Hydrogen Peroxide and Nitric Oxide are Involved in Salicylic Acid-Induced Salvianolic Acid B Production in Salvia miltiorrhiza Cell Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
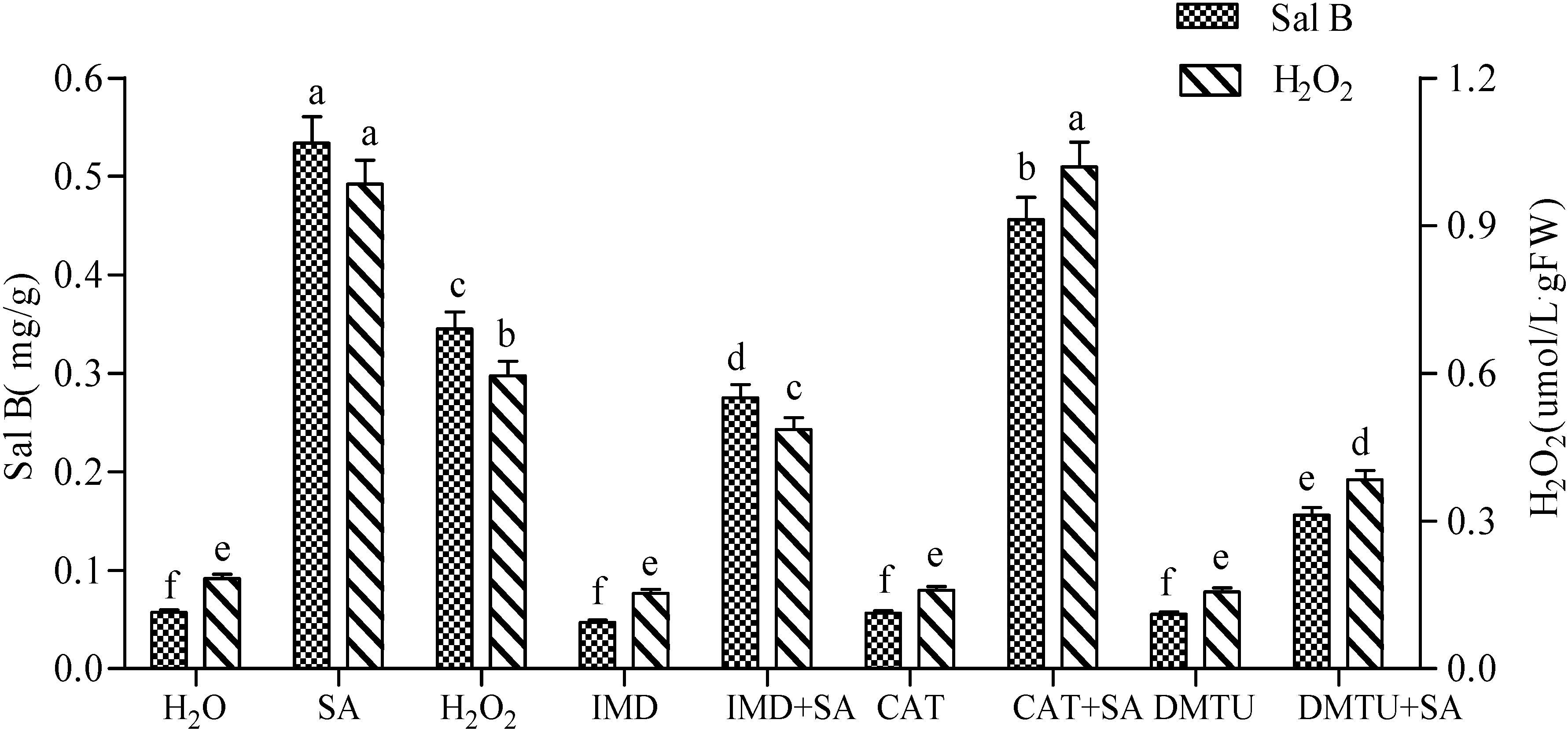
2.1. H2O2 Production in SA-Induced Cells and Its Effect on Sal B Accumulation


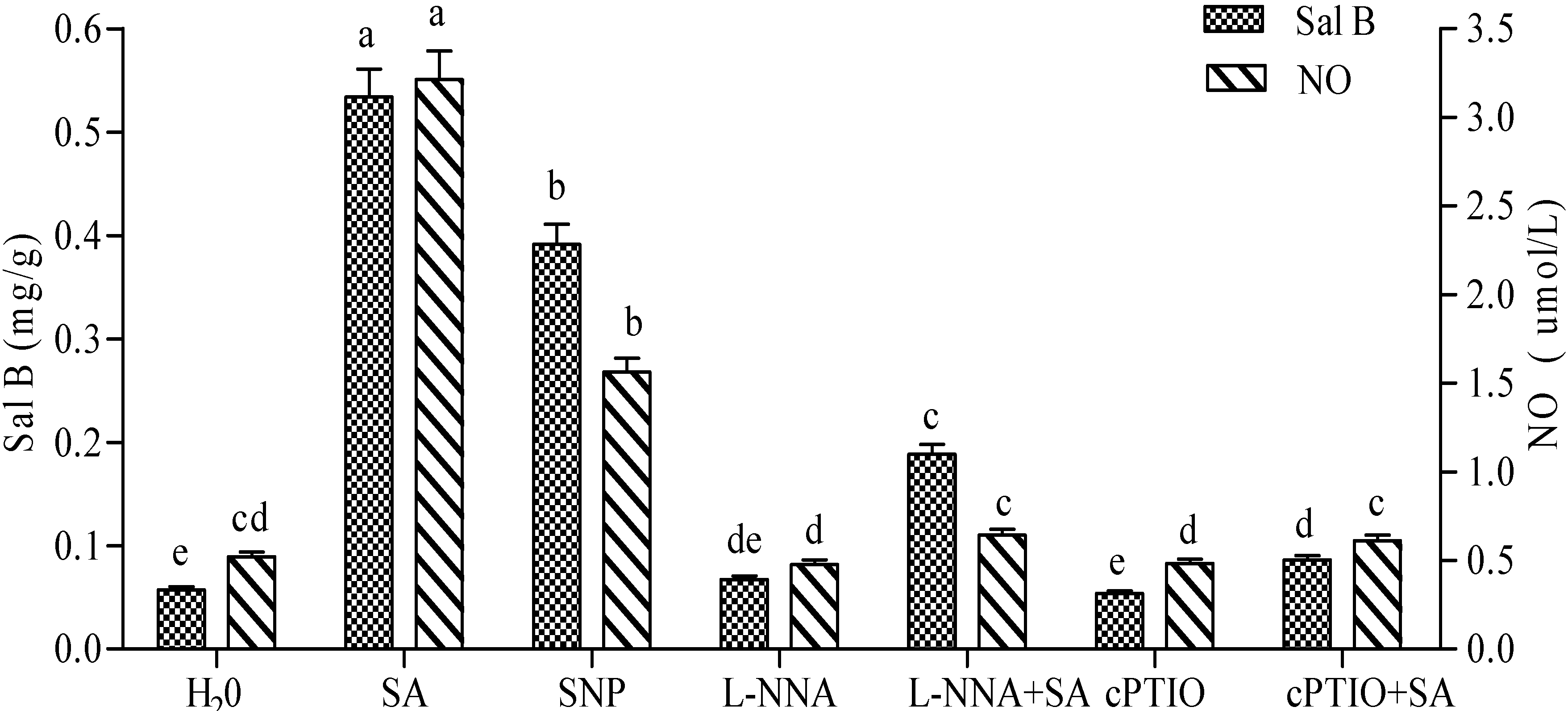
2.2. NO Production in SA-Induced Cells and Its Effect on Sal B Accumulation
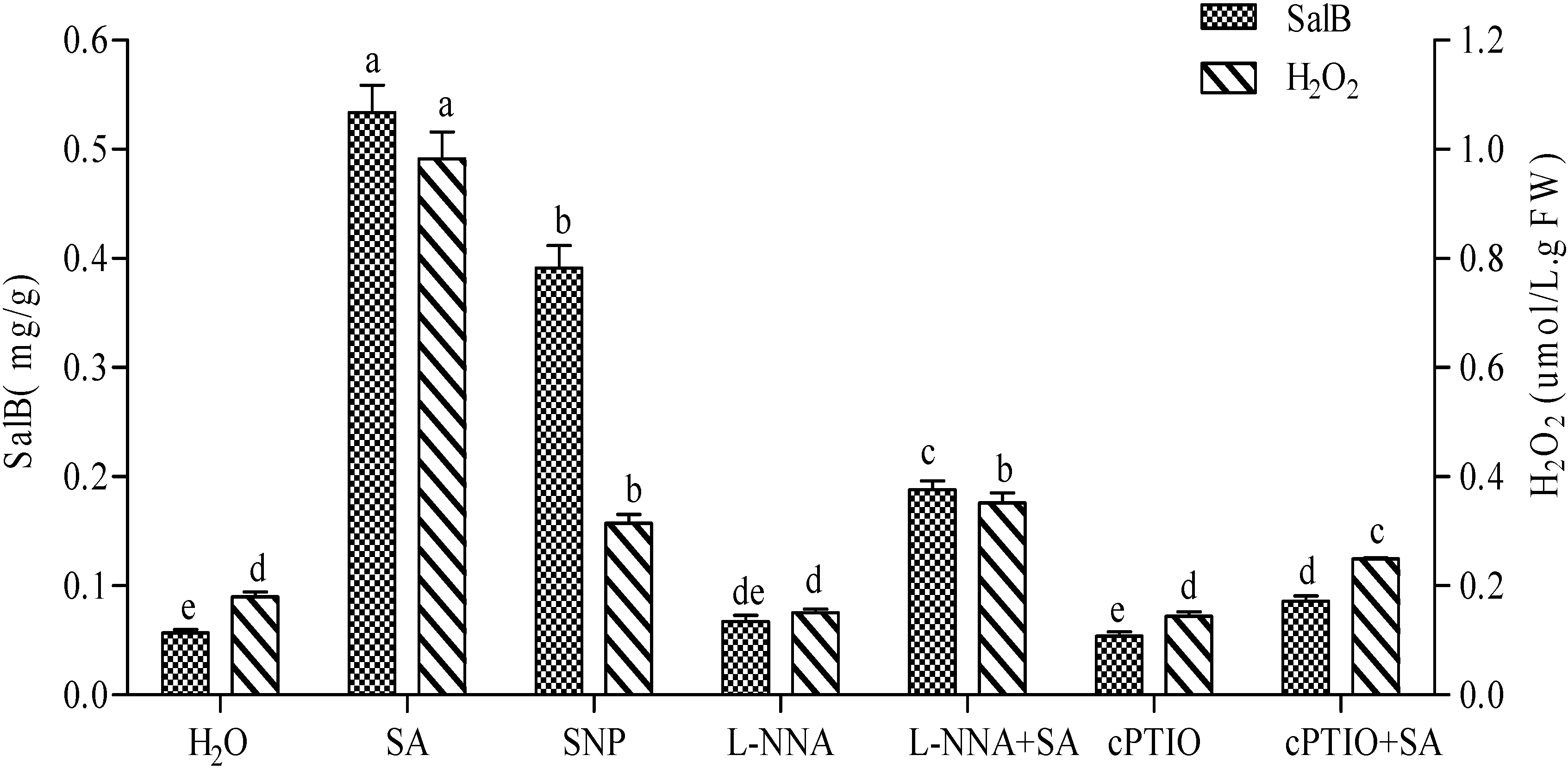
2.3. Production of NO Elicited by SA and Its Effect on the Accumulation of H2O2 and Sal B

2.4. Production of H2O2 Elicited by SA and Its Effect on the Accumulation of NO and Sal B

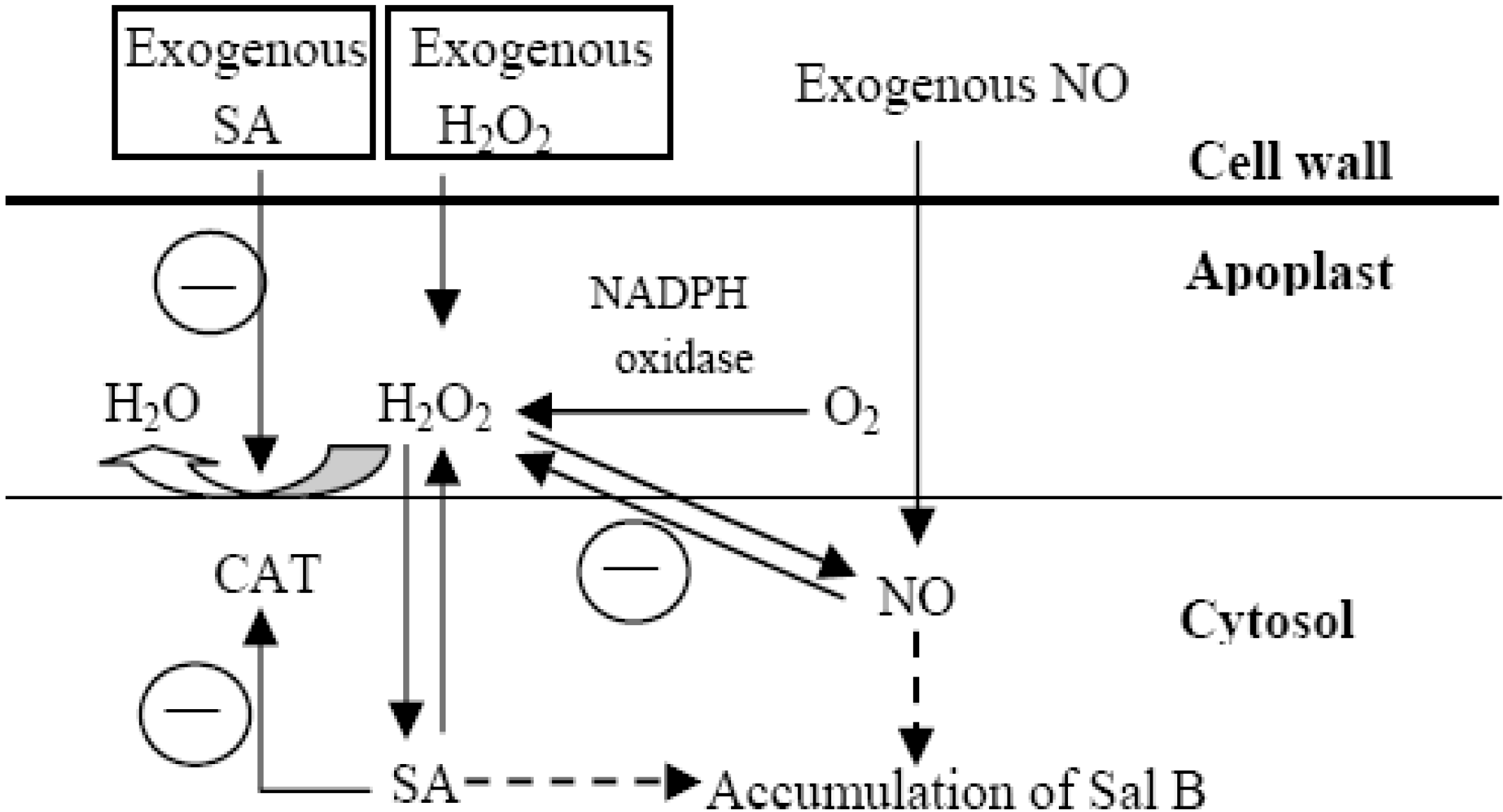
2.5. Both H2O2 and NO Mediate the SA-Elicited Sal B Production

2.6. Discussion

3. Experimental Section
3.1. Cell Culture and Treatment
3.2. Determination of H2O2 and NO
3.3. Sal B Extraction and HPLC Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chandra, S.; Chandra, R. Engineering secondary metabolite production in hairy roots. Phytochem. Rev. 2011, 10, 371–395. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar]
- Zhang, C.; Yan, Q.; Cheuk, W.; Wu, J. Enhancement of tanshinone production in Salvia miltiorrhiza hairy root culture by Ag+ elicitation and nutrient feeding. Planta Med. 2004, 70, 147–151. [Google Scholar]
- Vicente, M.R.S.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Tancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
- Chen, Z.; Silva, H.; Klessig, D.F. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 1993, 262, 1883–1886. [Google Scholar]
- Van Camp, W.; van Montagu, M.; Inzé, D. H2O2 and NO: Redox signals in disease resistance. Trend Plant. Sci. 1998, 3, 330–334. [Google Scholar] [CrossRef]
- Chamnongpol, S.; Willekens, H.; Moeder, W.; Langebartels, C.; Sandermann, H., Jr.; van Montagu, M.; Inze, D.; van Camp, W. Defense activation and enhanced pathogen tolerance induced by H2O2 in transgenic plants. Proc. Natl. Acad. Sci. USA 1998, 95, 5818–5823. [Google Scholar] [CrossRef]
- Delledonne, M.; Xia, Y.J.; Dixon, R.A.; Lamb, C. Nitric oxide as a signal in plant disease resistance. Nature 1998, 394, 585–588. [Google Scholar]
- Durner, J.; Shah, J.; Klessig, D.F. Salicylic acid and disease resistance in plants. Trend Plant Sci. 1997, 2, 266–274. [Google Scholar] [CrossRef]
- National Committee of China Pharmacopoeia. China Pharmacopoeia, Chemical Industry Press: Beijing, China, 2010; 52.
- Shu, Z.; Wang, Z.; Mu, X.; Liang, Z.; Guo, H. A dominant gene for male sterility in Salvia miltiorrhiza Bunge. PLoS One 2012, 7, e50903. [Google Scholar]
- Dong, J.; Wan, G.; Liang, Z. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture. J. Biotechnol. 2010, 148, 99–104. [Google Scholar] [CrossRef]
- Zhao, G.R.; Zhang, H.M.; Ye, T.X.; Xiang, Z.J.; Yuan, Y.J.; Guo, Z.X.; Zhao, L.B. Characterization of the radical scavenging and antioxidant activities of danshensu and salvianolic acid B. Food Chem. Toxicol. 2008, 46, 73–81. [Google Scholar]
- Sticher, L.; Mauch-Mani, B.; Métreaux, J.P. Systemic acquired resistance. Ann. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar]
- Taguchi, G.; Yazawa, T.; Hayashida, N.; Okazaki, M. Molecular cloning and heterologous expression of novel glucosyltransferases from tobacco cultured cells that have broad substrates specificity and are induced by salicylic acid and auxin. Eur. J. Biochem. 2001, 268, 4086–4094. [Google Scholar]
- Alvarez, M.E.; Pennell, R.I.; Meijer, P.J.; Ishikawa, A.; Dixon, R.A.; Lamb, C. Reactive oxygen intermediates mediate a systemic signal network in the establishment of plant immunity. Cell 1998, 92, 773–784. [Google Scholar]
- Leon, J.; Lawton, M.A.; Raskin, I. Hydrogen peroxide stimulates salicylic acid biosynthesis in tobacco. Plant Physiol. 1995, 108, 1673–1678. [Google Scholar]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P.; Murr, D.P.; Watkins, C.B. Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2-metabolizing enzymes. Plant Physiol. 1997, 115, 137–149. [Google Scholar]
- Shrasu, K.; Nakajima, H.; Rajasekhar, V.K.; Dixon, R.A.; Lamb, C. Salicylic acid potentiates an agonist-dependent gain control that amplifies pathogen signals in the activation of defense mechanisms. Plant Cell 1997, 9, 261–270. [Google Scholar]
- Durner, J.; Wendehenne, D.; Klessig, D.F. Defense gene induction in tobacco by nitric oxide, cyclic GMP and cyclic ADP ribose. Proc. Natl. Acad. Sci. USA 1998, 95, 10328–10333. [Google Scholar]
- Levine, A.; Tenhaken, R.; Dixon, R.; Lamb, C. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. [Google Scholar] [CrossRef]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bezier, A.; Lambert, B.; Joubert, J.M.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microb. Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef]
- Desikan, R.; Griffiths, R.; Hancock, J.T.; Neill, S. A new role for an old enzyme: Nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2002, 99, 16314–16318. [Google Scholar] [CrossRef]
- Garcia-Mata, L.; Lamattina, L. Nitric oxide and abscisic acid cross-talk in guard cells. Plant Physiol. 2002, 128, 790–792. [Google Scholar]
- Lu, S.; Su, W.; Li, H.; Guo, Z. Abscisic acid improves drought tolerance of triploid bermudagrass and involves H2O2- and NO-induced antioxidant enzyme activities. Plant Physiol. Biochem. 2009, 47, 132–138. [Google Scholar] [CrossRef]
- Wang, Y.; Ries, A.; Wu, K.; Yang, A.; Crawford, N.M. The Arabidopsis prohibition gene PHB3 functions in nitric oxide-mediated response and in hydrogen peroxide-induced nitric oxide accumulation. Plant Cell 2010, 22, 249–259. [Google Scholar] [CrossRef]
- Poderoso, J.J.; Peralta, J.G.; Lisdero, C.L.; Carreras, M.C.; Radisic, M.; Schöpfer, F.; Cadenas, E.; Boveris, A. Nitric oxide regulates oxygen uptake and hydrogen proxide release by the isolated beating rat heart. Am. J. Physiol. 1998, 274, C112–C119. [Google Scholar]
- Hu, X.; Neill, S.; Cai, W.; Tang, Z. Hydrogen peroxide and jasmonic acid medicate oligogalacturonic acid-induced saponin accumulation in suspension-cultured cells of Panax ginseng. Physiol. Plant 2003, 118, 414–421. [Google Scholar] [CrossRef]
- Dong, J.; Liu, Y.; Liang, Z.; Wang, W. Investigation on ultrasound-assisted extraction of salvianolic acid B from Salvia miltiorrhiza root. Ultrason. Sonochem. 2010, 17, 61–65. [Google Scholar] [CrossRef]
- Sample Availability: Please connect the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Guo, H.; Dang, X.; Dong, J. Hydrogen Peroxide and Nitric Oxide are Involved in Salicylic Acid-Induced Salvianolic Acid B Production in Salvia miltiorrhiza Cell Cultures. Molecules 2014, 19, 5913-5924. https://doi.org/10.3390/molecules19055913
Guo H, Dang X, Dong J. Hydrogen Peroxide and Nitric Oxide are Involved in Salicylic Acid-Induced Salvianolic Acid B Production in Salvia miltiorrhiza Cell Cultures. Molecules. 2014; 19(5):5913-5924. https://doi.org/10.3390/molecules19055913
Chicago/Turabian StyleGuo, Hongbo, Xiaolin Dang, and Juane Dong. 2014. "Hydrogen Peroxide and Nitric Oxide are Involved in Salicylic Acid-Induced Salvianolic Acid B Production in Salvia miltiorrhiza Cell Cultures" Molecules 19, no. 5: 5913-5924. https://doi.org/10.3390/molecules19055913