Discovery of Gramine Derivatives That Inhibit the Early Stage of EV71 Replication in Vitro

Abstract
:1. Introduction
2. Results
2.1. Antiviral Activity of Gramine and Its Derivatives

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Structural Formula | EC50 (µg/mL) a | CC50 (µg/mL) b | SI c | |||
|---|---|---|---|---|---|---|---|
| Vero | RD | Vero | RD | Vero | RD | ||
| Gramine |  | — d | — | 101.4 ± 5.6 e | 85.0 ± 16.3 | — | — |
| 4a |  | 12.7 ± 3.2 | 5.5 ± 2.4 | 32.6 ± 3.5 | 17.2 ± 1.6 | 2.6 | 3.1 |
| 4b |  | 5.1 ± 2.7 | 7.4 ± 1.1 | 18.6 ± 1.4 | 22.8 ± 1.7 | 3.6 | 3.1 |
| 4c |  | 5.6 ± 1.3 | 6.7 ± 0.8 | 22.4 ± 3.6 | 24.6 ± 3.5 | 4.0 | 3.7 |
| 4d |  | 9.2 ± 2.1 | 5.8 ± 1.4 | 37.8 ± 5.8 | 28.0 ± 5.2 | 4.1 | 4.8 |
| 4e |  | 7.3 ± 2.6 | 6.2 ± 2.8 | 40.7 ± 2.5 | 29.5 ± 3.3 | 5.6 | 4.8 |
| 4f |  7 7 | 14.4 ± 3.2 | — | 86.4 ± 7.4 | 44.0 ± 5.1 | 6.0 | — |
| 4g |  | 4.9 ± 0.7 | 4.9 ± 1.2 | 38.6 ± 5.7 | 32.0 ± 1.8 | 7.9 | 6.5 |
| 4h |  | 10.5 ± 2.1 | 11.2 ± 4.6 | 89.3 ± 12.6 | 52.2 ± 8.3 | 8.5 | 4.7 |
| 4i |  | 5.5 ± 1.6 | 6.1 ± 3.2 | 71.1 ± 3.8 | 65.2 ± 7.6 | 12.9 | 10.7 |
| 4j |  | 8.7 ± 2.9 | 8.3 ± 3.5 | 15.8 ± 3.3 | 17.8 ± 2.6 | 1.8 | 2.1 |
| 4k |  | 13.2 ± 1.3 | 9.8 ± 3.4 | 18.5 ± 4.8 | 27.6 ± 8.3 | 1.4 | 2.8 |
| 4l |  | — | — | 96.2 ± 15.5 | 65.2 ± 9.1 | — | — |
| 4m |  | — | — | >200 | >200 | — | — |
| 4n |  | — | — | 120.7 ± 20.5 | >200 | — | — |
| 4o |  | 16.9 ± 1.5 | 9.5 ± 1.7 | 72.8 ± 18.8 | 39.2 ± 11.6 | 4.3 | 4.1 |
| 4p |  | 2.1 ± 0.7 | 3.2 ± 0.4 | 6.6 ± 1.4 | 11.7 ± 4.6 | 3.1 | 3.7 |
| 4q |  | 1.8 ± 1.1 | 2.7 ± 1.2 | 10.8 ± 2.5 | 14.5 ± 3.6 | 6.0 | 5.4 |
| 4r |  | 9.9 ± 1.4 | 7.3 ± 1.3 | 132.5 ± 8.3 | 149.9 ± 16.2 | 13.4 | 20.5 |
| 4s |  | 7.6 ± 2.5 | 9.1 ± 3.1 | 108.7 ± 16.8 | 136.7 ± 9.3 | 14.3 | 15.0 |
| 4t |  | 1.9 ± 0.6 | 2.6 ± 0.8 | 10.5 ± 2.4 | 12.6 ± 3.5 | 5.5 | 4.8 |
| 4u |  | 2.3 ± 0.3 | 3.1 ± 1.3 | 4.9 ± 0.6 | 6.8 ± 2.5 | 2.1 | 2.2 |
| ribavirin f |  | 44.6 ± 13.5 | 32.1 | >500 | 435.8 | >11 | 13.6 |

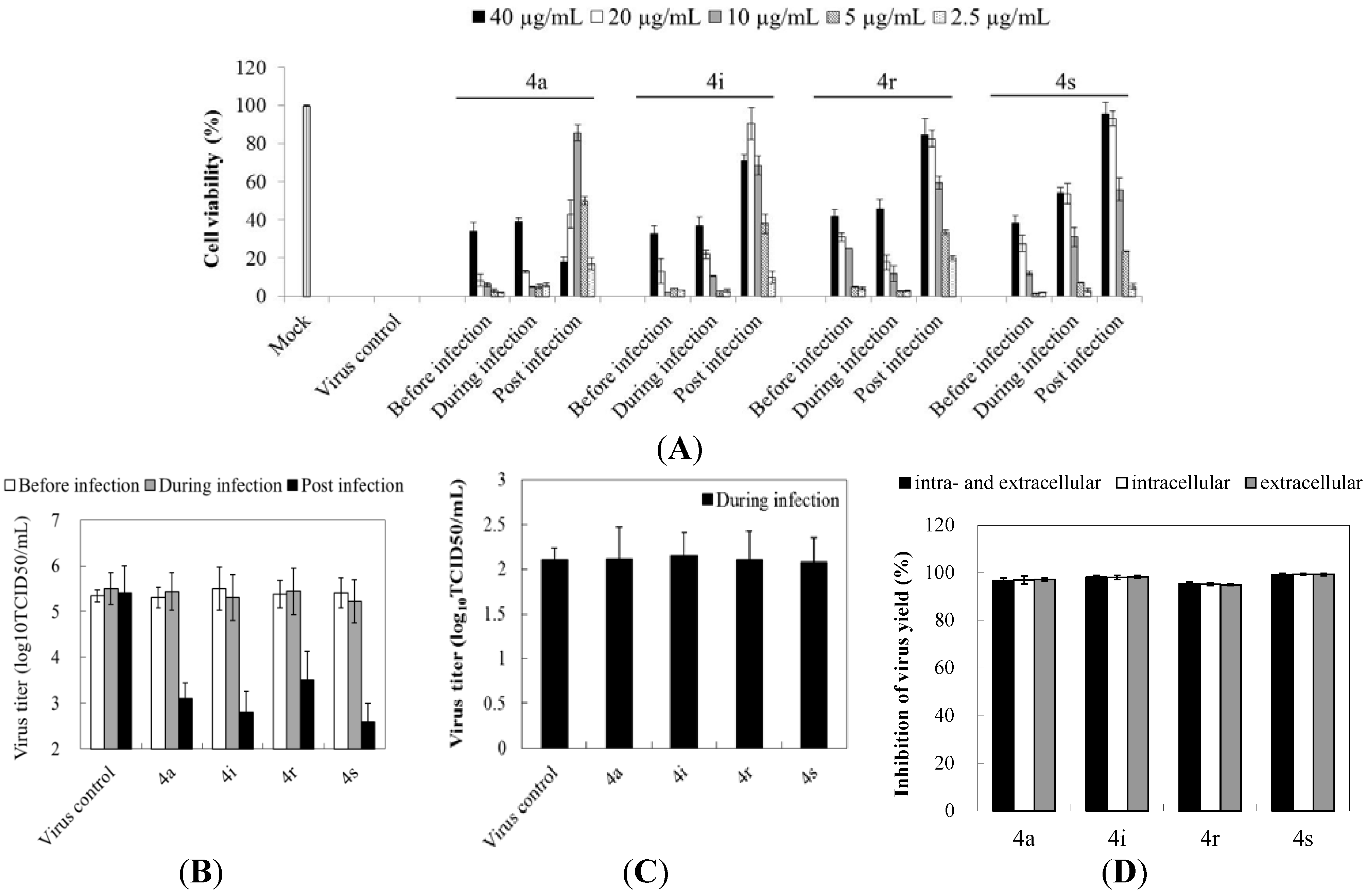
2.2. Preliminary Studies on Mechanisms of Action: Gramine Derivatives Mainly Target the Post-Infection Stage

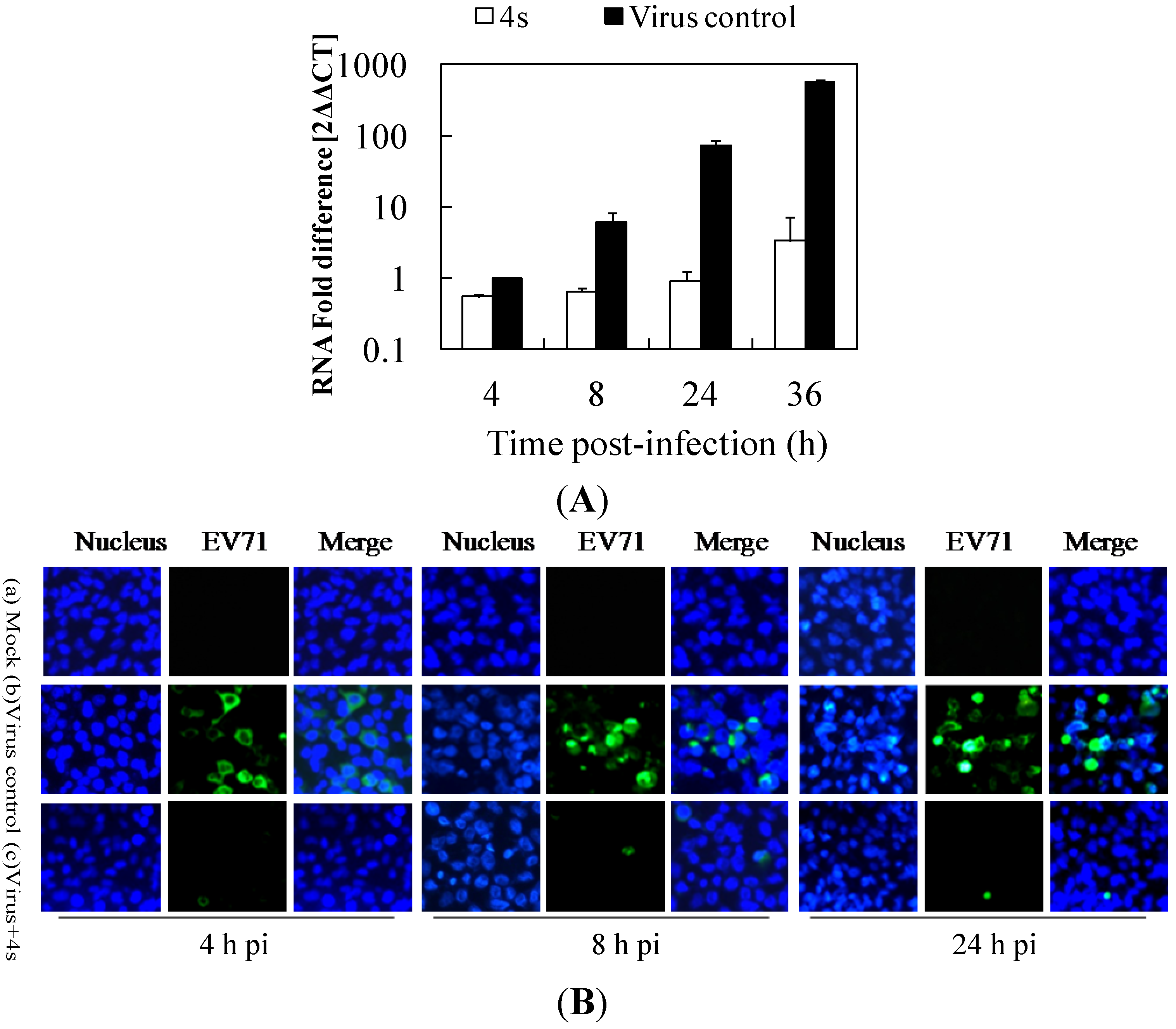
2.3. Gramine Derivatives Affect Viral Early Steps of Replication in Cells

2.4. Derivative 4s Inhibits Strongly Viral Replication in RD Cells

2.5. Compound 4s Inhibits EV71-Induced Apoptosis

3. Discussion
4. Experimental
4.1. Cells and Viruses
4.2. Preparation of the Tested Compounds
4.4. Determination of Cell Viability
4.5. Virus Yield Reduction Assay
4.6. Time-of-Addition Assay
4.7. RNA Extraction and Quantitative Reverse Transcription-PCR
4.8. Immunofluorescence Assay
4.9. Flow Cytometry Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schmidt, N.J.; Lennette, E.H.; Ho, H.H. An apparently new enterovirus isolated from patients with disease of the central nervous system. J. Infect. Dis. 1974, 129, 304–309. [Google Scholar] [CrossRef]
- AbuBakar, S.; Chee, H.Y.; Al-Kobaisi, M.F.; Jiang, X.S.; Chua, K.B.; Lam, S.K. Identification of enterovirus 71 isolates from an outbreak of hand, foot and mouth disease (HFMD) with fatal cases of encephalomyelitis in Malaysia. Virus Res. 1999, 61, 1–9. [Google Scholar] [CrossRef]
- McMinn, P.; Stratov, I.; Nagarajan, L.; Davis, S. Neurological manifestations of enterovirus 71 infection in children during an outbreak of hand, foot, and mouth disease in Western Australia. Clin. Infect. Dis. 2001, 32, 236–242. [Google Scholar] [CrossRef]
- Kehle, J.; Roth, B.; Metzger, C.; Pfitzner, A.; Enders, G. Molecular characterization of an Enterovirus 71 causing neurological disease in Germany. J. Neurovirol. 2003, 9, 126–128. [Google Scholar] [CrossRef]
- Hosoya, M.; Kawasaki, Y.; Sato, M.; Honzumi, K.; Kato, A.; Hiroshima, T.; Ishiko, H.; Suzuki, H. Genetic diversity of enterovirus 71 associated with hand, foot and mouth disease epidemics in Japan from 1983 to 2003. Pediatr. Infect. Dis. J. 2006, 25, 691–694. [Google Scholar]
- Bible, J.M.; Iturriza-Gomara, M.; Megson, B.; Brown, D.; Pantelidis, P.; Earl, P.; Ishiko, H.; Suzuki, H. Molecular epidemiology of human enterovirus 71 in the United Kingdom from 1998 to 2006. J. Clin. Microbiol. 2008, 46, 3192–3200. [Google Scholar] [CrossRef]
- Ho, M.; Chen, E.R.; Hsu, K.H.; Twu, S.J.; Chen, K.T.; Tsai, S.F.; Wang, J.R.; Shih, S.R. An epidemic of enterovirus 71 infection in Taiwan. N. Engl. J. Med. 1999, 341, 929–935. [Google Scholar] [CrossRef]
- Ding, N.Z.; Wang, X.M.; Sun, S.W.; Song, Q.; Li, S.N.; He, C.Q. Appearance of mosaic enterovirus 71 in the 2008 outbreak of China. Virus Res. 2009, 145, 157–161. [Google Scholar] [CrossRef]
- Li, J.; Huo, X.X.; Dai, Y.; Yang, Z.H.; Lei, Y.K.; Jiang, Y.Z.; Li, G.; Zhan, J.; Zhan, F. Evidences for intertypic and intratypic recombinant events in EV71 of hand, foot and mouth disease during an epidemic in Hubei Province, China, 2011. Virus Res. 2012, 16, 195–202. [Google Scholar]
- Chang, L.Y.; Huang, L.M.; Gau, S.S.; Wu, Y.Y.; Hsia, S.H.; Fan, T.Y.; Lin, K.L.; Huang, Y.C.; Lu, C.Y.; Lin, T.Y. Neurodevelopment and cognition in children after enterovirus 71 infection. N. Engl. J. Med. 2007, 356, 1226–1234. [Google Scholar]
- Xu, J.; Qian, Y.; Wang, S.X.; Serrano, J.M.; Li, W.; Huang, Z.H.; Lu, S. EV71: An emerging infectious disease vaccine target in the Far East? Vaccine 2010, 28, 3516–3521. [Google Scholar] [CrossRef]
- Shang, L.Q.; Xu, M.Y.; Yin, Z. Antiviral drug discovery for the treatment of entero-virus 71 infections. Antivir. Res. 2013, 97, 183–194. [Google Scholar]
- Li, Z.H.; Li, C.M.; Ling, P.; Shen, F.H.; Chen, S.H.; Liu, C.C.; Yu, C.K.; Chen, S.H. Ribavirin reduces mortality in enterovirus 71-infected mice by decreasing viral replication. J. Infect. Dis. 2008, 197, 854–857. [Google Scholar]
- Liu, M.L.; Lee, Y.P.; Wang, Y.F.; Lei, H.Y.; Liu, C.C.; Wang, S.M.; Su, I.J.; Wang, J.R.; Yeh, T.M.; Chen, S.H.; et al. Type I interferons protect mice against enterovirus 71 infection. J. Gen. Virol. 2005, 86, 3263–3269. [Google Scholar] [CrossRef]
- Pevear, D.C.; Tull, T.M.; Seipel, M.E.; Groarke, J.M. Activity of pleconaril against enteroviruses. Antimicrob. Agents Chemother. 1999, 43, 2109–2115. [Google Scholar]
- Iwata, S.; Saito, S.; Kon-ya, K.; Shizuri, Y.; Ohizumi, Y. Novel marine-derived halogen-containing gramine analogues induce vasorelaxation in isolated rat aorta. Eur. J. Pharmacol. 2001, 432, 63–70. [Google Scholar]
- Nakahata, N.; Harada, Y.; Tsuji, M.; Kon-ya, K.; Shizuri, Y.; Ohizumi, Y. Structure-activity relationship of gramine derivatives in Ca2q release from sarcoplasmic reticulum. Eur. J. Pharmacol. 1999, 382, 129–132. [Google Scholar] [CrossRef]
- Grün, S.; Frey, M.; Gierl, A. Evolution of the indole alkaloid biosynthesis in the genus Hordeum: Distribution of gramine and DIBOA and isolation of the benzoxazinoid biosynthesis genes from Hordeum lechleri. Phytochemistry 2005, 66, 1264–1272. [Google Scholar] [CrossRef]
- Katritzky, A.R.; Khelashvili, L.; Munawar, M.A. Syntheses of IAA-and IPA-Amino Acid Conjugates. J. Org. Chem. 2008, 73, 9171–9173. [Google Scholar] [CrossRef]
- Zheng, M.F.; Zheng, M.Y.; Ye, D.J.; Deng, Y.M.; Qiu, S.F.; Luo, X.M.; Chen, K.; Liu, H.; Jiang, H. Indole derivatives as potent inhibitors of 5-lipoxygenase: Design, synthesis, biological evaluation, and molecular modeling. Bioorg. Med. Chem. Lett. 2007, 17, 2414–2420. [Google Scholar] [CrossRef]
- Giampieri, M.; Balbi, A.; Mazzei, M.; Colla, P.L.; Ibba, C.; Loddo, R. Antiviral activity of indole derivatives. Antiviral Res. 2009, 83, 179–185. [Google Scholar] [CrossRef]
- Wang, R.; Shi, H.F.; Zhao, J.F.; He, Y.P.; Zhang, H.B.; Liu, J.P. Design, synthesis and aromatase inhibitory activities of novel indole-imidazole derivatives. Bioorg. Med. Chem. Lett. 2013, 23, 1760–1762. [Google Scholar] [CrossRef]
- Ke, S.Y.; Shi, L.Q.; Cao, X.F.; Yang, Q.Y.; Liang, Y.; Yang, Z.W. Heterocycle- functional gramine analogues: Solvent- and catalyst-free synthesis and their inhibition activities against cell proliferation. Eur. J. Med. Chem. 2012, 54, 248–254. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar]
- Weng, K.F.; Chen, L.L.; Huang, P.N.; Shih, S.R. Neural pathogenesis of enterovirus71 infection. Microbes Infect. 2010, 12, 505–510. [Google Scholar] [CrossRef]
- Shih, S.R.; Weng, K.F.; Stollar, V.; Li, M.L. Viral protein synthesis is required for Enterovirus71 to induce apoptosis in human glioblastoma cells. J. Neurovirol. 2008, 14, 53–61. [Google Scholar] [CrossRef]
- Shih, S.R.; Stollar, V.; Li, M.L. Host Factors in Enterovirus 71 Replication. J. Virol. 2011, 85, 9658–9666. [Google Scholar] [CrossRef]
- Chen, T.C.; Weng, K.F.; Chang, S.C.; Lin, J.Y.; Huang, P.N.; Shih, S.R. Development of antiviral agents for enteroviruses. J. Antimicrob. Chemother. 2008, 62, 1169–1173. [Google Scholar] [CrossRef]
- MicrobiologyBytes. Available online: http://www.microbiologybytes.com/virology/Picornaviruses.html (accessed on 17 August 2013).
- Shih, S.R.; Tsai, K.N.; Li, Y.S.; Chueh, C.C.; Chan, E.C. Inhibition of Enterovirus 71-Induced Apoptosis by Allophycocyanin Isolated From a Blue-Green Alga Spirulina platensis. J. Med. Virol. 2003, 70, 119–125. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds 4a to 4u are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Shi, L.; Wang, K.; Liu, M.; Yang, Q.; Yang, Z.; Ke, S. Discovery of Gramine Derivatives That Inhibit the Early Stage of EV71 Replication in Vitro. Molecules 2014, 19, 8949-8964. https://doi.org/10.3390/molecules19078949
Wei Y, Shi L, Wang K, Liu M, Yang Q, Yang Z, Ke S. Discovery of Gramine Derivatives That Inhibit the Early Stage of EV71 Replication in Vitro. Molecules. 2014; 19(7):8949-8964. https://doi.org/10.3390/molecules19078949
Chicago/Turabian StyleWei, Yanhong, Liqiao Shi, Kaimei Wang, Manli Liu, Qingyu Yang, Ziwen Yang, and Shaoyong Ke. 2014. "Discovery of Gramine Derivatives That Inhibit the Early Stage of EV71 Replication in Vitro" Molecules 19, no. 7: 8949-8964. https://doi.org/10.3390/molecules19078949