrops jararacussu Snake Venom and Bothropstoxin I, and Prevents Venom-Induced Myonecrosis. Molecules. 2014; 19(5):5790-5805. https://doi.org/10.3390/molecules19055790
Chicago/Turabian StyleFerraz, Miriéle C., Edson H. Yoshida, Renata V.S. Tavares, José C. Cogo, Adélia C.O. Cintra, Cháriston A. Dal Belo, Luiz M. Franco, Márcio G. Dos Santos, Flávia A. Resende, Eliana A. Varanda, and et al. 2014. "An Isoflavone from Dipteryx alata Vogel is Active against the in Vitro Neuromuscular Paralysis of Bothrops jararacussu Snake Venom and Bothropstoxin I, and Prevents Venom-Induced Myonecrosis" Molecules 19, no. 5: 5790-5805. https://doi.org/10.3390/molecules19055790