Reduction of Adhesion Molecule Production and Alteration of eNOS and Endothelin-1 mRNA Expression in Endothelium by Euphorbia hirta L. through Its Beneficial β-Amyrin Molecule

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
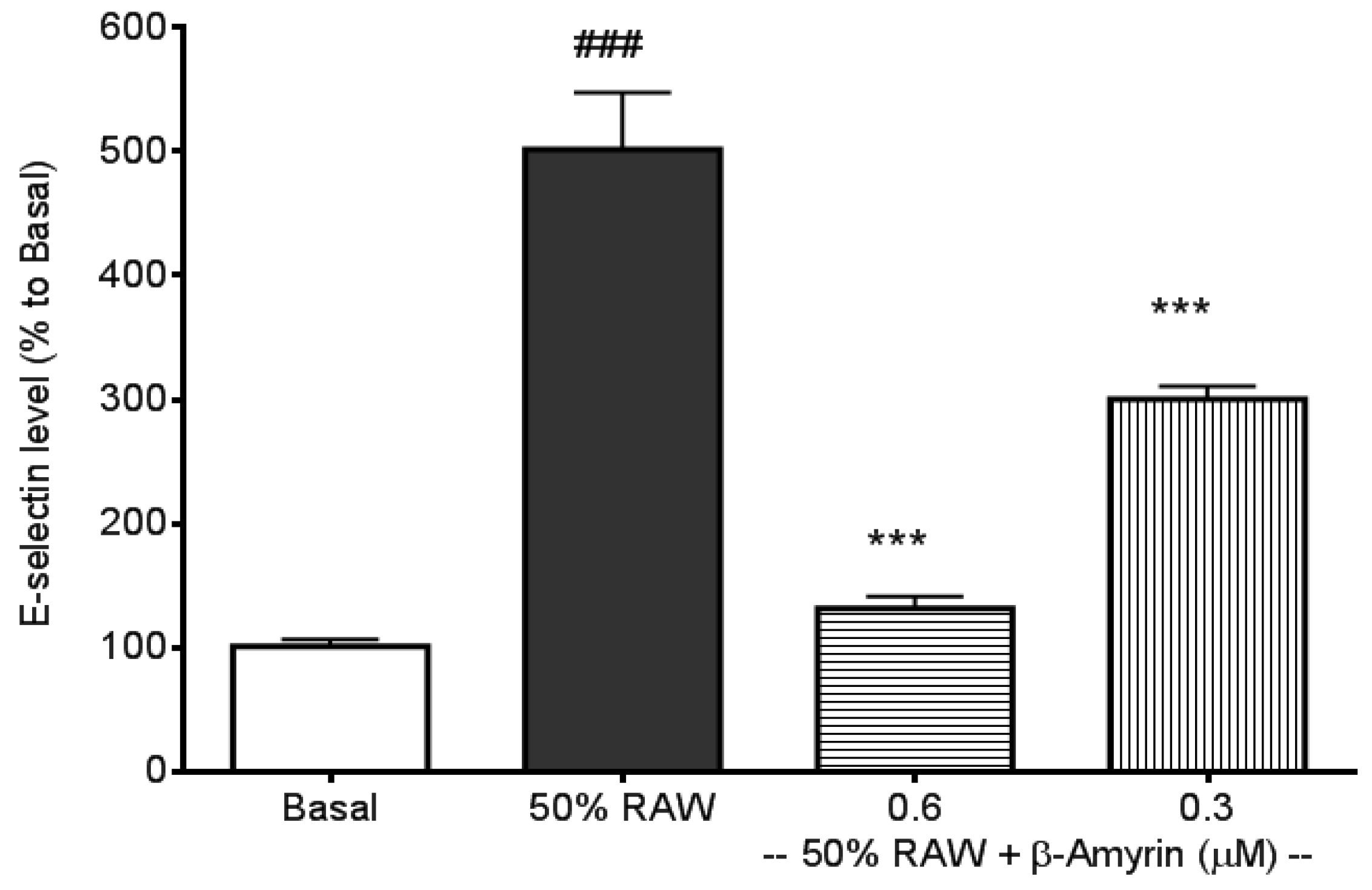
2.1. Inhibitory Effects of β-amyrin on Proinflammatory Cytokine-Induced E-selectin, sICAM-1 and sVCAM-1 Production



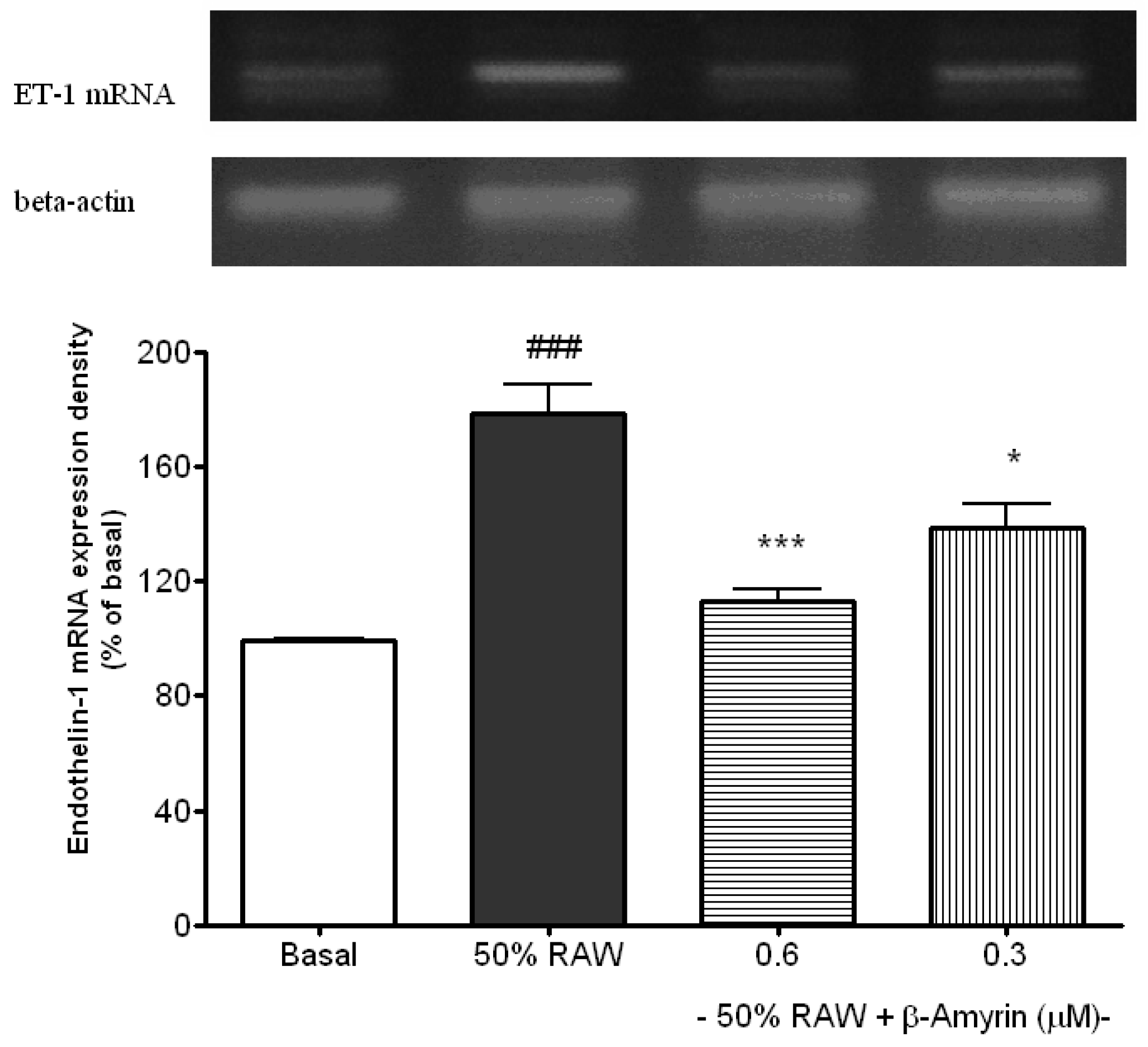
2.2. Inhibitory Effects of β-amyrin on Proinflammatory Cytokine-Induced ET-1 Gene Expression
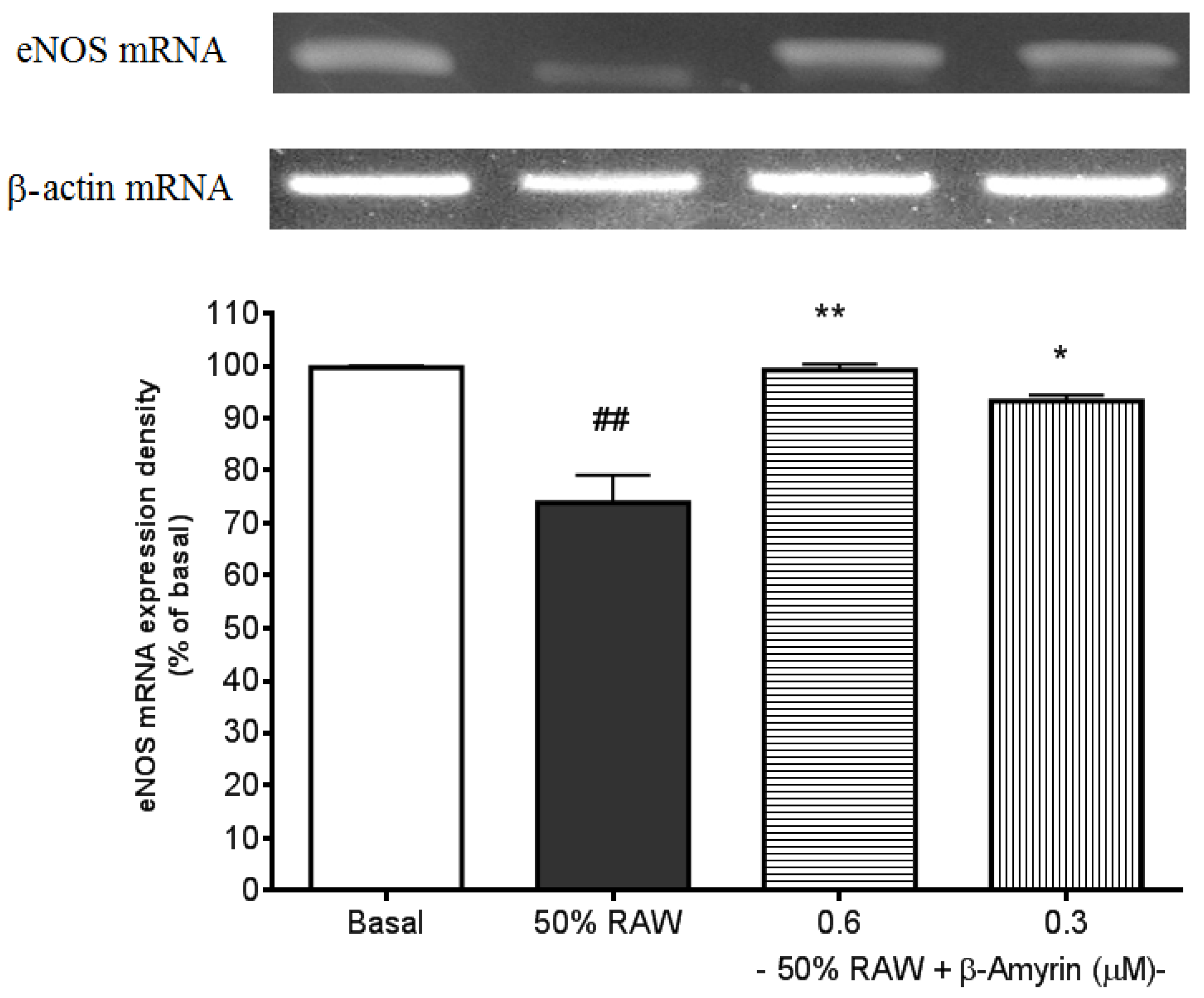
2.3. Ameliorative Effects of β-amyrin on Proinflammatory Cytokine-Suppressed eNOS mRNA Expression


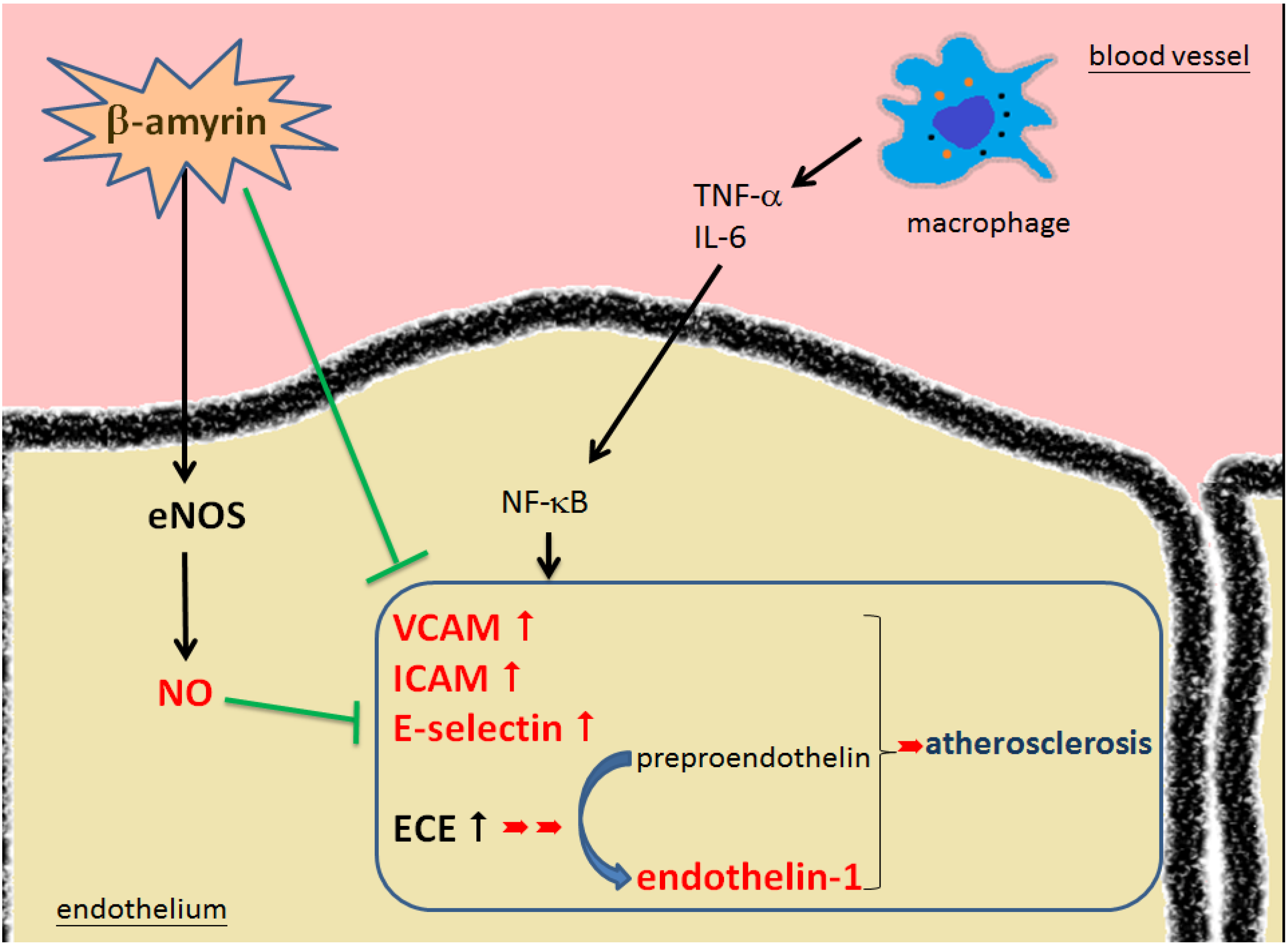
2.4. Pathways of β-amyrin’s Effectiveness Leading to Prevent the Development of Atherosclerosis

3. Experimental Section
3.1. Materials
3.2. Adhesion Molecules — E-selectin, sICAM-1 and sVCAM-1 Level Measurement
3.3. RNA Preparation and ET-1 and eNOS RNA Analysis by Reverse Transcription-Polymerase Chain Reaction
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ross, R. Atherosclerosis—An inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Jhaveri, K.A.; Debnath, P.; Chernoff, J.; Sanders, J.; Schwartz, M.A. The role of p21-activated kinase in the initiation of atherosclerosis. BMC Cardiovasc. Disord. 2012, 12, 55. [Google Scholar] [CrossRef]
- Kacimi, R.; Karliner, J.S.; Koudssi, F.; Long, C.S. Expression and Regulation of Adhesion Molecules in Cardiac Cells by Cytokines: Response to Acute Hypoxia. Circ. Res. 1998, 82, 576–586. [Google Scholar] [CrossRef]
- Ridker, P.M.; Buring, J.E.; Rifai, N. Soluble P-selectin and the risk of future cardiovascular events. Circulation 2001, 103, 491–495. [Google Scholar] [CrossRef]
- Nasuno, A.; Matsubara, T.; Hori, T.; Higuchi, K.; Imai, S.; Nakagawa, I.; Tsuchida, K.; Ozaki, K.; Mezaki, T.; Tanaka, T.; et al. Levels of soluble E-selectin and ICAM-1 in the coronary circulation of patients with stable coronary artery disease: Association with the severity of coronary atherosclerosis. Jap. Heart J. 2002, 43, 93–101. [Google Scholar] [CrossRef]
- Ridker, P.M.; Hennekens, C.H.; Buring, J.E.; Rifai, N. C-reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women. N. Engl. J. Med. 2000, 342, 836–843. [Google Scholar] [CrossRef]
- Blankenberg, S.; Barbaux, S.; Tiret, L. Adhesion molecules and atherosclerosis. Atherosclerosis 2003, 170, 191–203. [Google Scholar] [CrossRef]
- Nahrendorf, M.; Jaffer, F.A.; Kelly, K.A.; Sosnovik, D.E.; Aikawa, E.; Libby, P.; Weissleder, R. Noninvasive vascular cell adhesion molecule-1 imaging identifies inflammatory activation of cells in atherosclerosis. Circulation 2006, 114, 1504–1511. [Google Scholar] [CrossRef]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep. 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Blake, G.J.; Ridker, P.M. Inflammatory bio-markers and cardiovascular risk prediction. J. Int. Med. 2002, 252, 283–294. [Google Scholar] [CrossRef]
- Andreotti, F.; Porto, I.; Crea, F.; Maseri, A. Inflammatory gene polymorphisms and ischemic heart disease: Review of population association studies. Heart 2002, 87, 107–112. [Google Scholar] [CrossRef]
- Lu, H.-H.; Sheng, Z.-Q.; Wang, Y.; Zhang, L. Levels of soluble adhesion molecules in patients with various clinical presentations of coronary atherosclerosis. Chin. Med. J. (Engl.) 2010, 123, 3123. [Google Scholar]
- Tardif, J.C.; Gregoire, J.; Lavoie, M.A.; L’Allier, P.L. Vascular protectants for the treatment of atherosclerosis. Expert. Rev. Cardiovasc. 2003, 1, 385–392. [Google Scholar] [CrossRef]
- Merchant, S.H.; Gurule, D.M.; Larson, R.S. Amelioration of ischemia reperfusion injury with cyclic peptide blockade of ICAM-1. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1260–H1268. [Google Scholar]
- Achmad, T.H.; Rao, G.S. Chemotaxis of human blood monocytes toward endothelin-1 and the influence of calcium channel blockers. Biochem. Biophy. Res. Commun. 1992, 189, 994–1000. [Google Scholar] [CrossRef]
- Ivey, M.E.; Osman, N.; Little, P.J. Endothelin-1 signalling in vascular smooth muscle: Pathways controlling cellular functions associated with atherosclerosis. Atherosclerosis 2008, 199, 237–247. [Google Scholar] [CrossRef]
- Little, P.J.; Ivey, M.E.; Osman, N. Endothelin-1 actions on vascular smooth muscle cell functions as a target for the prevention of atherosclerosis. Curr. Vasc. Pharmacol. 2008, 6, 195–203. [Google Scholar] [CrossRef]
- Davignon, J.; Ganz, P. Role of endothelial dysfunction in atherosclerosis. Circulation 2004, 109, III-27–III-32. [Google Scholar]
- De Caterina, R.; Libby, P.; Peng, H-B.; Thannickal, V.J.; Rajavashisth, T.; Gimbrone, M., Jr.; Shin, W.S.; Liao, J.K. Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J. Clin. Invest. 1995, 96, 60. [Google Scholar] [CrossRef]
- Huang, P.L.; Huang, Z.; Mashimo, H.; Bloch, K.D.; Moskowitz, M.A.; Bevan, J.A.; Fishman, M.C. Hypertension in mice lacking the gene for endothelial nitric oxide synthase. Nature 1995, 377, 239–242. [Google Scholar] [CrossRef]
- Lanhers, M.C.; Fleurentin, J.; Dorfman, P.; Mortier, F.; Pelt, J.M. Analgesic, antipyretic and anti inflammatory properties of Euphorbia hirta. Planta Med. 1991, 57, 225–231. [Google Scholar] [CrossRef]
- Youssouf, M.S.; Kaiser, P.; Tahir, M.; Singh, G.D.; Singh, S.; Sharma, V.K.; Satti, N.K.; Haque, S.E.; Johri, R.K. Anti-anaphylactic effect of Euphorbia hirta. Fitoterapia 2007, 78, 535–539. [Google Scholar] [CrossRef]
- Sudhakar, M.; Rao, C.V.; Rao, P.M.; Raju, D.B.; Venkateswarlu, Y. Antimicrobial activity of Caesalpinia pulcherrima, Euphorbia hirta and Asystasia gangeticum. Fitoterapia 2006, 77, 378–380. [Google Scholar] [CrossRef]
- Elumalai, E.; Prasad, T.; Hemachandran, J.; Therasa, S.V.; Thirumalai, T.; David, E. Extracellular synthesis of silver nanoparticles using leaves of Euphorbia hirta and their antibacterial activities. J. Pharm. Sci. Res. 2010, 2, 549–554. [Google Scholar]
- Rajeh, M.A.B.; Zuraini, Z.; Sasidharan, S.; Latha, L.Y.; Amutha, S. Assessment of Euphorbia hirta L. leaf, flower, stem and root extracts for their antibacterial and antifungal activity and brine shrimp lethality. Molecules 2010, 15, 6008–6018. [Google Scholar] [CrossRef]
- Bakr, R.O.; El-Raey, M.A.E.-A.; Farouk, C.N.; Ahmed, H.E.; El-Said, O.M. Radical Scavenging Activity and Cytotoxicity of Euphorbia hirta L. Int. J. Phytomed. 2013, 4, 525–530. [Google Scholar]
- Johnson, P.B.; Abdurahman, E.M.; Tiam, E.A.; Abdu-Aguye, I.; Hussaini, I.M. Euphorbia hirta leaf extracts increase urine output and electrolytes in rats. J. Ethnopharmacol. 1999, 65, 63–69. [Google Scholar] [CrossRef]
- Singh, G.; Kaiser, P.; Youssouf, M.; Singh, S.; Khajuria, A.; Koul, A.; Bani, S.; Kapahi, B.K.; Satti, N.K.; Suri, K.A.; et al. Inhibition of early and late phase allergic reactions by Euphorbia hirta L. Phytother. Res. 2006, 20, 316–321. [Google Scholar] [CrossRef]
- Shih, M.F.; Cheng, Y.D.; Shen, C.R.; Cherng, J.Y. A molecular pharmacology study into the anti-inflammatory actions of Euphorbia hirta L. on the LPS-induced RAW 264.7 cells through selective iNOS protein inhibition. J. Nat. Med. 2010, 64, 330–335. [Google Scholar] [CrossRef]
- Dinarello, C.A. Biologic basis for interleukin-1 in disease. Blood 1996, 87, 2095–2147. [Google Scholar]
- Cherng, J.Y.; Shen, C.R.; Lin, H.H.; Shih, M.-F. Beneficial effects of Chlorella-11 peptide on blocking the LPS-induced macrophage activation and alleviating the thermal injury induced inflammation in rats. Int. J. Immunopathol. Pharmacol. 2010, 24, 817–826. [Google Scholar]
- Shih, M.F.; Chen, L.C.; Cherng, J.Y. Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability. Mar. Drugs 2013, 11, 3861–3874. [Google Scholar] [CrossRef]
- Granger, D.N.; Vowinkel, T.; Petnehazy, T. Modulation of the inflammatory response in cardiovascular disease. Hypertension 2004, 43, 924–931. [Google Scholar] [CrossRef]
- Krieglstein, C.F.; Granger, D.N. Adhesion molecules and their role in vascular disease. Am. J. Hypertens 2001, 14, 44S–54S. [Google Scholar] [CrossRef]
- Amberger, A.; Hala, M.; Saurwein-Teissl, M.; Metzler, B.; Grubeck-Loebenstein, B.; Xu, Q.; Wick, G. Suppressive effects of anti-inflammatory agents on human endothelial cell activation and induction of heat shock proteins. Mol. Med. 1999, 5, 117–128. [Google Scholar]
- Winkles, J.A.; Alberts, G.F.; Brogi, E.; Libby, P. Endothelin-1 and endothelin receptor mRNA expression in normal and atherosclerotic human arteries. Biochem. Biophys. Res. Commun 1993, 191, 1081–1088. [Google Scholar] [CrossRef]
- Babaei, S.; Picard, P.; Ravandi, A.; Monge, J.C.; Lee, T.C.; Cernacek, P.; Stewart, D.J. Blockade of endothelin receptors markedly reduces atherosclerosis in LDL receptor deficient mice: Role of endothelin in macrophage foam cell formation. Cardiovasc. Res. 2000, 48, 158–167. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.; Higgs, E. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Hwang, H.J.; Jung, T.W.; Hong, H.C.; Choi, H.Y.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Choi, K.M.; Choi, D.S.; Baik, S.H. Progranulin Protects Vascular Endothelium against Atherosclerotic Inflammatory Reaction via Akt/eNOS and Nuclear Factor-kappaB Pathways. PLoS One 2013, 8, e76679. [Google Scholar]
- Zhang, H.; Park, Y.; Wu, J.; Chen, X.; Lee, S.; Yang, J.; Dellsperger, K.C.; Zhang, C. Role of TNF-alpha in vascular dysfunction. Clin. Sci. (Lond.) 2009, 116, 219–230. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef]
- Gemmell, E.; Walsh, L.J.; Savage, N.W.; Seymour, G.J. Adhesion molecule expression in chronic inflammatory periodontal disease tissue. J. Periodontal Res. 2013, 48, 1–21. [Google Scholar]
- Hou, Y.C.; Wu, J.M.; Wang, M.Y.; Wu, M.H.; Chen, K.Y.; Yeh, S.L.; Lin, M.T. Glutamine supplementation attenuates expressions of adhesion molecules and chemokine receptors on T cells in a murine model of acute colitis. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Liu, K.Y.; Hu, S.; Chan, B.C.; Wat, E.C.; Lau, C.B.; Hon, K.L.; Fung, K.P.; Leung, P.C.; Hui, P.C.; Lam, C.W.; et al. Anti-inflammatory and anti-allergic activities of Pentaherb formula, Moutan Cortex (Danpi) and gallic acid. Molecules 2013, 18, 2483–2500. [Google Scholar] [CrossRef]
- O’Connell, K.A.; Edidin, M. A mouse lymphoid endothelial cell line immortalized by simian virus 40 binds lymphocytes and retains functional characteristics of normal endothelial cells. J. Immun. 1990, 144, 521–525. [Google Scholar]
- Prism software, 6th version; GraphPAD Inc.: La Jolla, CA, USA, 2012.
- Sample Availability: Samples of the compounds are commercially available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shih, M.F.; Cherng, J.Y. Reduction of Adhesion Molecule Production and Alteration of eNOS and Endothelin-1 mRNA Expression in Endothelium by Euphorbia hirta L. through Its Beneficial β-Amyrin Molecule. Molecules 2014, 19, 10534-10545. https://doi.org/10.3390/molecules190710534
Shih MF, Cherng JY. Reduction of Adhesion Molecule Production and Alteration of eNOS and Endothelin-1 mRNA Expression in Endothelium by Euphorbia hirta L. through Its Beneficial β-Amyrin Molecule. Molecules. 2014; 19(7):10534-10545. https://doi.org/10.3390/molecules190710534
Chicago/Turabian StyleShih, Mei Fen, and Jong Yuh Cherng. 2014. "Reduction of Adhesion Molecule Production and Alteration of eNOS and Endothelin-1 mRNA Expression in Endothelium by Euphorbia hirta L. through Its Beneficial β-Amyrin Molecule" Molecules 19, no. 7: 10534-10545. https://doi.org/10.3390/molecules190710534