Transcriptional Responses of the Bdtf1-Deletion Mutant to the Phytoalexin Brassinin in the Necrotrophic Fungus Alternaria brassicicola

 ,
,
Abstract
:1. Introduction
2. Results
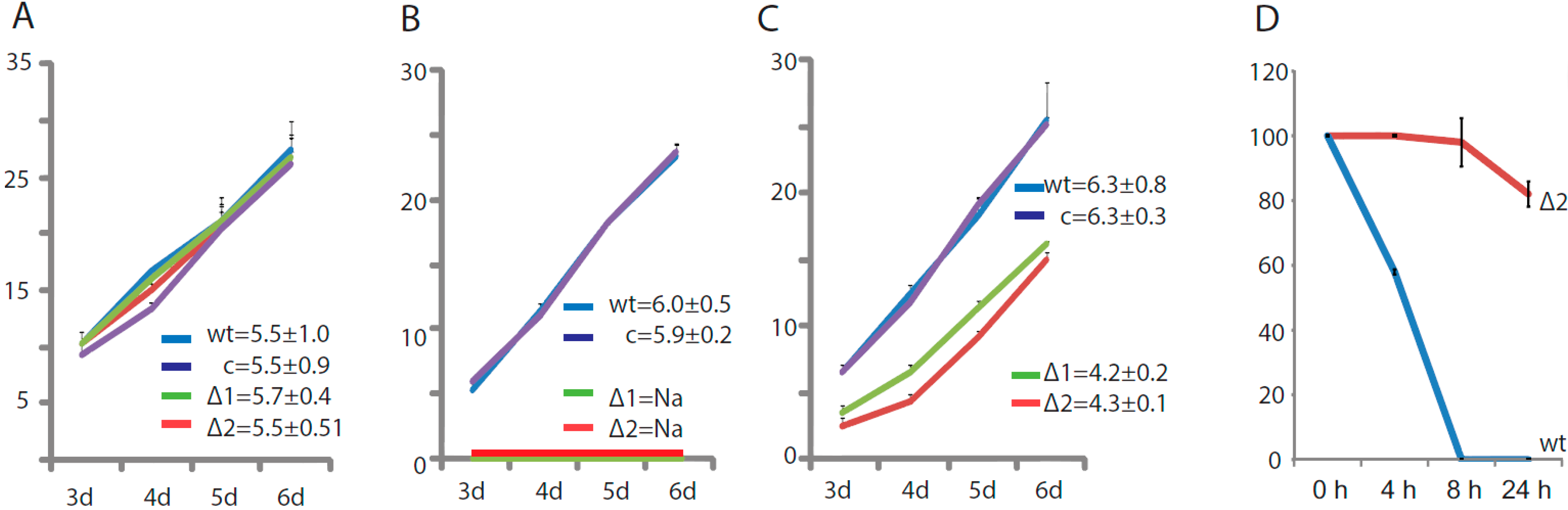
2.1. Effects of Brassinin on ∆bdtf1 Mycelium

2.2. Gene Expression during Plant Infection
2.3. Statistics of Gene Expression Profiles
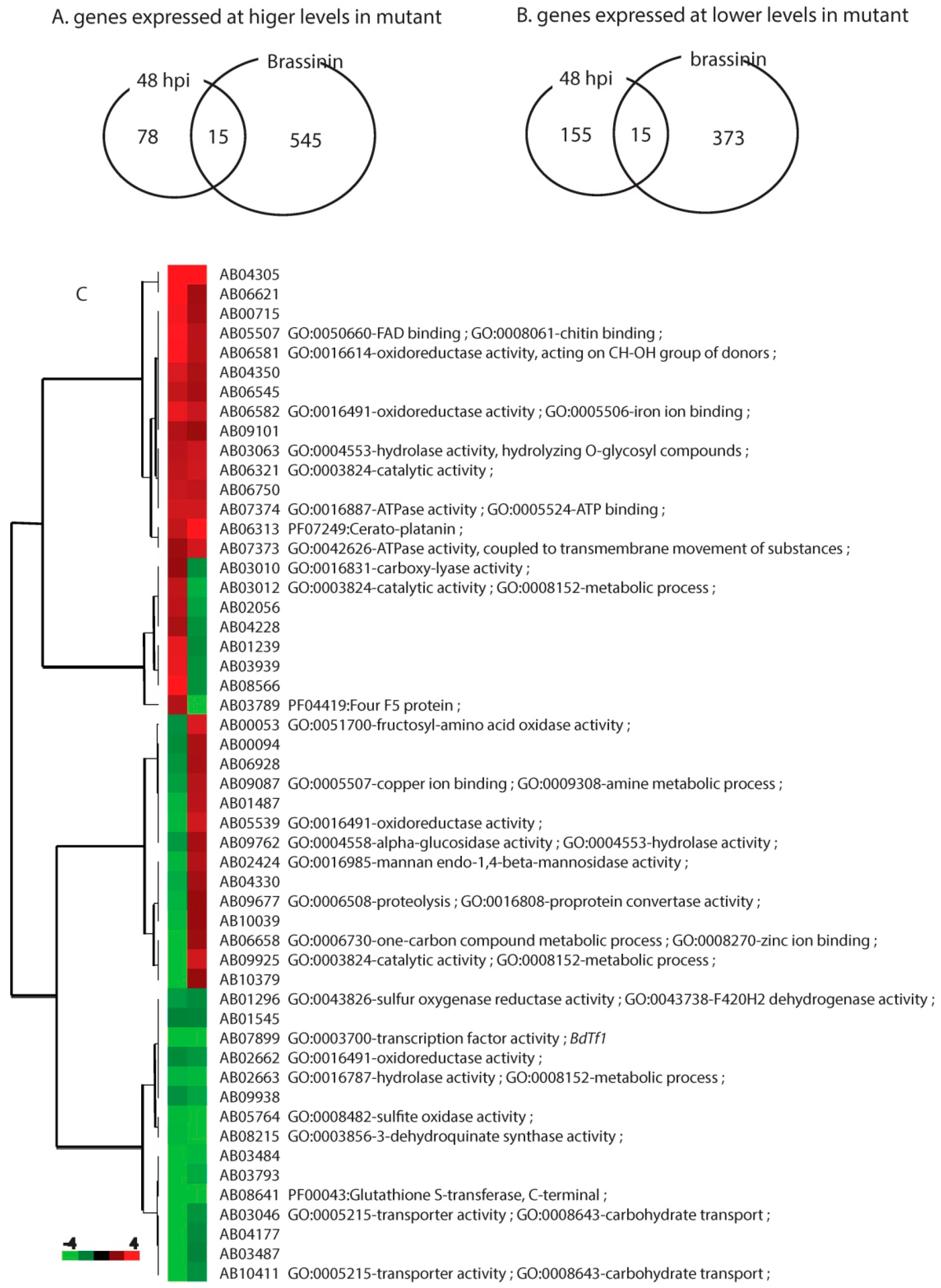
2.4. Brassinin Effects: Genes at Lower Levels in the Mutant

{kind=link}
{kind=link}
| Annotation | Description | p-value |
|---|---|---|
| GO:0043141 | ATP-dependent 5'-3' DNA helicase activity | 0.000191 |
| GO:0008026 | ATP-dependent helicase activity | 0.000191 |
| GO:0008758 | UDP-2,3-diacylglucosamine hydrolase activity | 0.000191 |
| GO:0047429 | nucleoside-triphosphate diphosphatase activity | 0.000191 |
| GO:0004787 | thiamin-pyrophosphatase activity | 0.000191 |
| GO:0008413 | 8-oxo-7,8-dihydroguanine triphosphatase activity | 0.000191 |
| GO:0004386 | helicase activity | 0.000191 |
| GO:0019176 | dihydroneopterin monophosphate phosphatase activity | 0.000191 |
| GO:0019177 | dihydroneopterin triphosphate pyrophosphohydrolase activity | 0.000191 |
| GO:0008828 | dATP pyrophosphohydrolase activity | 0.000191 |
| GO:0000810 | diacylglycerol pyrophosphate phosphatase activity | 0.000191 |
| GO:0043139 | 5'-3' DNA helicase activity | 0.000191 |
| GO:0005488 | binding | 0.000213 |
| GO:0030554 | adenyl nucleotide binding | 0.000213 |
| GO:0017110 | nucleoside-diphosphatase activity | 0.000213 |
| GO:0008796 | bis(5'-nucleosyl)-tetraphosphatase activity | 0.000312 |
| GO:0004551 | nucleotide diphosphatase activity | 0.000312 |
| GO:0000166 | nucleotide binding | 0.000587 |
| GO:0003678 | DNA helicase activity | 0.000629 |
| GO:0004003 | ATP-dependent DNA helicase activity | 0.000629 |
| GO:0003676 | nucleic acid binding | 0.000772 |
| GO:0017076 | purine nucleotide binding | 0.001009 |
| GO:0008094 | DNA-dependent ATPase activity | 0.001162 |
| GO:0003824 | catalytic activity | 0.002675 |
| GO:0017171 | serine hydrolase activity | 0.004091 |
| GO:0004086 | carbamoyl-phosphate synthase activity | 0.004091 |
| GO:0008236 | serine-type peptidase activity | 0.004091 |
| GO:0016887 | ATPase activity | 0.01031 |
| GO:0005730 | nucleolus | 0.010587 |
| GO:0050660 | FAD binding | 0.010587 |
| GO:0048037 | cofactor binding | 0.012798 |
| GO:0032040 | small subunit processome | 0.013819 |
| GO:0031177 | phosphopantetheine binding | 0.020373 |
| GO:0017111 | nucleoside-triphosphatase activity | 0.029526 |
| GO:0016638 | oxidoreductase activity, acting on the CH-NH2 group of donors | 0.031008 |
| GO:0005524 | ATP binding | 0.037275 |
| GO:0032559 | adenyl ribonucleotide binding | 0.037275 |
| GO:0016874 | ligase activity | 0.042934 |
| GO:0006537 | glutamate biosynthetic process | 0.043581 |
| GO:0042623 | ATPase activity, coupled | 0.049164 |
2.5. Brassinin Effects: Genes at Higher Levels in the Mutant
| Annotation | Description | Representation | p-value |
|---|---|---|---|
| GO:0009987 | cellular process | Under | 0.001924 |
| GO:0044237 | cellular metabolic process | Under | 0.013208 |
| KOG0065 | Pleiotropic drug resistance proteins (PDR1-15), ABC superfamily | Over | 0.041251 |
| PF06422 | CDR ABC transporter | Over | 0.044766 |
2.6. Gene Expression Patterns During Plant Infection
| Annotation | Description | p-value |
|---|---|---|
| GO:0016798 | hydrolase activity acting on glycosyl bonds | 7.38 × 10−9 |
| GO:0004553 | hydrolase activity, hydrolyzing O-glycosyl compounds | 7.38 × 10−9 |
| GO:0005975 | carbohydrate metabolic process | 5.80 × 10−8 |
| GO:0005622 | intracellular | 9.67 × 10−4 |
| GO:0044424 | intracellular part | 0.025823 |
| GO:0006139 | nucleobase, nucleoside, nucleotide and nucleic acid metabolic process | 0.025823 |
2.7. Fungal Genes Affected Under Both Conditions
3. Discussion
3.1. Effects of Brassinin on Protein Synthesis

| ProteinID | HMM-Secretion | 2 Mycelial Growth with Brassinin | 3 Plant Infection | Go Annotation | Manual Blast Results | ||
|---|---|---|---|---|---|---|---|
| Wild type | ∆bdtf1 | Wild type | ∆bdtf1 | ||||
| AB01296.1 | 10.3 | 4.5 | 3.1 | 1.4 | GO:0043826-sulfur oxygenase reductase activity; | Aldo/keto reductase are major group of enzymes involved in detoxification | |
| AB01545.1 | 199.8 | 99.0 | 195.2 | 95.9 | NA | xanthine phosphoribosyltransferase or purine salvage enzyme | |
| AB02662.1 | 199.2 | 93.9 | 95.3 | 41.9 | GO:0016491-oxidoreductase activity; | Aldo/keto reductase involved in detoxification | |
| AB02663.1 | 1 S | 100.7 | 37.0 | 33.0 | 10.4 | GO:0016787-hydrolase activity ; | HAD-superfamily subfamily IIA hydrolase |
| AB03046.1 | 94.4 | 17.6 | 177.8 | 80.5 | GO:0005215-transporter activity; | Sugar transporter STL1 induced when cells are subjected to osmotic shock | |
| AB03484.1 | 75.9 | 13.3 | 75.6 | 25.6 | NA | Similar to glutathione-dependent formaldehyde-activating GFA | |
| AB03487.1 | 3.9 | 0.7 | 20.9 | 10.3 | NA | Glutathione S-transferase omega-like | |
| AB03793.1 | 4.6 | 1.1 | 54.3 | 21.8 | NA | Cupin domain, salicylate hydroxylase | |
| AB04177.1 | 5.3 | 1.1 | 73.6 | 35.1 | NA | Methyltransferase involved in epigenetic regulation. | |
| AB05764.1 | 3.0 | 0.9 | 1.4 | 0.3 | GO:0008482-sulfite oxidase activity; | nitrate reductase | |
| AB07899.1 | 11.0 | 0.0 | 29.7 | 0.1 | GO:0003700-transcription factor activity; | Bdtf1 | |
| AB08215.1 | 5.1 | 1.7 | 56.7 | 8.5 | GO:0003856-3-dehydroquinate synthase activity; | 3-dehydroquinate synthase | |
| AB08641.1 | 18.7 | 0.4 | 185.1 | 15.2 | NA | glutathione S-transferase | |
| AB09938.1 | 129.7 | 59.7 | 61.0 | 24.8 | NA | arginine N-methyltransferase | |
| AB10411.1 | 72.5 | 12.4 | 110.7 | 50.9 | GO:0005215-transporter activity; | Sugar transporter STL1 | |
3.2. Genes Important for Cell Protection
3.3. Compensatory Genes
3.4. Enzymatic Modification of Brassinin
3.5. Candidates for Brassinin-Detoxifying Enzymes
4. Experimental Section
4.1. Fungal Strains and their Maintenance
4.2. Assays for Brassinin Digestion and Preparation of Mycelium for RNA-Seq
4.3. Preparation of Fungal Tissues from Infected Host Plants
4.4. Generation of RNA-Seq Data
Representation Analysis of Functional Annotation Terms
4.5. qRT-PCR
5. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Takasugi, M.; Katsui, N.; Shirata, A. Isolation of three novel sulphur-containing phytoalexins from the chinese cabbage Brassica campestris L. ssp. pekinensis (cruciferae). J. Chem. Soc. Chem. Commun. 1986, 1077–1078. [Google Scholar] [CrossRef]
- Pedras, M.S.; Chumala, P.B.; Jin, W.; Islam, M.S.; Hauck, D.W. The phytopathogenic fungus Alternaria brassicicola: Phytotoxin production and phytoalexin elicitation. Phytochemistry 2009, 70, 394–402. [Google Scholar] [CrossRef]
- Govrin, E.M.; Levine, A. Infection of Arabidopsis with a necrotrophic pathogen, Botrytis cinerea, elicits various defense responses but does not induce systemic acquired resistance (SAR). Plant Mol. Biol. 2002, 48, 267–276. [Google Scholar] [CrossRef]
- Sellam, A.; Iacomi-Vasilescu, B.; Hudhomme, P.; Simoneau, P. In vitro antifungal activity of brassinin, camalexin and two isothiocyanates against the crucifer pathogens Alternaria brassicicola and Alternaria brassicae. Plant Pathol. 2007, 56, 296–301. [Google Scholar] [CrossRef]
- Sellam, A.; Dongo, A.; Guillemette, T.; Hudhomme, P.; Simoneau, P. Transcriptional responses to exposure to the brassicaceous defence metabolites camalexin and allyl-isothiocyanate in the necrotrophic fungus Alternaria brassicicola. Mol. Plant. Pathol. 2007, 8, 195–208. [Google Scholar] [CrossRef]
- Joubert, A.; Bataille-Simoneau, N.; Campion, C.; Guillemette, T.; Hudhomme, P.; Iacomi-Vasilescu, B.; Leroy, T.; Pochon, S.; Poupard, P.; Simoneau, P. Cell wall integrity and high osmolarity glycerol pathways are required for adaptation of Alternaria brassicicola to cell wall stress caused by brassicaceous indolic phytoalexins. Cell Microbiol. 2011, 13, 62–80. [Google Scholar] [CrossRef] [Green Version]
- Rogers, E.E.; Glazebrook, J.; Ausubel, F.M. Mode of action of the Arabidopsis thaliana phytoalexin camalexin and its role in Arabidopsis-pathogen interactions. Mol. Plant. Microbe Interact. 1996, 9, 748–757. [Google Scholar] [CrossRef]
- Pedras, M.S.; Ahiahonu, P.W.; Hossain, M. Detoxification of the cruciferous phytoalexin brassinin in Sclerotinia sclerotiorum requires an inducible glucosyltransferase. Phytochemistry 2004, 65, 2685–2694. [Google Scholar] [CrossRef]
- Pedras, M.S.; Minic, Z.; Jha, M. Brassinin oxidase, a fungal detoxifying enzyme to overcome a plant defense—Purification, characterization and inhibition. FEBS J. 2008, 275, 3691–3705. [Google Scholar] [CrossRef]
- Pedras, M.S.; Yaya, E.E.; Glawischnig, E. The phytoalexins from cultivated and wild crucifers: Chemistry and biology. Nat. Prod. Rep. 2011, 28, 1381–1405. [Google Scholar] [CrossRef]
- Srivastava, A.; Cho, I.K.; Cho, Y. The Bdtf1 gene in Alternaria brassicicola is important in detoxifying brassinin and maintaining virulence on Brassica species. Mol. Plant. Microbe Interact. 2013, 26, 1429–1440. [Google Scholar] [CrossRef]
- Pedras, M.S.; Minic, Z.; Sarma-Mamillapalle, V.K. Substrate specificity and inhibition of brassinin hydrolases, detoxifying enzymes from the plant pathogens Leptosphaeria maculans and Alternaria brassicicola. FEBS J. 2009, 276, 7412–7428. [Google Scholar] [CrossRef]
- Cho, Y.; Davis, J.W.; Kim, K.H.; Wang, J.; Sun, Q.H.; Cramer, R.A.J.; Lawrence, C.B. A high throughput targeted gene disruption method for Alternaria brassicicola functional genomics using linear minimal element (LME) constructs. Mol. Plant. Microbe Interact. 2006, 19, 7–15. [Google Scholar] [CrossRef]
- Ohm, R.A.; Feau, N.; Henrissat, B.; Schoch, C.L.; Horwitz, B.A.; Barry, K.W.; Condon, B.J.; Copeland, A.C.; Dhillon, B.; Glaser, F.; et al. Diverse lifestyles and strategies of plant pathogenesis encoded in the genomes of eighteen Dothideomycetes fungi. PLoS Pathog. 2012, 8, e1003037. [Google Scholar] [CrossRef] [Green Version]
- Pedras, M.S.; Minic, Z. Differential protein expression in response to the phytoalexin brassinin allows the identification of molecular targets in the phytopathogenic fungus Alternaria brassicicola. Mol. Plant Pathol 2012, 13, 483–493. [Google Scholar] [CrossRef]
- Pedras, M.S.; Minic, Z.; Sarma-Mamillapalle, V.K. Synthetic inhibitors of the fungal detoxifying enzyme brassinin oxidase based on the phytoalexin camalexin scaffold. J. Agric. Food Chem. 2009, 57, 2429–2435. [Google Scholar] [CrossRef]
- Coleman, J.J.; Wasmann, C.C.; Usami, T.; White, G.J.; Temporini, E.D.; McCluskey, K.; VanEtten, H.D. Characterization of the gene encoding pisatin demethylase (FoPDA1) in Fusarium oxysporum. Mol. Plant. Microbe Interact. 2011, 24, 1482–1491. [Google Scholar] [CrossRef]
- Schafer, W.; Straney, D.; Ciuffetti, L.; VanEtten, H.D.; Yoder, O.C. One enzyme makes a fungal pathogen, but not a saprophyte, virulent on a new host plant. Science 1989, 246, 247–249. [Google Scholar]
- Pedras, M.S.; Hossain, S.; Snitynsky, R.B. Detoxification of cruciferous phytoalexins in Botrytis cinerea: Spontaneous dimerization of a camalexin metabolite. Phytochemistry 2011, 72, 199–206. [Google Scholar]
- Pedras, M.S.; Ahiahonu, P.W. Metabolism and detoxification of phytoalexins and analogs by phytopathogenic fungi. Phytochemistry 2005, 66, 391–411. [Google Scholar] [CrossRef]
- Cho, Y.; Kim, K.H.; la Rota, M.; Scott, D.; Santopietro, G.; Callihan, M.; Lawrence, C.B. Identification of virulence factors by high throughput targeted gene deletion of regulatory genes in Alternaria brassicicola. Mol. Microbiol. 2009, 72, 1316–1333. [Google Scholar] [CrossRef]
- Cho, Y.; Ohm, R.A.; Grigoriev, I.V.; Srivastava, A. Fungal-specific transcription factor AbPf2 activates pathogenicity in Alternaria brassicicola. Plant J. 2013, 75, 498–514. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nordberg, H.; Shabalov, I.; Aerts, A.; Cantor, M.; Goodstein, D.; Kuo, A.; Minovitsky, S.; Nikitin, R.; Ohm, R.A.; et al. The genome portal of the Department of Energy Joint Genome Institute. Nucleic Acids Res. 2012, 40, D26–32. [Google Scholar] [CrossRef]
- JGI fungal portal MycoCosm. Available online: http://jgi.doe.gov/Abrassicicola (accessed on 9 July 2014).
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Cho, Y.; Srivastava, A.; Ohm, R.A.; Lawrence, C.B.; Wang, K.H.; Grigoriev, I.V.; Marahatta, S.P. Transcription factor Amr1 induces melanin biosynthesis and suppresses virulence in Alternaria brassicicola. PLoS Pathog. 2012, 8, e1002974. [Google Scholar] [CrossRef]
- Srivastava, A.; Ohm, R.A.; Oxiles, L.; Brooks, F.; Lawrence, C.B.; Grigoriev, I.V.; Cho, Y. A zinc-finger-family transcription factor, AbVf19, is required for the induction of a gene subset important for virulence in Alternaria brassicicol. Mol. Plant Microbe Interact. 2012, 25, 443–452. [Google Scholar] [CrossRef]
- Sample availability: Contact YC, E-Mail: [email protected]; Tel.: +82-43-240-6255; Fax: +82-43-240-6259.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cho, Y.; Ohm, R.A.; Devappa, R.; Lee, H.B.; Grigoriev, I.V.; Kim, B.Y.; Ahn, J.S. Transcriptional Responses of the Bdtf1-Deletion Mutant to the Phytoalexin Brassinin in the Necrotrophic Fungus Alternaria brassicicola. Molecules 2014, 19, 10717-10732. https://doi.org/10.3390/molecules190810717
Cho Y, Ohm RA, Devappa R, Lee HB, Grigoriev IV, Kim BY, Ahn JS. Transcriptional Responses of the Bdtf1-Deletion Mutant to the Phytoalexin Brassinin in the Necrotrophic Fungus Alternaria brassicicola. Molecules. 2014; 19(8):10717-10732. https://doi.org/10.3390/molecules190810717
Chicago/Turabian StyleCho, Yangrae, Robin A. Ohm, Rakshit Devappa, Hyang Burm Lee, Igor V. Grigoriev, Bo Yeon Kim, and Jong Seog Ahn. 2014. "Transcriptional Responses of the Bdtf1-Deletion Mutant to the Phytoalexin Brassinin in the Necrotrophic Fungus Alternaria brassicicola" Molecules 19, no. 8: 10717-10732. https://doi.org/10.3390/molecules190810717