α-Glucosidase Inhibitory Activity of Polyphenols from the Burs of Castanea mollissima Blume

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
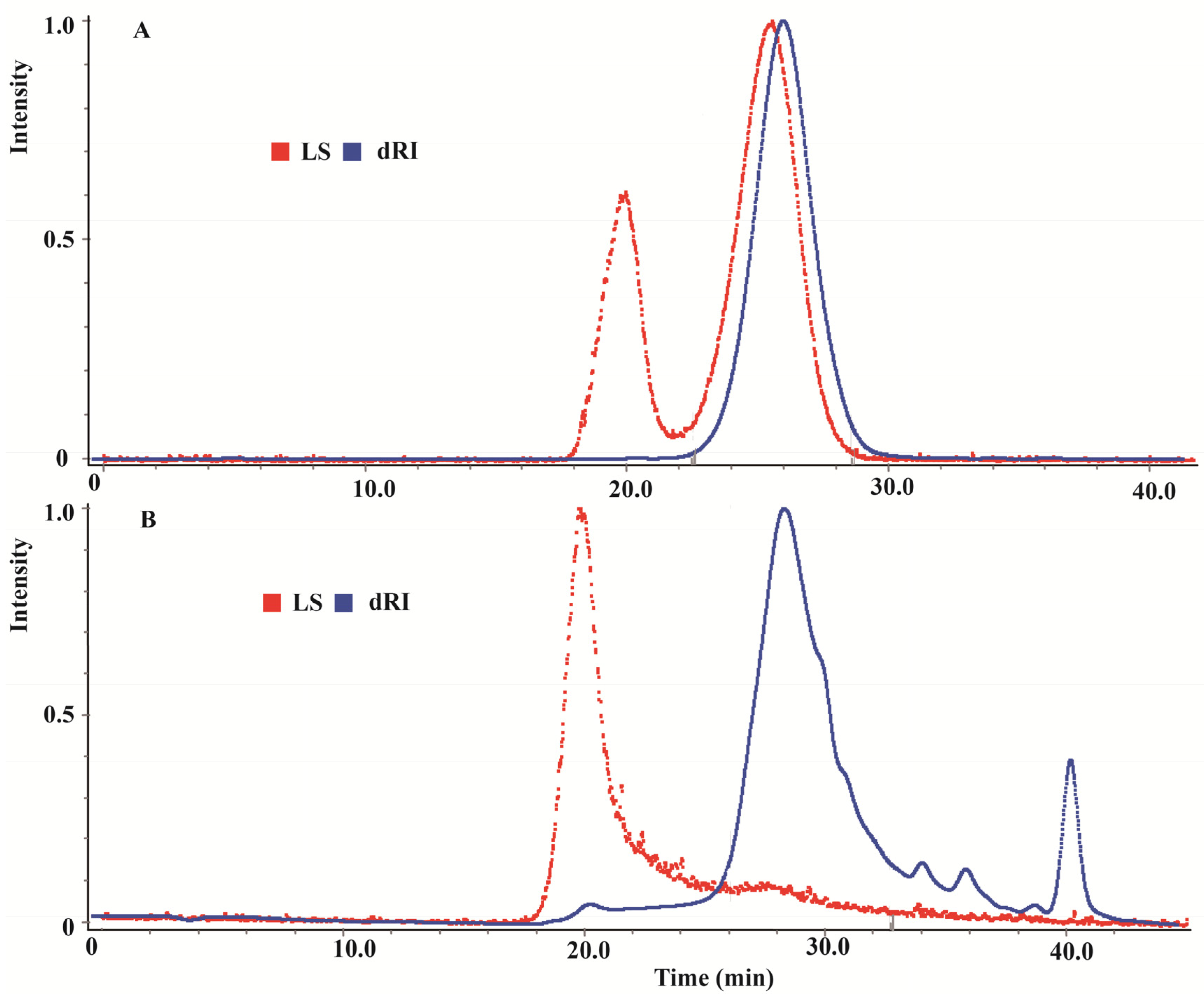
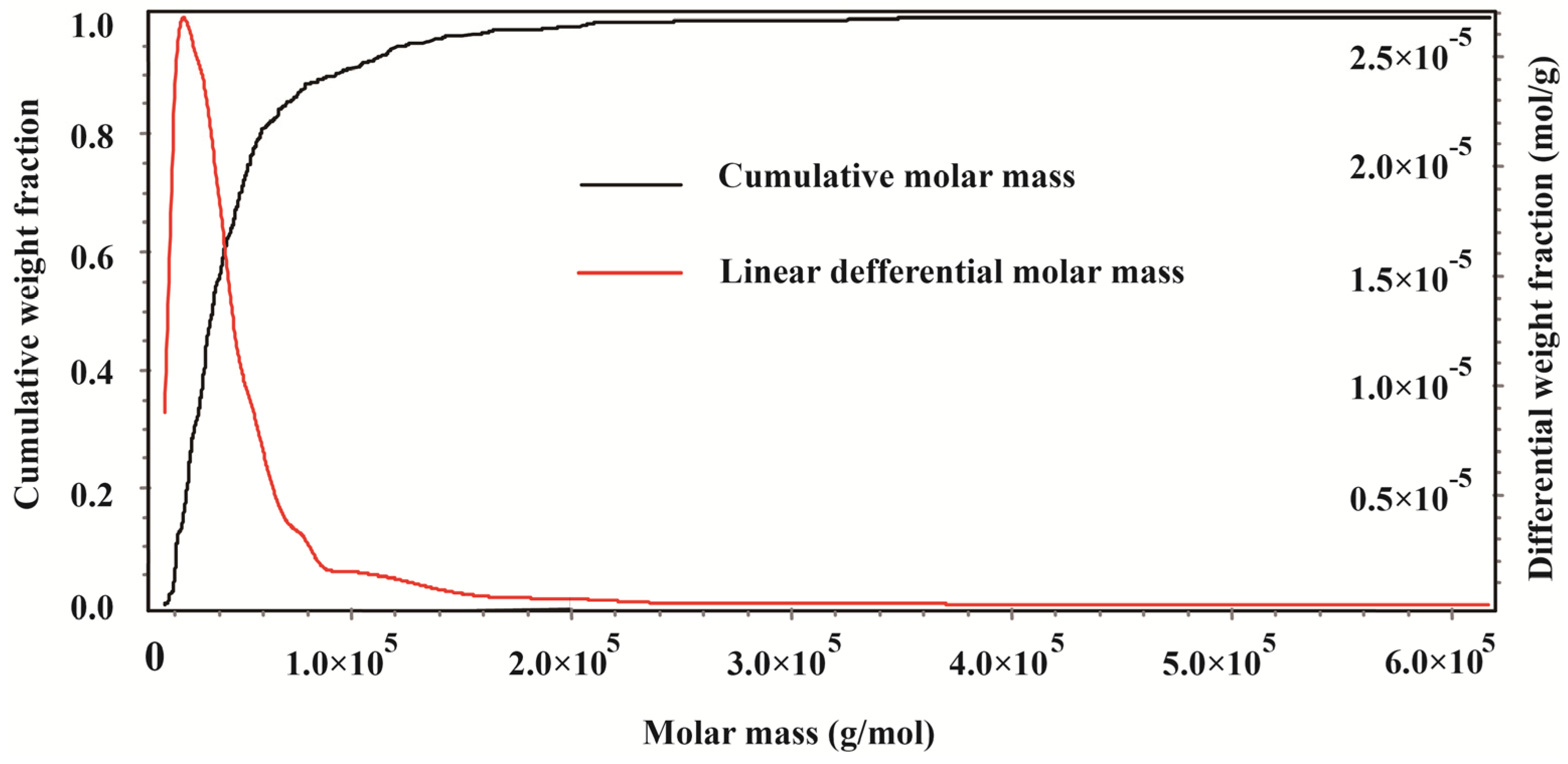
2.1. Gel Permeation Chromatography of CMPE


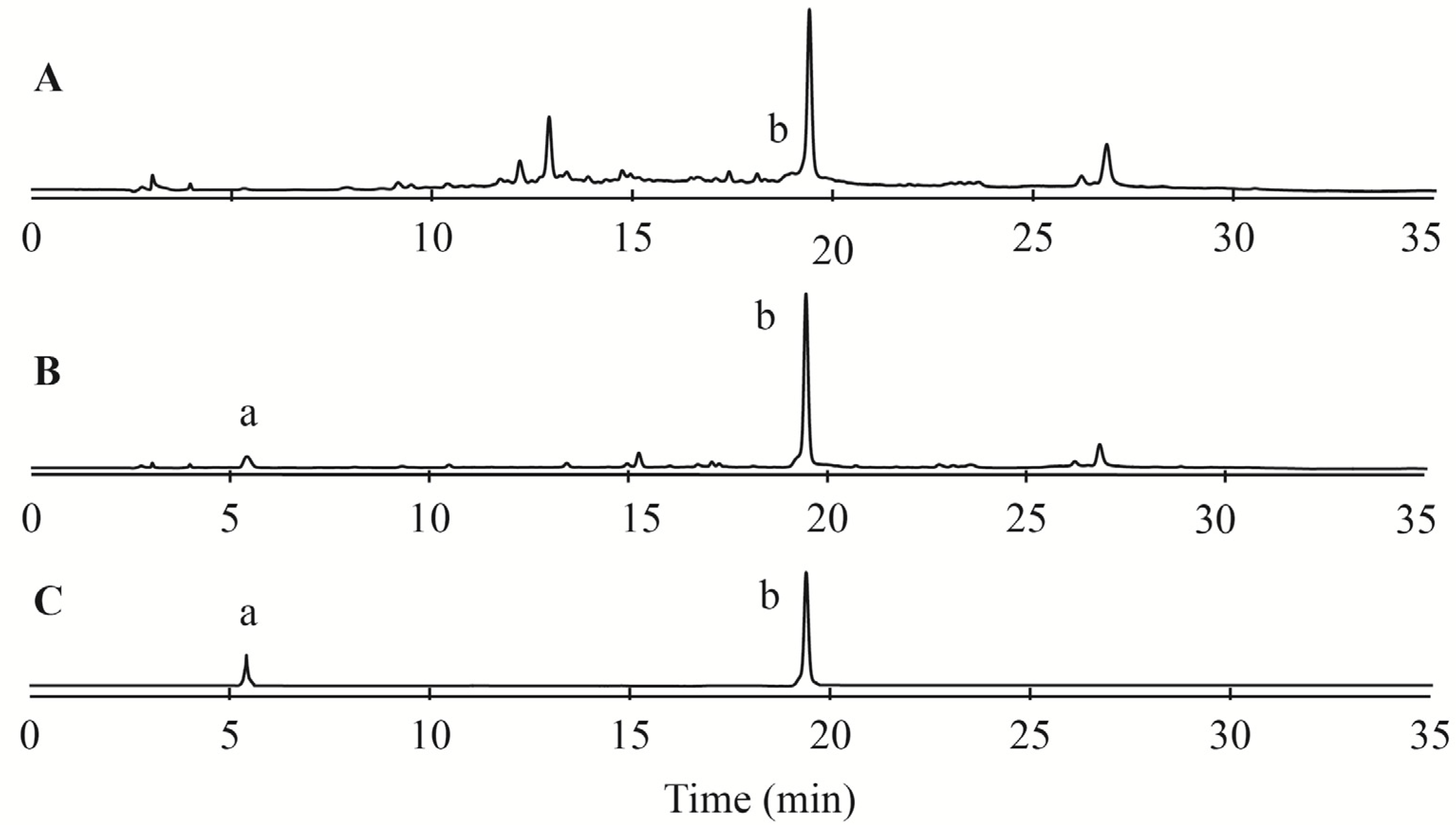
2.2. Acid Hydrolysis Analysis of CMPE

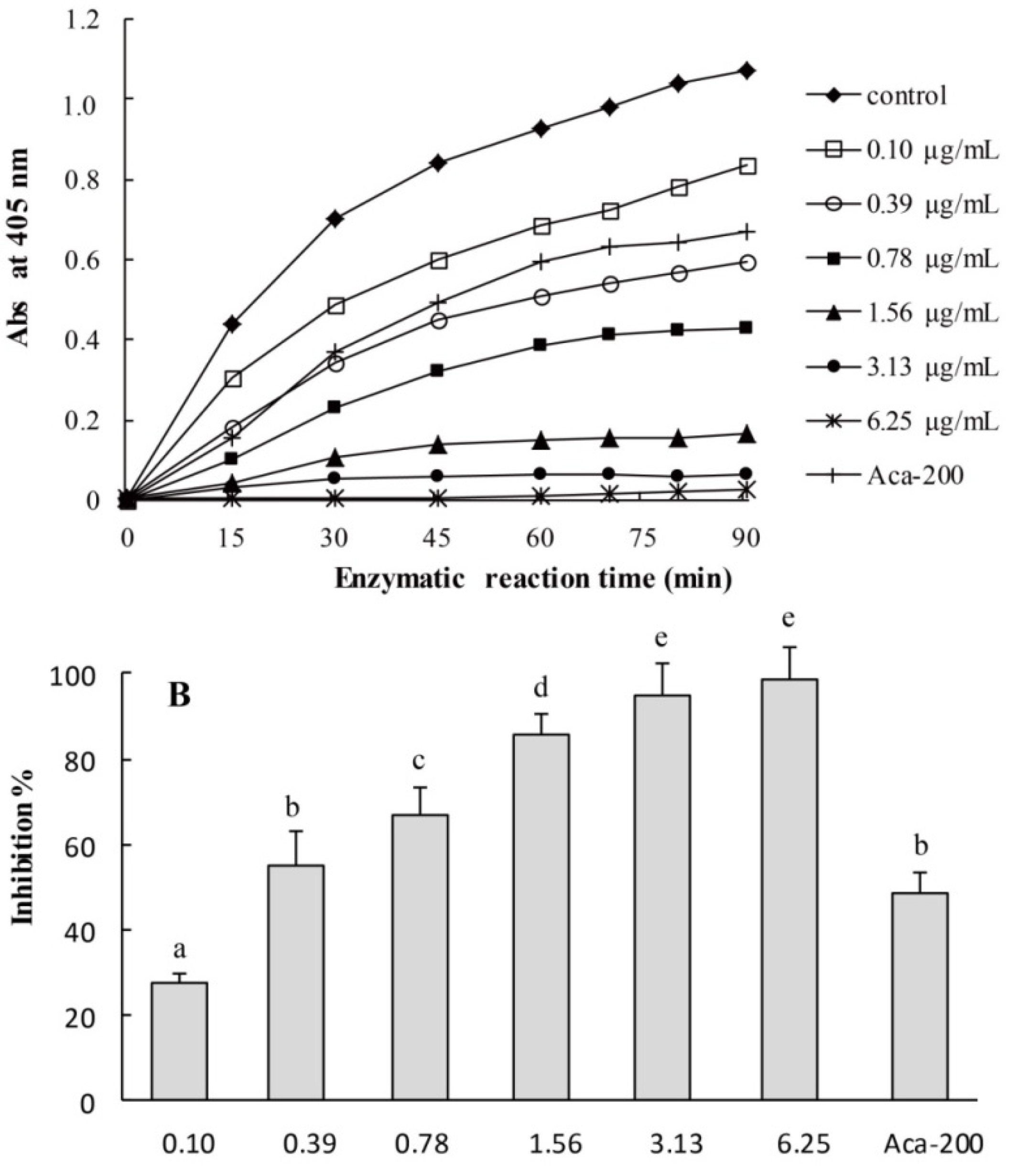
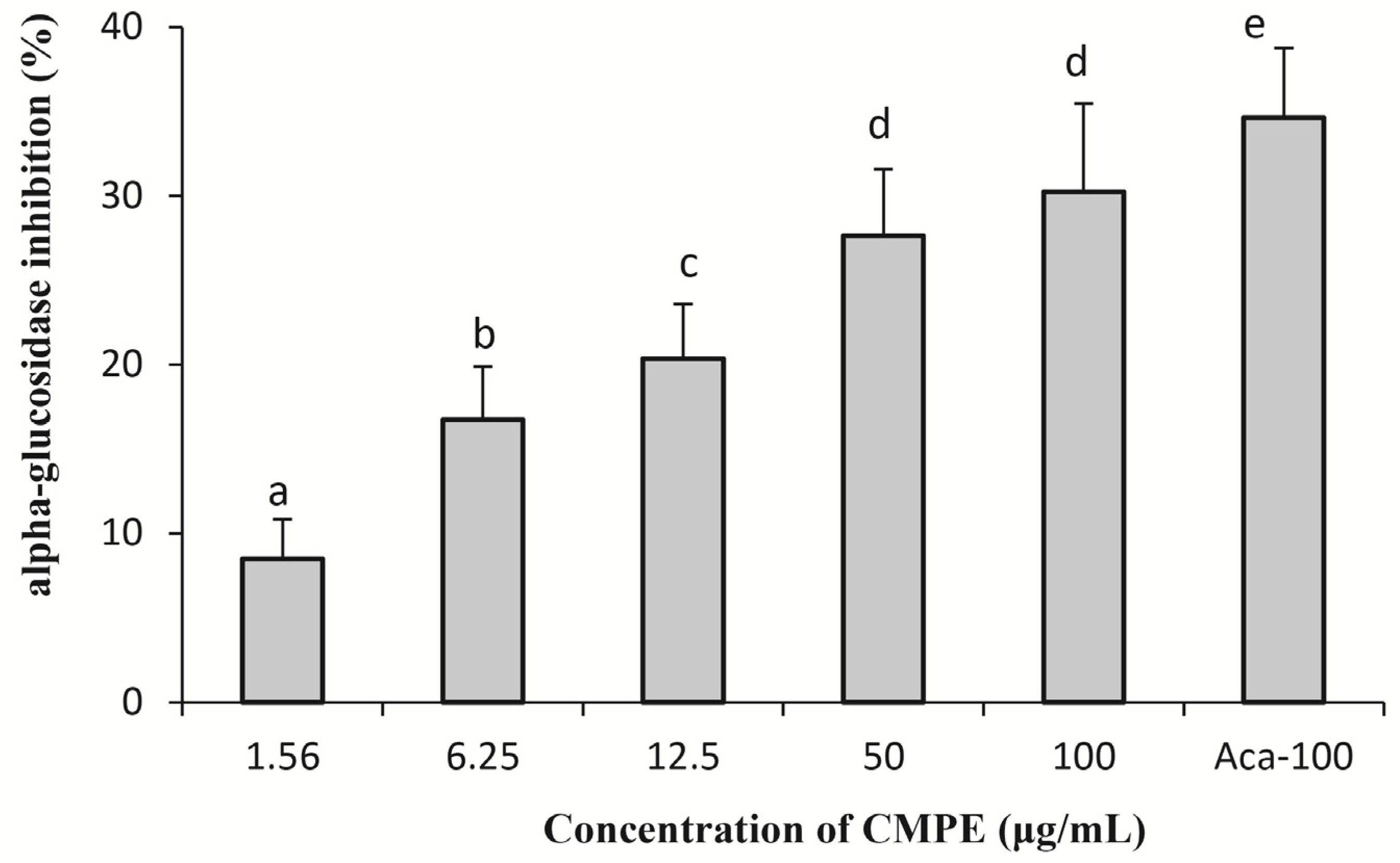
2.3. Yeast Alpha-Glucosidase Inhibition Assays of CMPE


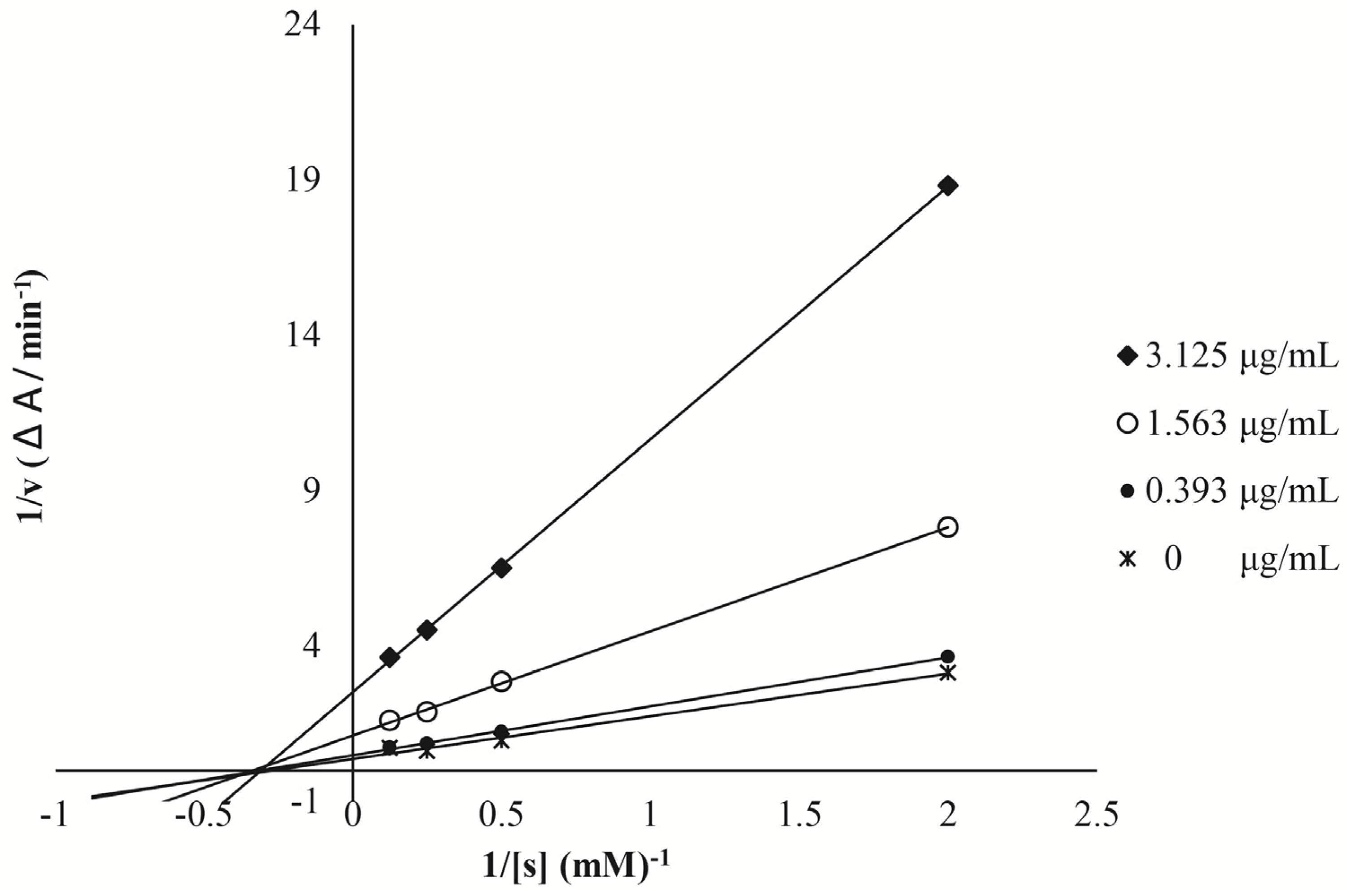
2.4. In Vitro Mammalian α-Glucosidase Inhibition Assays of CMPE
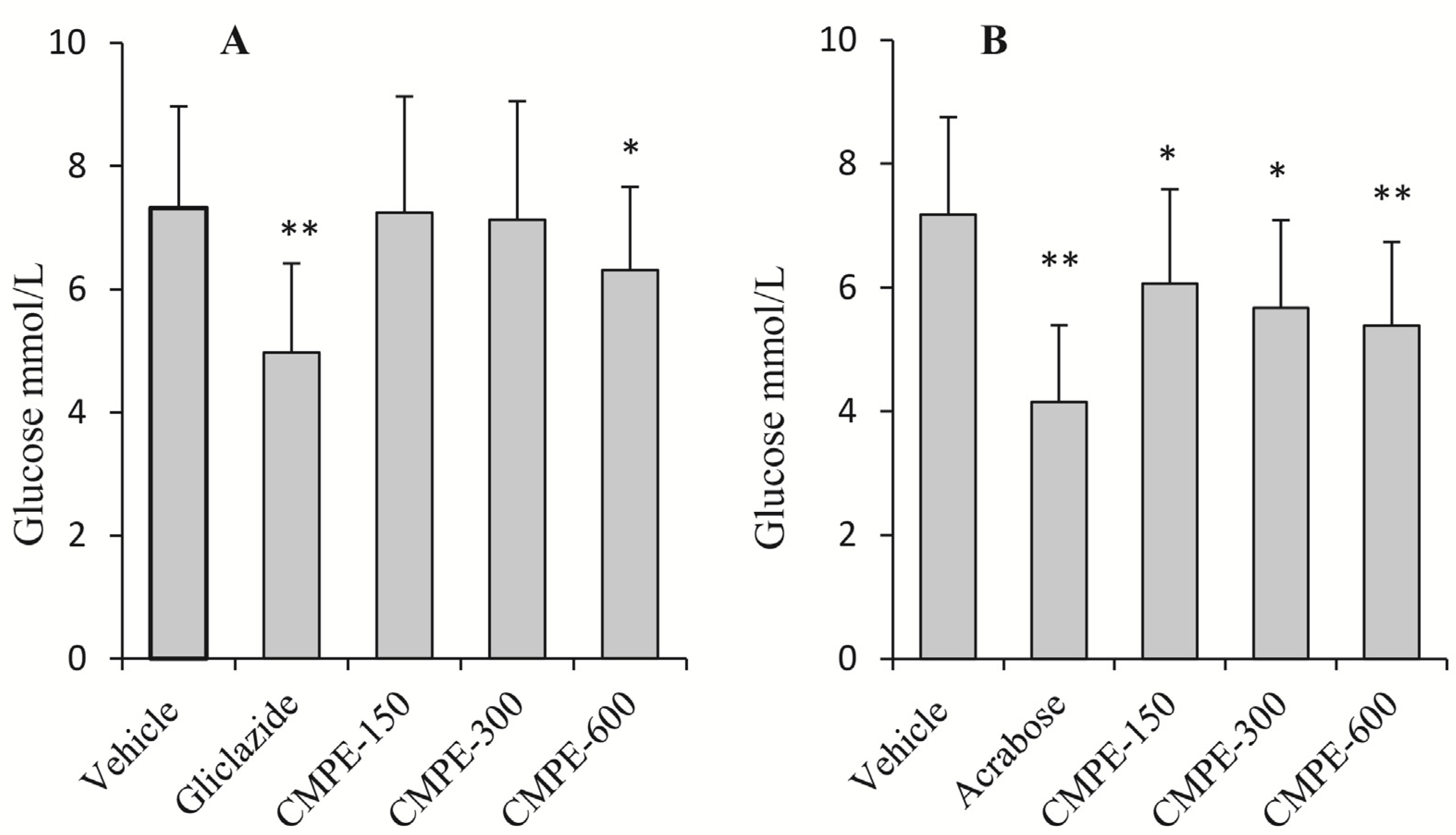
2.5. In Vivo Mammalian α-Glucosidase CMPE Inhibition Assays


2.6. Discussion
3. Experimental
3.1. Plant Materials and Chemicals
3.2. Animals
3.3. Preparation of Polyphenol Extracts of C. mollissima
3.4. Gel Permeation Chromatography of CMPE
3.5. Acid Hydrolysis of CMPE
3.6. In Vitro Yeast α-Glucosidase Inhibition Assays
3.7. In Vitro Mammalian α-Glucosidase Inhibition Assays
3.8. Influence of CMPE on in Vivo Serum Glucose Levels
3.9. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, J.S.; Hyun, T.K.; Kim, M.J. The inhibitory effects of ethanol extracts from sorghum, foxtail millet and proso millet on α-glucosidase and α-amylase activities. Food Chem. 2011, 124, 1647–1651. [Google Scholar] [CrossRef]
- Yin, P.P.; Zhao, S.; Chen, S.Y.; Liu, J.Y.; Shi, L.L.; Wang, X.J.; Liu, Y.J.; Ma, C. Hypoglycemic and hypolipidemic effects of polyphenols from burs of Castanea mollissima Blume. Molecules 2011, 16, 9764–9774. [Google Scholar] [CrossRef]
- Ye, X.P.; Song, C.Q.; Yuan, P.; Mao, R.G. α-Glucosidase and α-amylase inhibitory activity of common constituents from traditional Chinese medicine used for diabetes mellitus. Chin. J. Nat. Med. 2010, 8, 349–352. [Google Scholar]
- Casirola, D.M.; Ferraris, R.P. Alpha-Glucosidase inhibitors prevent diet-induced increases in intestinal sugar transport in diabetic mice. Metabolism 2006, 55, 832–841. [Google Scholar] [CrossRef]
- Zhang, L.; Hogan, S.; Li, J.R.; Sun, S.; Canning, C.; Zheng, S.J. Grape skin extract inhibits mammalian intestinal a-glucosidase activitand suppresses postprandial glycemic response in streptozocin-treated mice. Food Chem. 2011, 126, 466–471. [Google Scholar] [CrossRef]
- Madar, Z. The effect of acarbose and miglitol (BAY-M-1099) on postprandial glucose levels following ingestion of various sources of starch by non-diabetic and streptozotocin-induced diabetic rats. J. Nutr. 1989, 119, 2023–2029. [Google Scholar]
- Hiroyuki, F.; Tomohide, Y.; Kazunori, O. Efficacy and safety of Touchi extract, an alpha-glucosidase inhibitor derived from fermented soybeans, in non-insulin-dependent diabetic mellitus. J. Nutr. Biochem. 2001, 12, 351–356. [Google Scholar] [CrossRef]
- Murai, A.; Iwamura, K.; Takada, M.; Ogawa, K.; Usui, T.; Okumura, J. Control of postprandial hyperglycaemia by galactosyl maltobionolactone and its novel anti-amylase effect in mice. Life Sci. 2002, 71, 1405–1415. [Google Scholar] [CrossRef]
- Amarowicz, R.; Troszyńska, A.; Shahidi, F. Antioxidant activity of almond seed extract and its fractions. J. Food Lipids 2005, 12, 344–358. [Google Scholar] [CrossRef]
- Fujisawa, T.; Ikegami, H.; Inoue, K.; Kawabata, Y.; Ogihara, T. Effect of two alpha-glucosidase inhibitors, voglibose and acarbose, on postprandial hyperglycemia correlates with subjective abdominal symptoms. Metabolism 2005, 54, 387–390. [Google Scholar] [CrossRef]
- Shobana, S.; Sreerama, Y.N.; Malleshi, N.G. Composition and enzyme inhibitory properties of finger millet (Eleusine coracana L.) seed coat phenolics: Mode of inhibition of α-glucosidase and pancreatic amylase. Food Chem. 2009, 115, 1268–1273. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.J.; Song, H.C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar]
- Liu, L.; Deseo, M.A.; Morris, C.; Winter, K.M.; Leach, D.N. Investigation of α-glucosidase inhibitory activity of wheat bran and germ. Food Chem. 2011, 126, 553–561. [Google Scholar] [CrossRef]
- Bakkalbaşi, E.; Menteş, O.; Artik, N. Food ellagitannins-occurrence, effects of processing and storage. Crit. Rev. Food. Sci. Nutr. 2009, 49, 283–298. [Google Scholar]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Törrönen, A.R. Identification and cuantification of phenolics compounds of Fragaria and Rubus species (Family Rosaseae). J. Agric. Food Chem. 2004, 52, 6178–6187. [Google Scholar]
- Kaponen, J.M.; Happonen, A.M.; Mattila, P.H.; Törrönen, A. Contents of anthocyanins and ellagitannins in selected foods consumed in Finland. J. Agric. Food. Chem. 2007, 55, 1612–1619. [Google Scholar] [CrossRef]
- Augilera-Carbo, A.; Augur, C.; Prado-Barragan, L.A.; Favela-Torres, E.; Aguilar, C.N. Microbial production of ellagic acid and biodegradation of ellagitannins. Appl. Microbiol. Biotechnol. 2008, 78, 189–199. [Google Scholar] [CrossRef]
- Spiller, H.A.; Sawyer, T.S. Toxicology of oral antidiabetic medications. Am. J. Health-Sys. Ph. 2006, 63, 929–938. [Google Scholar] [CrossRef]
- Gao, H.Y.; Wu, D.; Lu, C.; Xu, X.M.; Huang, J.; Sun, B.H.; Wu, L.J. Constituents from the testas of Castanea mollissima Blume with alpha-glucosidase inhibitory activity. J. Asian Nat. Prod. Res. 2010, 12, 144–149. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, J.Y.; Chen, S.Y.; Shi, L.L.; Liu, Y.J.; Ma, C. Antioxidant potential of polyphenols and tannins from burs of Castanea mollissima Blume. Molecules 2011, 16, 8590–8600. [Google Scholar] [CrossRef]
- Karonen, M.; Parker, J.; Agrawal, A.; Salminen, J.P. First evidence of hexameric and heptameric ellagitannins in plants detected by liquid chromatography/electrospray ionisation mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 3151–3156. [Google Scholar]
- Adachi, T.M.C.; Sakurai, K.; Shihara, N.; Tsuda, K.; Yasuda, K. Morphological changes and increased sucrase and isomaltase acitivity in small intestines of insulin-deficient and type-2 diabetic rats. J. Endocrinol. 2003, 50, 271–279. [Google Scholar]
- Yao, Z.; Qi, J.; Wang, L. Isolation, fractionation and characterization of melanin-like pigments from chestnut (Castanea mollissima) shells. J. Food Sci. 2012, 77, C671–C676. [Google Scholar] [CrossRef]
- Wang, S.; Ding, X.B.; Chen, Y.F.; Yu, D.Q. Chestnutamide: A novel alkaloid from flowers of Castanea mollissima Blume. J. Asian Nat. Prod. Res. 2001, 3, 89–93. [Google Scholar] [CrossRef]
- Vázquez, G.; Fernández-Agulló, A.; Gómez-Castro, C.; Freire, M.S.; Antorrena, G.; González-Álvarez, J. Response surface optimization of antioxidants extraction from chestnut (Castanea sativa) bur. Ind. Crops Prod. 2012, 35, 126–134. [Google Scholar] [CrossRef]
- Pinto, M.S.; Lajolo, F.M.; Genovese, M.I. Bioactive compounds and quantification of total ellagic acid in strawberries (Fragaria x ananassa Duch.). Food Chem. 2008, 107, 1629–1635. [Google Scholar] [CrossRef]
- Si, M.M.; Lou, J.S.; Zhou, C.X.; Shen, J.N.; Wu, H.H.; Yang, B.; He, Q.J.; Wu, H.S. Insulin releasing and alpha-glucosidase inhibitory activity of ethyl acetate fraction of Acorus calamus in vitro and in vivo. J. Ethnopharmacol. 2010, 128, 154–159. [Google Scholar] [CrossRef]
- Oki, T.; Matsui, T.; Osajima, Y. Inhibitory effect of α-glucosidase inhibitors varies according to its origin. J. Agric. Food Chem. 1999, 47, 550–553. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the polyphenol extracts from the burs of Castanea mollissima Blume are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, J.; Zhao, S.; Yin, P.; Yan, L.; Han, J.; Shi, L.; Zhou, X.; Liu, Y.; Ma, C. α-Glucosidase Inhibitory Activity of Polyphenols from the Burs of Castanea mollissima Blume. Molecules 2014, 19, 8373-8386. https://doi.org/10.3390/molecules19068373
Zhang J, Zhao S, Yin P, Yan L, Han J, Shi L, Zhou X, Liu Y, Ma C. α-Glucosidase Inhibitory Activity of Polyphenols from the Burs of Castanea mollissima Blume. Molecules. 2014; 19(6):8373-8386. https://doi.org/10.3390/molecules19068373
Chicago/Turabian StyleZhang, Jianwei, Shan Zhao, Peipei Yin, Linlin Yan, Jin Han, Lingling Shi, Xiaojing Zhou, Yujun Liu, and Chao Ma. 2014. "α-Glucosidase Inhibitory Activity of Polyphenols from the Burs of Castanea mollissima Blume" Molecules 19, no. 6: 8373-8386. https://doi.org/10.3390/molecules19068373