Enhancement of Phenolics, Resveratrol and Antioxidant Activity by Nitrogen Enrichment in Cell Suspension Culture of Vitis vinifera

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
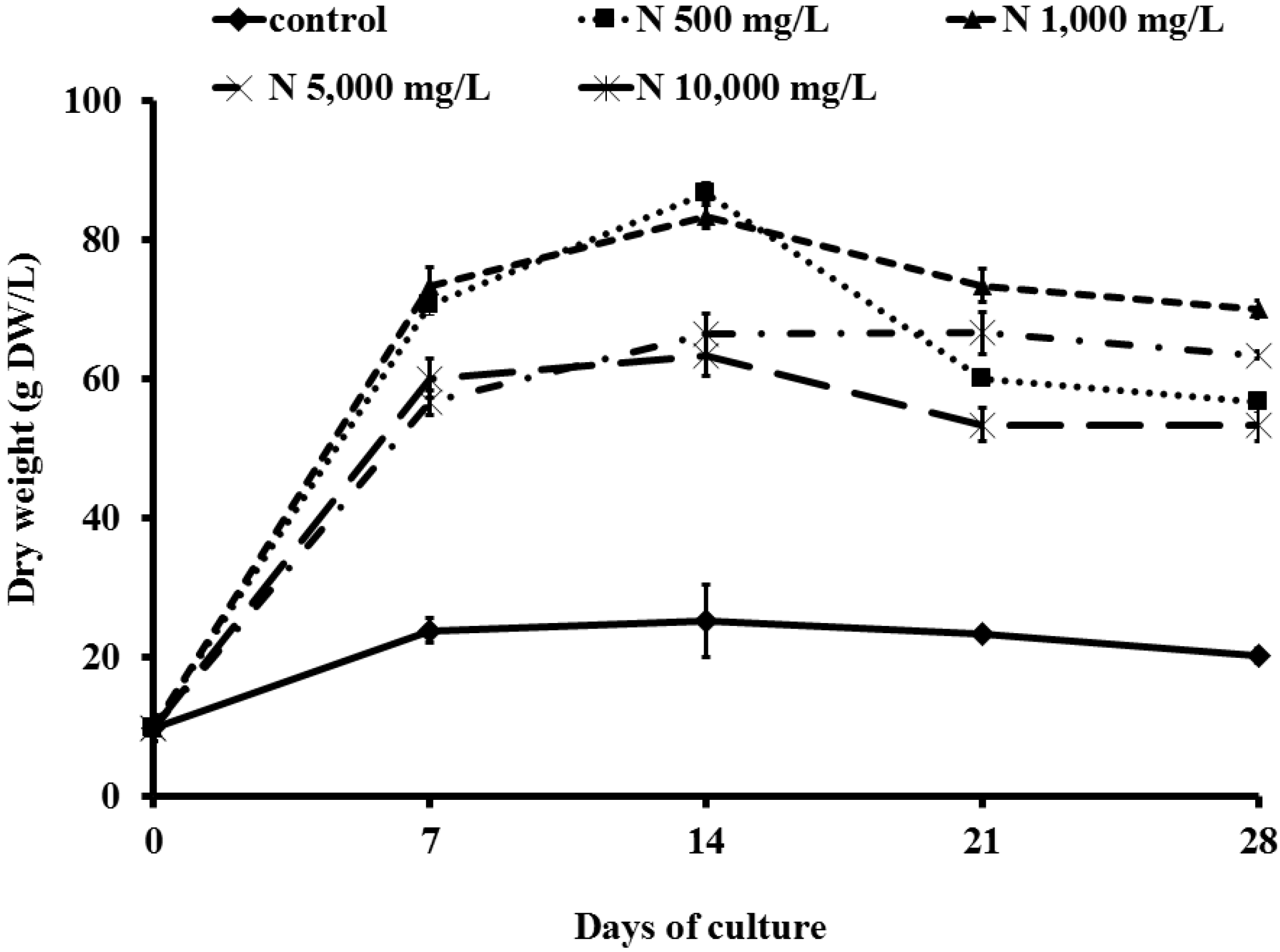
2.1. Effect of Ammonium Nitrate (NH4NO3) on Cell Growth

2.2. Effect of Ammonium Nitrate (NH4NO3) on Phenolic Content

2.3. Effect of Ammonium Nitrate (NH4NO3) on Resveratrol Contents


2.4. Antioxidant Activities
2.4.1. Determination of Free Radical Scavenging Activity Using 2, 2-Diphenyl-1-picrylhydrazyl (DPPH●) Assay

2.4.2. Determination of Free Radical Scavenging Activity Using the 2, 2'-Azinobis-(3-ethylbenzo-thiazoline-6-sulfonic acid) (ABTS•+) Assay
3. Experimental
3.1. Plant Materials
3.2. Callus Induction and Subculture
3.3. Cell Suspension Culture
3.4. Determination of Cell Biomass
3.5. Determination of the Total Phenolic and Resveratrol Contents
3.5.1. Determination of Total Phenolics
3.5.2. Determination of Trans-Resveratrol
3.6. Measurement of Antioxidant Activities
3.6.1. 1,1-Diphenyl-2-picrylhydrazyl (DPPH●) Assay
3.6.2. 2,2-Azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS•+) Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cherif, A.O. Phytochemicals components as bioactive foods. In Bioactive Compounds in Phytomedicine; Rasooli, I., Ed.; InTech Europe: Rijeka, Croatia, 2012; pp. 113–124. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- He, S.; Yan, X. From resveratrol to its derivatives: New sources of natural antioxidant. Curr. Med. Chem. 2013, 20, 1–13. [Google Scholar]
- Ferri, M.; Dipalo, S.C.F.; Bagni, N.; Tassoni, A. Chitosan elicits mono-glucosylated stilbene production and release in fed-batch bioreactor cultures of grape cells. Food Chem. 2011, 124, 1473–1479. [Google Scholar] [CrossRef]
- Narayan, M.S.; Venkataraman, L.V. Effect of sugar and nitrogen on the production of anthocyanin in cultured carrot (Daucus carota) cells. J. Food Sci. 2002, 67, 84–86. [Google Scholar] [CrossRef]
- Nagella, P.; Murthy, H.N. Establishment of cell suspension cultures of Withania somnifera for the production of withanolide A. Bioresour. Technol. 2010, 101, 6735–6739. [Google Scholar] [CrossRef]
- Praveen, N.; Murthy, H.N.; Chung, I.M. Improvement of growth and gymnemic acid production by altering the macro elements concentration and nitrogen source supply in cell suspension cultures of Gymnema sylvestre R. Br. Ind. Crop Prod. 2011, 33, 282–286. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, D.-E.; Lee, H.-S.; Kim, S.-K.; Lee, W.-S.; Kim, S.-H.; Kim, M.-W. Influence of auxins, cytokinins, and nitrogen on production of rutin from callus and adventitious roots of the white mulberry tree (Morus alba L.). Plant Cell Tiss. Org. 2010, 105, 9–19. [Google Scholar]
- Gueven, A.; Knorr, D. Isoflavonoid production by soy plant callus suspension culture. J. Food Eng. 2011, 103, 237–243. [Google Scholar] [CrossRef]
- Bensaddek, L.; Gillet, F.; Saucedo, J.E.N.; Fliniaux, M.-A. The effect of nitrate and ammonium concentrations on growth and alkaloid accumulation of Atropa belladonna hairy roots. J. Biotechnol. 2001, 85, 35–40. [Google Scholar] [CrossRef]
- Zhong, J.J.; Wang, S.J. Effects of nitrogen source on the production of ginseng saponin and polysaccharide by cell cultures of Panax quinquefolium. Process Biochem. 1998, 33, 671–675. [Google Scholar] [CrossRef]
- Amdoun, R.; Khelifi, L.; Khelifi-Slaoui, M.; Amroune, S.; Benyoussef, E.-H.; Thi, D.V.; Assaf-Ducrocq, C.; Gontier, E. Influence of minerals and elicitation on Datura stramonium L. tropane alkaloid production: Modelization of the in vitro biochemical response. Plant Sci. 2009, 177, 81–87. [Google Scholar] [CrossRef]
- Stewart, A.J.; Chapman, W.; Jenkins, G.I.; Graham, I.; Martin, T.; Crozier, A. The effect of nitrogen and phosphorous deficiency on flavonol accumulation in plant tissues. Plant Cell Environ. 2001, 24, 1189–1197. [Google Scholar] [CrossRef]
- Kovacik, J.; Klejdus, B.; Backor, M.; Repcak, M. Phenylalanine ammonialyase activity and phenolic compounds accumulation in nitrogen-deficient Matricaria chamomilla leaf rosettes. Plant Sci. 2007, 172, 393–399. [Google Scholar] [CrossRef]
- Scheible, W.-R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-wide reprogramming of primary and secondary metabolism, protein synthesis, cellular growth processes, and the regulatory infrastructure of Arabidopsis in response to nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef]
- Khunthajaroen, T.; Kerdchoechuen, O.; Laohakunjit, N. Resveratrol, catechin and antioxidant activity from ‘Pok Dum’ grape extracts. J. Agric. Sci. 2011, 42, 121–124. [Google Scholar]
- Sakuta, M.; Takagi, T.; Komamine, A. Effects of nitrogen source on betacyanin accumulation and growth in suspension cultures of Phytolacca americana. Physiol. Plantarum. 1987, 71, 459–463. [Google Scholar] [CrossRef]
- Cui, X.-H.; Murthy, H.N.; Wu, C.-H.; Paek, K.-Y. Adventitious root suspension cultures of Hypericum perforatum: Effect of nitrogen source on production of biomass and secondary metabolites. In Vitro Cell. Dev.-Pl. 2010, 46, 437–444. [Google Scholar] [CrossRef]
- Kovacik, J.; Backor, M. Changes of phenolic metabolism and oxidative status in nitrogen-deficient Matricaria chamomilla plants. Plant Soil 2007, 297, 255–265. [Google Scholar] [CrossRef]
- Giri, L.; Dhyania, P.; Rawata, S.; Bhatta, I.D.; Nandia, S.K.; Rawala, R.S.; Pandeb, V. In vitro production of phenolic compounds and antioxidant activity in callus suspension cultures of Habenaria edgeworthii: A rare Himalayan medicinal orchid. Ind. Crop Prod. 2012, 39, 1–6. [Google Scholar] [CrossRef]
- Teguo, P.W.; Fauconneau, B.; Deffieux, G.; Huguet, F.; Vercauteren, J.; Merillon, J.M. Isolation, identification, and antioxidant activity of three stilbene glucosides newly extracted from Vitis vinifera cell cultures. J. Nat. Prod. 1998, 61, 655–657. [Google Scholar] [CrossRef]
- Sample Availability: Samples of trans-resveratrol compounds from cell suspension culture of Vitis vinifera cv. Pok Dum are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sae-Lee, N.; Kerdchoechuen, O.; Laohakunjit, N. Enhancement of Phenolics, Resveratrol and Antioxidant Activity by Nitrogen Enrichment in Cell Suspension Culture of Vitis vinifera. Molecules 2014, 19, 7901-7912. https://doi.org/10.3390/molecules19067901
Sae-Lee N, Kerdchoechuen O, Laohakunjit N. Enhancement of Phenolics, Resveratrol and Antioxidant Activity by Nitrogen Enrichment in Cell Suspension Culture of Vitis vinifera. Molecules. 2014; 19(6):7901-7912. https://doi.org/10.3390/molecules19067901
Chicago/Turabian StyleSae-Lee, Napaporn, Orapin Kerdchoechuen, and Natta Laohakunjit. 2014. "Enhancement of Phenolics, Resveratrol and Antioxidant Activity by Nitrogen Enrichment in Cell Suspension Culture of Vitis vinifera" Molecules 19, no. 6: 7901-7912. https://doi.org/10.3390/molecules19067901