Evidence for the Involvement of Spinal Cord-Inhibitory and Cytokines-Modulatory Mechanisms in the Anti-Hyperalgesic Effect of Hecogenin Acetate, a Steroidal Sapogenin-Acetylated, in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

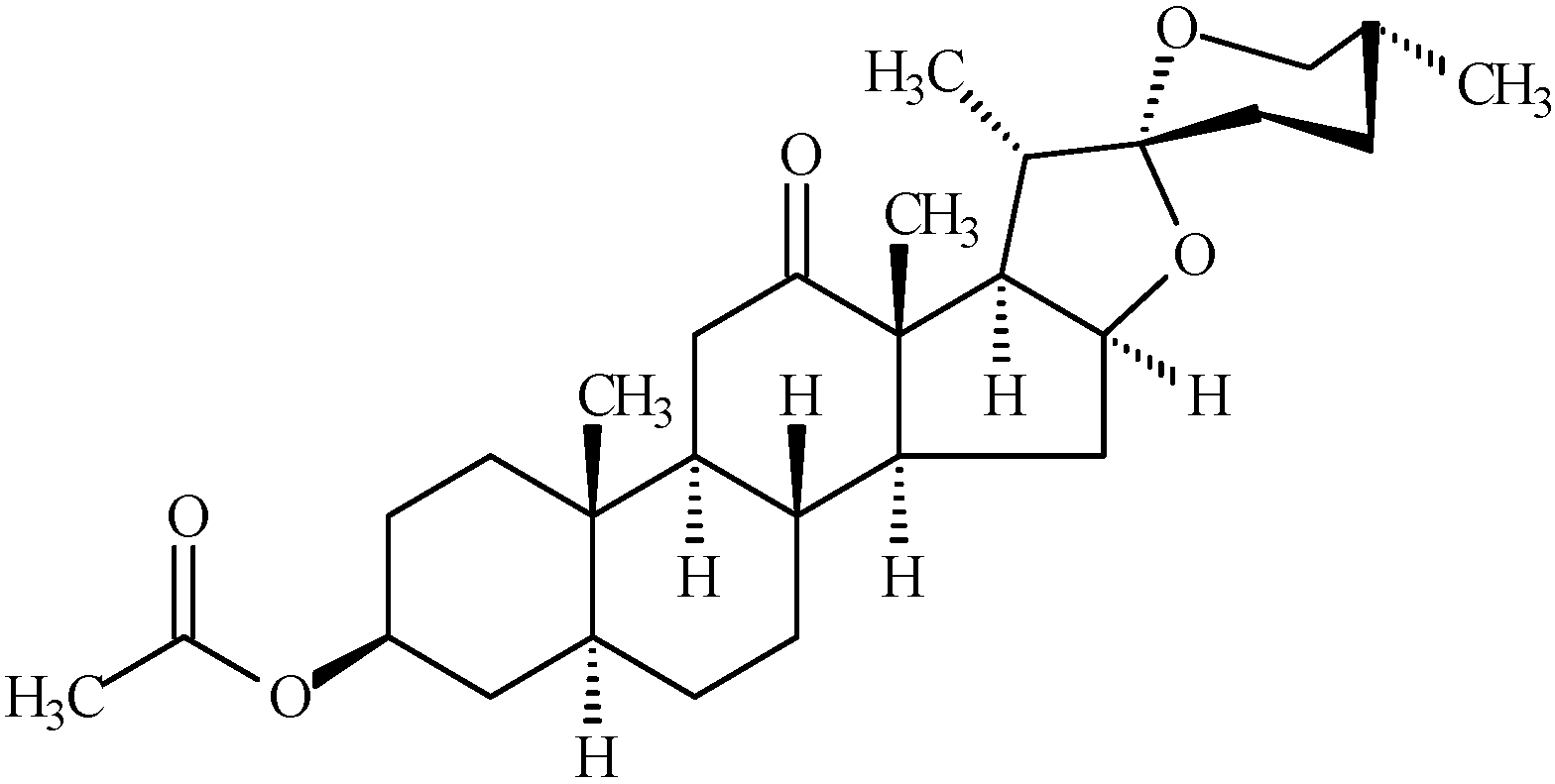
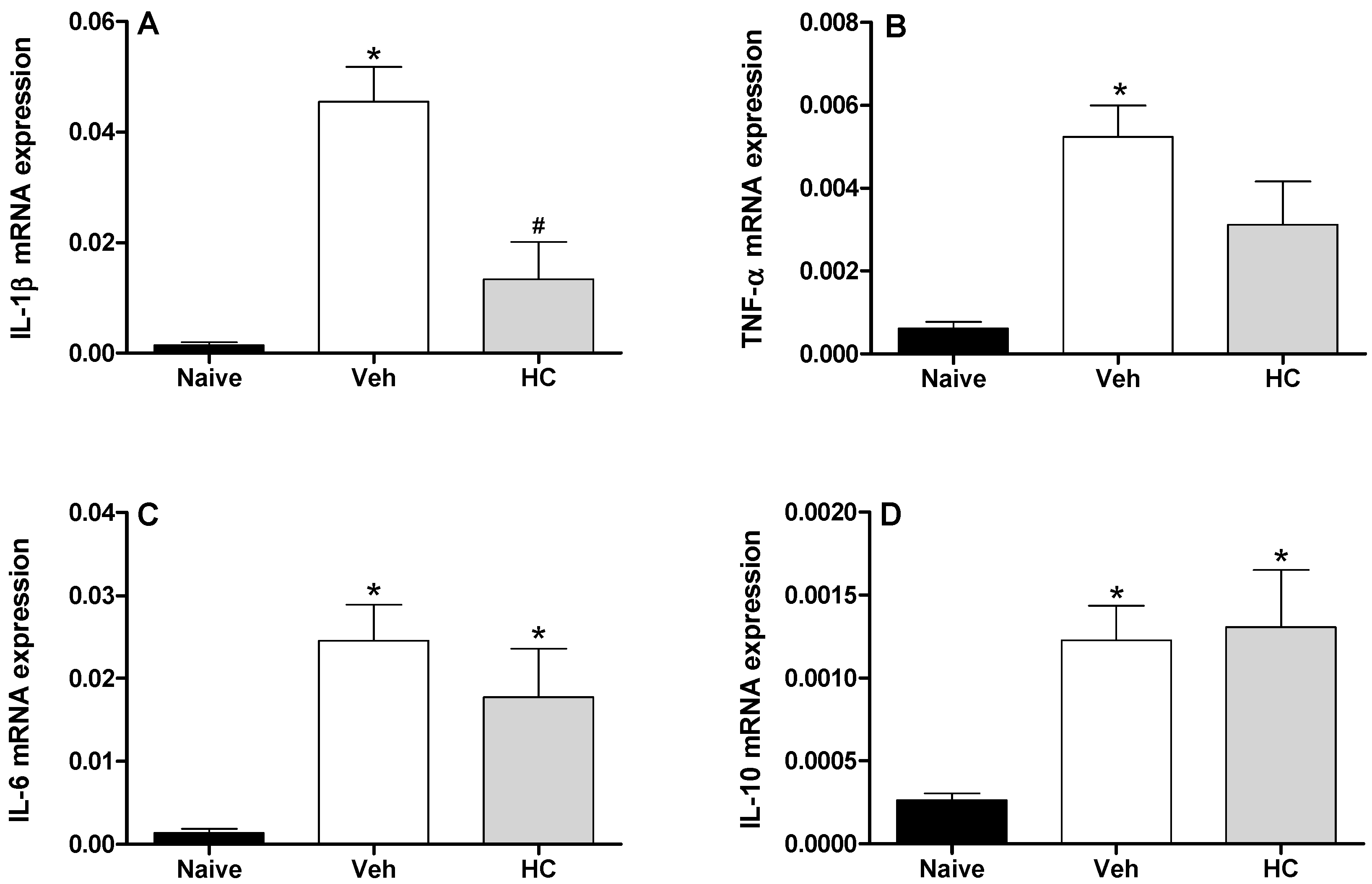
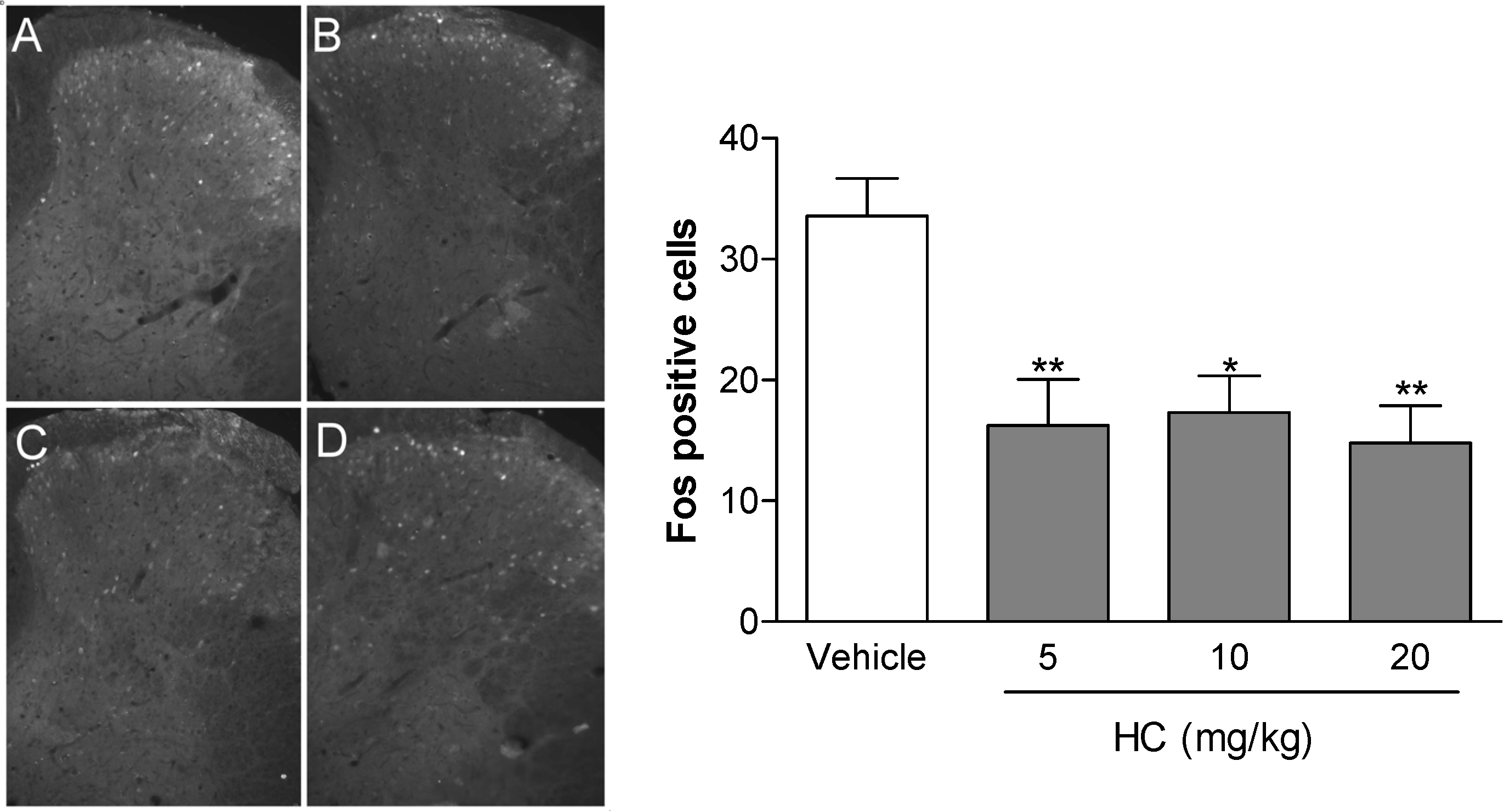
2. Results and Discussion




3. Experimental
3.1. Animals
3.2. Hyperalgesic Stimulus and Nociceptive Threshold Evaluation
3.3. Motor Function Assay (Rota-Rod Test)
3.4. Immunofluorescence
3.5. Acquisition and Analyses of Images
3.6. Real Time PCR
3.7. Drugs and Reagents
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kissin, I. The development of new analgesics over the past 50 years: A lack of real breakthrough drugs. Anesth. Analg. 2010, 110, 780–789. [Google Scholar] [CrossRef]
- Amaral, J.F.; Silva, M.I.G.; Neto, M.R.A.; Neto, P.F.T.; Moura, B.A.; Melo, C.T.V.; Araújo, F.L.O.; de Sousa, D.P.; Vasconcelos, P.F.; Vasconcelos, S.M.; et al. Antinociceptive Effect of the Monoterpene R-(+)-Limonene in Mice. Biol. Pharm. Bull. 2007, 30, 1217–1220. [Google Scholar] [CrossRef]
- Mendonça, J.M.D.; Lyra, D.P.; Rabelo, J.S.; Siqueira, J.S.; Balisa-Rocha, B.J.; Gimenes, F.R.E.; Bonjardim, L.R. Analysis and detection of dental prescribing errors at Primary Health Care Units in Brazil. Pharm. World Sci. 2010, 32, 30–35. [Google Scholar] [CrossRef]
- Li, J.W.H.; Vederas, J.C. Drug discovery and natural products: End of an era or an endless frontier? Science 325, 2009, 161–165. [Google Scholar]
- Schmidtko, A.; Lötsch, J.; Freynhagen, R.; Geisslinger, G. Ziconotide for treatment of severe chronic pain. Lancet 2010, 375, 1569–1577. [Google Scholar] [CrossRef]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Guimarães, A.G.; Quintans, J.S.S.; Quintans-Júnior, L.J. Monoterpenes with analgesic activity - a systematic review. Phytother. Res. 2013, 27, 1–15. [Google Scholar] [CrossRef]
- Guimarães, A.G.; Serafini, M.R.; Quintans-Júnior, L.J. Terpenes and derivatives as a new perspective for pain treatment: A patent review. Expert Opin. Ther. Pat. 2014, 24, 243–265. [Google Scholar]
- Eskander, J.; Lavaud, C.; Harakat, D. Steroidal saponins from the leaves of Agave macroacantha. Fitoterapia 2010, 81, 371–374. [Google Scholar] [CrossRef]
- Hocking, G.M. A Dictionary of Natural Products; Plexus Publishing Inc: Medford, OR, USA, 1997. [Google Scholar]
- Kostova, I.; Dinchev, D. Saponins in Tribulus terrestris - chemistry and bioactivity. Phytochem. Rev. 2005, 4, 111–137. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Sakai, T.; Koezuka, Y. Glycolipid derivatives as therapeutic agents. Expert Opin. Ther. Pat. 1999, 9, 917–930. [Google Scholar] [CrossRef]
- Ghoghari, A.M.; Rajani, M. Densitometric determination of hecogenin from Agave americana Leaf using HPTLC. Chromatographia 2006, 64, 113–116. [Google Scholar] [CrossRef]
- Peana, A.T.; Moretti, M.D.; Manconi, V.; Desole, G.; Pippia, P. Anti-inflammatory activity of aqueous extracts and steroidal sapogenins of Agave Americana. Planta Med. 1997, 63, 199–202. [Google Scholar] [CrossRef]
- Santos Cerqueira, G.; dos Santos e Silva, G.; Vasconcelos, R.E.; de Freitas, F.A.P.; Moura, A.B.; Macedo, S.D.; Souto, L.A.; Filho, B.J.M.; de Leal, A.L.K.; de Brito, C.G.A.; et al. Effects of hecogenin and its possible mechanism of action on experimental models of gastric ulcer in mice. Eur. J. Pharmacol. 2012, 683, 260–269. [Google Scholar]
- Corbiere, C.; Liagre, B.; Bianchi, A.; Bordji, K.; Dauça, M.; Netter, P.; Beneytout, J.L. Different contribution of apoptosis to the antiproliferative effects of diosgenin and other plant steroids, hecogenin and tigogenin, on human 1547 osteosarcoma cells. Int. J. Oncol. 2003, 4, 899–905. [Google Scholar]
- Villarreal, C.F.; Funez, M.I.; Figueiredo, F.; Cunha, F.Q.; Parada, C.A.; Ferreira, S.H. Acute and persistent nociceptive paw sensitisation in mice: The involvement of distinct signalling pathways. Life Sci. 2009, 85, 822–829. [Google Scholar] [CrossRef]
- Salvemini, D.; Wang, Z.Q.; Wyatt, P.S.; Bourdon, D.M.; Marino, M.H.; Manning, P.T.; Currie, M.G. Nitric oxide: A key mediator in the early and late phase of carrageenan-induced rat paw inflammation. Br. J. Pharmacol. 1996, 118, 829–838. [Google Scholar] [CrossRef]
- Ferreira, S.H.; Lorenzetti, B.B.; Poole, S. Bradykinin initiates cytokine-mediated inflammatory hyperalgesia. Br. J. Pharmacol. 1993, 110, 1227–1231. [Google Scholar] [CrossRef]
- Morris, C.J. Carrageenan-induced paw edema in the rat and mouse. Methods Mol. Biol. 2003, 225, 115–121. [Google Scholar]
- Cunha, T.M.; Verri, W.A., Jr.; Silva, J.S.; Poole, S.; Cunha, F.Q.; Ferreira, S.H. A cascade of cytokines mediates mechanical inflammatory hypernociception in mice. Proc. Natl. Acad. Sci. USA 2005, 102, 1755–1760. [Google Scholar]
- Dray, A. Inflammatory mediators of pain. Br. J. Anaesth. 1995, 75, 125–131. [Google Scholar] [CrossRef]
- Toulmond, S.; Parnet, P.; Linthorst, A.C. When cytokines get on your nerves: Cytokine networks and CNS pathologies. Trends Neurosci. 1996, 19, 409–410. [Google Scholar] [CrossRef]
- Cunha, F.Q.; Ferreira, S.H. Peripheral hyperalgesic cytokines. Adv. Exp. Med. Biol. 2003, 521, 22–39. [Google Scholar]
- Brown, K.A.; Brain, S.D.; Edgeworth, J.D.; Lewis, S.M.; Treacher, D.F. Neutrophils in development of multiple organ failure in sepsis. Lancet 2006, 368, 157–169. [Google Scholar] [CrossRef]
- Verri, W.A., Jr.; Cunha, T.M.; Parada, C.A.; Poole, S.; Cunha, F.Q.; Ferreira, S.H. Hypernociceptive role of cytokines and chemokines: Targets for analgesic drug development? Pharmacol. Ther. 2006, 112, 116–138. [Google Scholar] [CrossRef]
- Oliveira, M.G.B.; Marques, R.B.; Santana, M.F.; Santos, A.B.D.; Brito, F.A.; Barreto, E.O.; de Sousa, D.P.; Almeida, F.R.C.; Badauê Passos, D., Jr.; Antoniolli, A.R.; et al. α-terpineol reduces mechanical hypernociception and inflammatory response. Basic Clin. Pharmacol. Toxicol. 2012, 111, 120–125. [Google Scholar]
- Brito, E.M.; Abreu, R.N.D.C.; Honório Júnior, J.E.R.; Barbosa-Filho, J.M.; Florenço de Sousa, F.C.; Silva, M.C.C.; Lima, M.M.B.C.; Menezes Rabelo, G.; França Fonteles, M.M.; Vasconcelos, S.M.M. Behavioral effects and parameters of oxidative stress with hecogenin in central nervous system of mice. Behav. Brain Res. 2011, 21, S543. [Google Scholar]
- Gama, K.B.; Quintans, J.S.S.; Antoniolli, A.R.; Quintans-Júnior, L.J.; Santana, W.A.; Branco, A.; Soares, M.B.P.; Villarreal, C.F. Evidence for the involvement of descending pain-inhibitory mechanisms in the antinociceptive effect of hecogenin acetate. J. Nat. Prod. 2013, 76, 559–563. [Google Scholar] [CrossRef]
- Sachs, D.; Cunha, F.Q.; Ferreira, S.H. Peripheral analgesic blockade of hypernociception: Activation of arginine/NO/cGMP/protein kinase G/ATP-sensitive K+ channel pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 3680–3685. [Google Scholar] [CrossRef]
- Willis, W.D.; Coggeshall, R.E. Sensory Mechanisms of the Spinal Cord, 3rd ed.; Plenum: New York, NY, USA, 2004. [Google Scholar]
- Baba, H.; Doubell, T.P.; Woolf, C.J. Peripheral inflammation facilitates Aβ fiber-mediated synaptic input to the substantia gelatinosa of the adult rat spinal cord. J. Neurosci. 1999, 19, 859–867. [Google Scholar]
- Morgan, J.I.; Curran, T. Immediate-early genes: Ten years on. Trends Neurosci. 1995, 18, 66–67. [Google Scholar] [CrossRef]
- Coggeshall, R.E. Fos, nociception and the dorsal horn. Prog. Neurobiol. 2005, 77, 299–352. [Google Scholar]
- Morgado, C.; Terra, P.P.; Tavares, I. Neuronal hyperactivity at the spinal cord and periaqueductal grey during painful diabetic neuropathy: Effects of gabapentin. Eur. J. Pain 2010, 14, 693–699. [Google Scholar] [CrossRef]
- Burchiel, K.J.; Russell, L.C.; Lee, R.P.; Sima, A.A. Spontaneous activity of primary afferent neurons in diabetic BB/Wistar rats. A possible mechanism of chronic diabetic neuropathic pain. Diabetes 1985, 34, 1210–1213. [Google Scholar] [CrossRef]
- Bertrand, S.; Ng, G.Y.; Purisai, M.G.; Wolfe, S.E.; Severidt, M.W.; Nouel, D.; Robitaille, R.; Low, M.J.; O’Neill, G.P.; Metters, K.; et al. The anticonvulsant, antihyperalgesic agent gabapentin is an agonist at brain γ-aminobutyric acid type B receptors negatively coupled to voltage-dependent calcium channels. J. Pharmacol. Exp. Ther. 2001, 298, 15–24. [Google Scholar]
- Guimarães, A.G.; Xavier, M.A.; de Santana, M.T.; Camargo, E.A.; Santos, C.A.; Brito, F.A.; Barreto, E.O.; Cavalcanti, S.C.H.; Antoniolli, Â.R.; Oliveira, R.C.M.; et al. Carvacrol attenuates mechanical hypernociception and inflammatory response. Naunyn. Schmiedebergs Arch. Pharmacol. 2012, 385, 253–263. [Google Scholar]
- Institute for Laboratory Animal Research. Guidance for the Description of Animal Research in Scientific Publications; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Cunha, T.M.; Verri, W.A., Jr.; Vivancos, G.G.; Moreira, I.F.; Reis, S.; Parada, C.A.; Cunha, F.Q.; Ferreira, S.H. An electronic pressure-meter nociception paw test for mice. Braz. J. Med. Biol. Res. 2004, 37, 401–407. [Google Scholar] [CrossRef]
- Quintans-Júnior, L.J.; Melo, M.S.; de Sousa, D.P.; Araujo, A.A.S.; Onofre, A.C.; Gelain, D.P.; Gonçalves, J.C.; Araújo, D.A.; Almeida, J.R.; Bonjardim, L.R. Antinociceptive effects of citronellal in formalin-, capsaicin-, and glutamate-induced orofacial nociception in rodents and its action on nerve excitability. J. Orofac. Pain 2010, 24, 305–312. [Google Scholar]
- Barr, G.A. Formalin-induced c-fos expression in the brain of infant rats. J. Pain 2011, 12, 263–271. [Google Scholar] [CrossRef]
- Brito, R.G.; Santos, P.L.; Prado, D.S.; Santana, M.T.; Araújo, A.A.S.; Bonjardim, L.R.; Santos, M.R.V.; de Lucca Júnior, W.; Oliveira, A.P.; Quintans-Júnior, L.J. Citronellol reduces orofacial nociceptive behavior in mice - evidence of involvement of retrosplenial cortex and periaqueductal gray areas. Basic Clin. Pharmacol. Toxicol. 2013, 112, 215–221. [Google Scholar] [CrossRef]
- Kassuya, C.A.; Silvestre, A.A.; Rehder, V.L.; Calixto, J.B. Anti-allodynic and anti-oedematogenic properties of the extract and lignans from Phyllanthus amarus in models of persistent inflammatory and neuropathic pain. Eur. J. Pharmacol. 2003, 478, 145–153. [Google Scholar] [CrossRef]
- Sample Availability: Samples of compounds hecogenin acetate, carrageenan, and PGE2 are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Quintans, J.S.S.; Barreto, R.S.S.; De Lucca, W., Júnior; Villarreal, C.F.; Kaneto, C.M.; Soares, M.B.P.; Branco, A.; Almeida, J.R.G.S.; Taranto, A.G.; Antoniolli, A.R.; et al. Evidence for the Involvement of Spinal Cord-Inhibitory and Cytokines-Modulatory Mechanisms in the Anti-Hyperalgesic Effect of Hecogenin Acetate, a Steroidal Sapogenin-Acetylated, in Mice. Molecules 2014, 19, 8303-8316. https://doi.org/10.3390/molecules19068303
Quintans JSS, Barreto RSS, De Lucca W Júnior, Villarreal CF, Kaneto CM, Soares MBP, Branco A, Almeida JRGS, Taranto AG, Antoniolli AR, et al. Evidence for the Involvement of Spinal Cord-Inhibitory and Cytokines-Modulatory Mechanisms in the Anti-Hyperalgesic Effect of Hecogenin Acetate, a Steroidal Sapogenin-Acetylated, in Mice. Molecules. 2014; 19(6):8303-8316. https://doi.org/10.3390/molecules19068303
Chicago/Turabian StyleQuintans, Jullyana S.S., Rosana S.S. Barreto, Waldecy De Lucca, Júnior, Cristiane F. Villarreal, Carla M. Kaneto, Milena B.P. Soares, Alexsandro Branco, Jackson R.G.S. Almeida, Alex G. Taranto, Angelo R. Antoniolli, and et al. 2014. "Evidence for the Involvement of Spinal Cord-Inhibitory and Cytokines-Modulatory Mechanisms in the Anti-Hyperalgesic Effect of Hecogenin Acetate, a Steroidal Sapogenin-Acetylated, in Mice" Molecules 19, no. 6: 8303-8316. https://doi.org/10.3390/molecules19068303