Comprehensive Phenolic Profiling of Cyclopia genistoides (L.) Vent. by LC-DAD-MS and -MS/MS Reveals Novel Xanthone and Benzophenone Constituents


Abstract
:1. Introduction
2. Results and Discussion
2.1. HPLC-DAD Method Development

2.2. LC-DAD-ESI-MS and -MS/MS Identification of Compounds Present in Hot Water Extracts of Unfermented and Fermented C. genistoides

2.2.1. Benzophenone Derivatives

{kind=link}
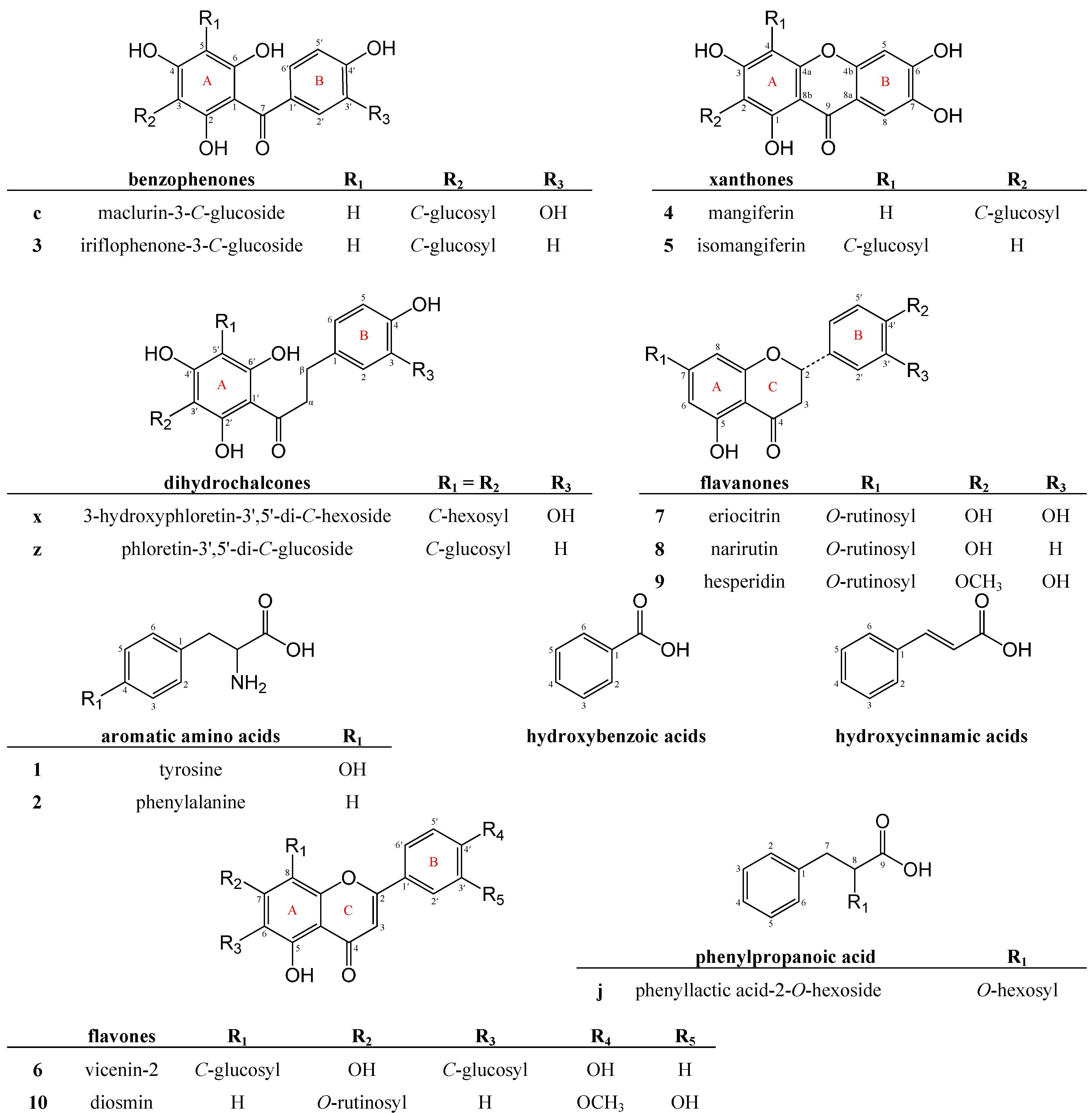{kind=link}
{kind=link}
{kind=link}
| Nr | tR (min) | Proposed Compound | λmax (nm) | Mode | Accurate Mass, exp. | Proposed Formula | Error (ppm) | Precursor Ion | LC-MS/MS Ions a,b |
|---|---|---|---|---|---|---|---|---|---|
| a | 4.06 | maclurin-di-O,C-hexoside | 235, 290, 320 sh | + | 587.1625 | C25H31O16 | −2.2 | 587 | 425, 407, 389, 371, 353, 341, 329, 305, 287, 275, 261, 243, 231, 219, 195, 177, 165, 153, 137, 121 |
| − | 585.1469 | C25H29O16 | 2.2 | 585 | 495, 465, 385, 355, 333, 303, 285, 261, 223, 193, 165, 125 | ||||
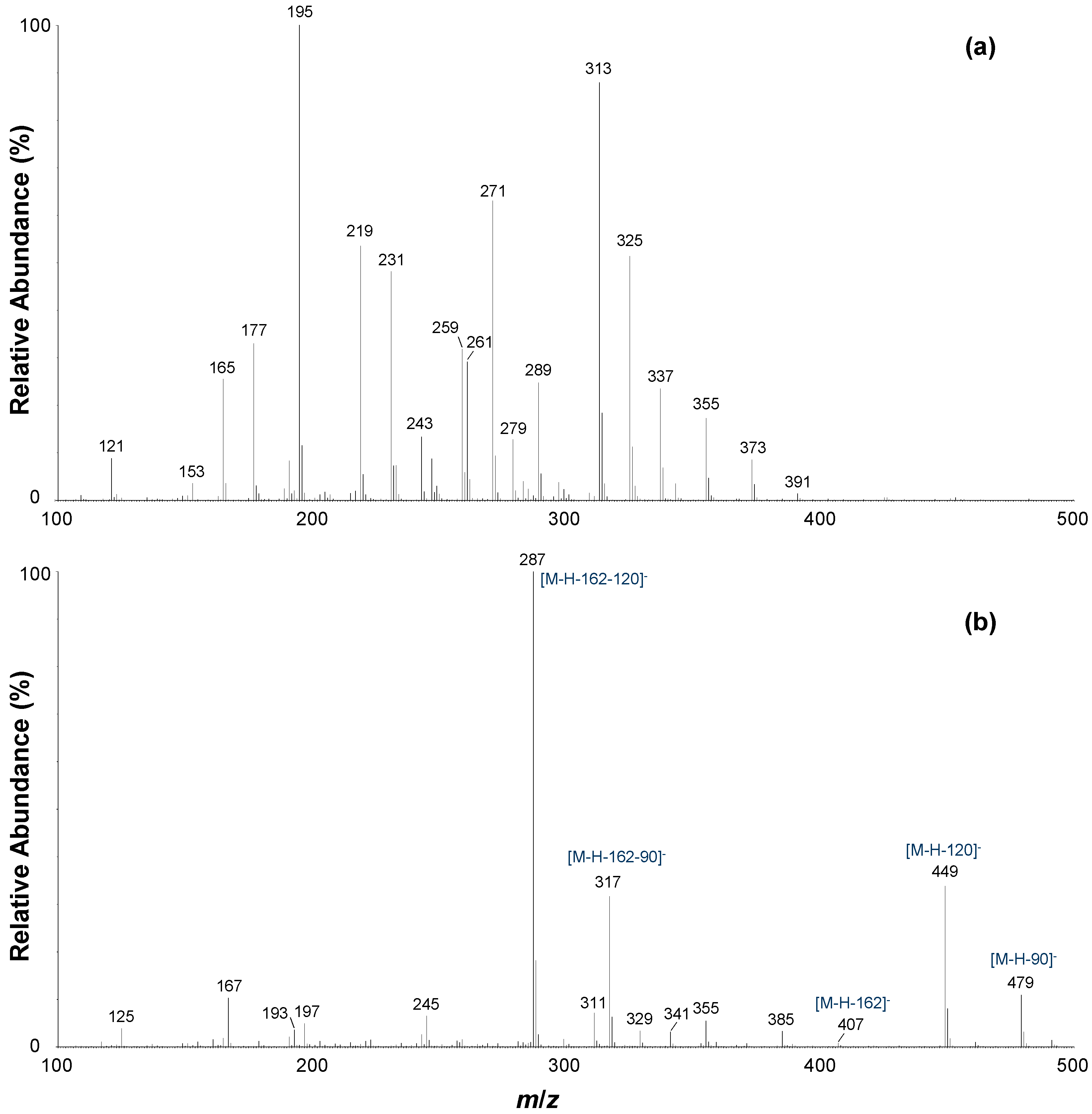
| b | 7.24 | iriflophenone-di-O,C-hexoside isomer | 234, 294 | + | 571.1664 | C25H31O15 | 0.2 | 571 | 373, 355, 337, 325, 313, 289, 279, 271, 261, 243, 231, 219, 195, 177, 165, 121 |
| − | 569.1509 | C25H29O15 | 0.5 | 569 | 479, 449, 407, 385, 355, 341, 329, 317, 311, 287, 245, 197, 193, 167, 125 | ||||
| c | 8.17 | maclurin-3-C-glucoside | 235, 290, 318 sh | + | 425.1080 | C19H21O11 | −0.4 | 425 | 353, 341, 329, 287, 261, 243, 231, 219, 195, 177, 165, 153, 137, 121 |
| − | 423.0923 | C19H19O11 | −0.9 | 423 | 333, 303, 261, 223, 205, 193, 165, 151, 137, 125, 109 | ||||
| d | 10.49 | iriflophenone-di-O,C-hexoside isomer | 225, 290 | + | 571.1503 | C25H31O15 | −5.6 | 571 | 373, 337, 325, 313, 289, 279, 271, 261, 243, 231, 219, 195, 177, 165, 121 |
| − | 569.1511 | C25H29O15 | 0.9 | 569 | 479, 449, 385, 355, 317, 287, 245, 193, 167, 125 | ||||
| f | 13.27 | iriflophenone-di-O,C-hexoside isomer | 225, 290 | + | 571.1639 | C25H31O15 | −4.2 | 571 | 425, 355, 337, 325, 313, 289, 279, 271, 261, 247, 231, 219, 195, 177, 165, 121 |
| − | 569.1503 | C25H29O15 | −0.5 | 569 | 479, 449, 385, 355, 317, 287, 245, 193, 167, 125 | ||||
| 3 | 13.70 | iriflophenone-3-C-glucoside | 235, 295 | + | 409.1133 | C19H21O10 | −0.5 | 409 | 337, 325, 313, 279, 271, 261, 243, 231, 219, 195, 177, 165, 153, 121 |
| − | 407.0967 | C19H19O10 | −2.7 | 407 | 317, 299, 287, 257, 245, 215, 201, 193, 167, 151, 137, 125 | ||||
| i | 16.65 | iriflophenone-di-C-hexoside c | 225, 302 (weak) | + | 571.1655 | C25H31O15 | −1.4 | 571 | 481, 470, 463, 451, 433, 421, 403, 391, 379, 367, 355, 349, 337, 325, 313, 295, 285, 273, 261, 243, 231, 219, 189, 177, 121 |
| − | 569.1509 | C25H29O15 | 0.5 | 569 | 479, 461, 431, 389, 359, 329, 317, 287, 239, 167 |

2.2.2. Xanthone Derivatives
| Nr | tR (min) | Proposed Compound | λmax (nm) | Mode | Accurate Mass, exp. | Proposed Formula | Error (ppm) | Precursor Ion | LC-MS/MS Ions a,b |
|---|---|---|---|---|---|---|---|---|---|
| g | 16.28 | tetrahydroxyxanthone-C-hexoside dimer c | 259, 317, 365 | + | 843.1610 | C38H35O22 | −0.5 | 843 | 843, 827, 807, 789, 772, 759, 743, 729, 711, 705, 693, 687, 675, 669, 657, 651, 639, 627, 603, 598, 585, 573, 562, 555, 531, 479, 425, 417 |
| − | 841.1494 | C38H33O22 | 0.5 | 841, CE = 45 V | 841, 823 , 805, 751, 733, 721, 703, 673, 661, 631, 613, 601, 589, 559, 527, 477, 437, 419, 401, 365, 359, 329, 313, 299, 271, 259 | ||||
| k | 18.31 | tetrahydroxyxanthone-C-hexoside dimer c | 259, 318, 368 | + | 843.1597 | C38H35O22 | −2.7 | 843 | 843, 825, 808, 789, 771, 753, 729, 705, 687, 669, 651, 639, 627, 603, 585, 573, 555, 472, 425 |
| − | 841.1463 | C38H33O22 | 0.0 | 841, CE = 45 V | 841, 823, 805, 751, 733, 721, 703, 691, 673, 661, 631, 613, 601, 589, 559, 437, 419, 407, 373, 359, 329, 313, 299, 271, 259 | ||||
| l | 19.02 | tetrahydroxyxanthone-di-O,C-hexoside | 259, 314, 367 | + | 585.1454 | C25H29O16 | −0.3 | 585 | 405, 387, 369, 357, 351, 339, 327, 313, 303, 299, 285, 273, 261 |
| − | 583.1287 | C25H27O16 | −2.1 | 583 | 583, 565, 493, 463, 421, 403, 331, 313, 301, 271, 259 | ||||
| n | 21.17 | aspalathin derivative of (iso)mangiferin | 261, 319, 372 | + | 873.2104 | C40H41O22 | 1.7 | 873 | 819, 807, 731, 694, 675, 658, 631, 627, 616, 604, 591, 573, 561, 541, 525, 507, 489, 475, 459, 447, 439, 423, 405, 387, 369, 357, 345, 327, 313, 303, 289, 273, 261, 247, 217, 196, 163, 151, 149, 139, 123 |
| − | 871.1948 | C40H39O22 | 1.7 | 871 | 871, 751, 691, 601, 571, 557, 539, 449, 437, 421, 331, 301, 269, 243 | ||||
| r | 23.26 | nothofagin derivative of (iso)mangiferin | 261, 319, 372 | + | 857.2137 | C40H41O21 | −0.3 | 857 | 677, 659, 641, 623, 599, 575, 557, 541, 523, 509, 487, 475, 463, 447, 439, 423, 405, 387, 369, 357, 327, 303, 285, 273, 257, 245, 231, 151, 139, 119, 107 |
| − | 855.1984 | C40H39O21 | 0.8 | 855 | 855, 837, 765, 735, 675, 657, 585, 555, 421, 403, 331, 313, 301 | ||||
| 4 | 25.23 | mangiferin (2-C-β-d-glucopyranosyl-1,3,6,7-tetrahydroxyxanthone) | 258, 318, 366 | + | 423.0931 | C19H19O11 | 0.9 | 423 | 369, 351, 339, 327, 313, 303, 299, 285, 273, 257 |
| − | 421.0770 | C19H17O11 | −0.2 | 421 | 331, 313, 301, 285, 271, 259 | ||||
| 5 | 25.86 | isomangiferin (4-C-β-d-glucopyranosyl-1,3,6,7-tetrahydroxyxanthone) | 255, 316, 365 | + | 423.0929 | C19H19O11 | 0.5 | 423 | 387, 369, 357, 341, 327, 313, 303, 285, 273 |
| − | 421.0762 | C19H17O11 | −2.1 | 421 | 331, 313, 301, 285, 271, 259 | ||||
| y | 37.41 | tetrahydroxyxanthone-C-hexoside isomer | 258, 317, 366 | + | 423.0930 | C19H19O11 | 0.7 | 423 | 369, 351, 339, 327, 313, 303, 299, 285, 273, 257 |
| − | 421.0777 | C19H17O11 | 1.4 | 421 | 331, 313, 301, 285, 271, 259 | ||||
| aa | 44.24 | tetrahydroxyxanthone-C-hexoside isomer | 258, 318, 366 | + | 423.0948 | C19H19O11 | 5.0 | 423 | 351, 339, 327, 313, 303, 299, 285, 273, 257 |
| − | 421.0771 | C19H17O11 | 0.0 | 421 | 331, 313, 301, 285, 271, 259 | ||||
| cc | 50.68 | schoepfin A derivative of (iso)mangiferin | 261, 317, 368 | + | 841.2220 | C40H41O20 | 3.4 | 841 | 661, 643, 621, 583, 559, 541, 523, 509, 491, 475, 463, 439, 431, 423, 405, 387, 357, 351, 327, 303, 273, 257, 231 |
| − | 839.2026 | C40H39O20 | −1.1 | 839 | 839, 821, 749, 719, 677, 551, 539, 527, 461, 449, 431, 421, 403, 331, 301, 271 |

| Nr | tR (min) | Proposed Compound | λmax (nm) | Mode | Accurate Mass, exp. | Proposed Formula | Error (ppm) | Precursor Ion | LC-MS/MS Ions a,b |
|---|---|---|---|---|---|---|---|---|---|
| Amino Acids | |||||||||
| 1 | 2.83 | tyrosine | 225, 233, 274 (weak) | + | 182.0818 | C9H12NO3 | 0.5 | 182 | 182, 148, 136, 119, 107, 95, 91, 77 |
| 2 | 4.89 | phenylalanine | 225, 233, 259 (weak) | + | 166.0871 | C9H12NO2 | 1.8 | 166 | 120, 103, 91, 77 |
| Glycosylated phenolic acids | |||||||||
| e | 11.14 | dihydroxybenzoic acid-O-pentoside | 247, 289 sh, 314 | − | 285.0607 | C12H13O8 | −1.1 | 285 | 153, 152, 109, 108 |
| h | 16.57 | dihydroxybenzoic acid-O-dipentoside | 290 (weak) | − | 417.1032 | C17H21O12 | −0.2 | 417 | 417, 285, 241, 153, 152, 109, 108 |
| j | 16.77 | phenyllactic acid 2-O-hexoside | nd | − | 327.1080 | C15H19O8 | 0.0 | 327 | 165, 147, 103 |
| m | 19.18 | coumaric acid-O-(pentosyl)hexoside | nd c | − | 457.1336 | C20H25O12 | −2.2 | 457 | 457, 325, 163, 119 |
| o | 21.52 | caffeic acid-O-(pentosyl)hexoside | 225, 277 | − | 473.1281 | C20H25O13 | −3.0 | 473 | 473, 341, 323, 179, 135 |
| Flavones | |||||||||
| 6 | 27.60 | apigenin-6,8-di-C-glucoside (vicenin-2) | 270, 330 | + | 595.1652 | C27H31O15 | −1.8 | 595 | 505, 457, 439, 427, 421, 409, 403, 391, 379, 355, 349, 337, 325, 307, 295 |
| − | 593.1514 | C27H29O15 | 1.3 | 593 | 593, 575, 503, 485, 473, 455, 413, 395, 383, 365, 353 | ||||
| 10 | 48.10 | diosmetin-7-O-rutinoside (diosmin) | 260, 319 (weak) | + | 609.1819 | C28H33O15 | 0.0 | 609 | 301 |
| − | 607.1673 | C28H31O15 | 1.6 | 607 | 299, 284 | ||||
| Dihydrochalcones | |||||||||
| x | 33.54 | 3-hydroxyphloretin-3',5'-di-C-hexoside | 285 | + | 615.1926 | C27H35O16 | 0.2 | 615 | 513, 495, 477, 465, 447, 435, 423, 411, 399, 381, 369, 357, 345, 327, 313, 301, 259, 247, 235, 217, 205, 193, 165, 123 |
| − | 613.1771 | C27H33O16 | 0.3 | 613 | 505, 493, 475, 433, 415, 403, 385, 373, 361, 331, 251, 239, 209 | ||||
| z | 39.26 | phloretin-3',5'-di-C-glucoside | 286 | + | 599.1971 | C27H35O15 | −0.8 | 599 | 497, 479, 461, 449, 431, 419, 413, 407, 395, 383, 377, 365, 353, 341, 329, 301, 259, 247, 235, 107 |
| − | 597.1830 | C27H33O15 | 1.8 | 597 | 489, 477, 459, 429, 417, 399, 387, 369, 357, 345, 327, 315 | ||||
2.2.3. Flavanones
| Nr | tR (min) | Proposed Compound | λmax (nm) a | Mode | Accurate Mass, exp. | Proposed Formula | Error (ppm) | Precursor Ion | LC-MS/MS Ions b,c |
|---|---|---|---|---|---|---|---|---|---|
| p | 22.44 | eriodictyol-O-hexose-O-pentose | 225, 282 | + | 583.1705 | C26H31O15 | 7.2 | 583 | 356, 289, 219, 195, 154 |
| − | 581.1508 | C26H29O15 | 0.3 | 581 | 581, 445, 419, 401, 313, 299, 287, 161, 151, 135, 125 | ||||
| q1 | 22.90 | eriodictyol-O-hexose-O-pentose | 225, 282 d | + | 583.1436 | nd e | 583 | 289, 261, 195, 163 | |
| − | 581.1519 | C26H29O15 | 2.2 | 581 | 581, 445, 419, 401, 299, 287, 161, 151, 135, 125 | ||||
| q2 | 22.90 | eriodictyol-O-hexose-O-deoxyhexose | 225, 282 d | + | 597.1788 | C27H33O15 | −5.2 | 597 | 473, 313, 289, 195, 163 |
| − | 595.1656 | C27H31O15 | −1.2 | 595 | 595, 459, 433, 313, 287, 169, 161, 151, 135, 125 | ||||
| s | 24.38 | eriodictyol-O-hexose-O-deoxyhexose | 225, 282 | + | 597.1826 | C27H33O15 | 1.2 | 597 | 355, 289, 219, 195, 163 |
| − | 595.1666 | C27H31O15 | 0.5 | 595 | 595, 459, 433, 313, 287, 169, 161, 151, 135, 125 | ||||
| t | 28.54 | naringenin derivative | 278 (weak) | + | 567.1711 | C26H31O14 | −0.5 | 567 | 573, 569, 478, 452, 414, 404, 381, 372, 352, 339, 330, 301, 285, 273, 261, 236, 196, 173 |
| − | 565.1545 | C26H29O14 | −2.1 | 565 | 565, 445, 419, 299, 271, 257, 227, 199, 179, 169, 149, 145, 125, 117 | ||||
| u | 28.95 | naringenin derivative | 280 | + | 567.1668 | C26H31O14 | −8.1 | 567 | 285, 273, 261, 195 |
| − | 565.1558 | C26H29O14 | 0.2 | 565 | 565, 445, 419, 299, 271, 257, 209, 203, 169, 149, 145, 125 | ||||
| v | 29.28 | naringenin-O-hexose-O-deoxyhexose | 281 | + | 581.1891 | C27H33O14 | 3.6 | 581 | 351, 339, 315, 297, 285, 273, 261, 231, 219, 195, 165, 153, 147 |
| − | 579.1722 | C27H31O14 | 1.4 | 579 | 579, 485, 459, 433, 415, 313, 271, 253, 209, 151, 145, 125 | ||||
| w | 30.36 | naringenin-O-hexose-O-deoxyhexose | 280 | + | 581.1866 | C27H33O14 | −0.7 | 581 | 351, 339, 315, 297, 285, 273, 261, 251, 231, 219, 195, 147 |
| − | 579.1722 | C27H31O14 | 1.4 | 579 | 579, 485, 459, 433, 415, 313, 271, 253, 209, 169, 151, 145, 125 | ||||
| 7 | 33.35 | eriodictyol-7-O-rutinoside (eriocitrin) | 281 | + | 597.1812 | C27H33O15 | −1.2 | 597 | 289 |
| − | 595.1664 | C27H31O15 | 0 | 595 | 595, 459, 287, 175, 151, 135, 125, 107, 83 | ||||
| 8 | 39.47 | naringenin-7-O-rutinoside (narirutin) | 280 | + | 581.1856 | C27H33O14 | −2.4 | 581 | 339, 315, 289, 285, 273, 263, 245, 219, 195, 163, 153, 147 |
| − | 579.1688 | C27H31O14 | −4.5 | 579 | 313, 295, 271, 151 | ||||
| 9 | 43.58 | hesperetin-7-O-rutinoside (hesperidin) | 283 | + | 611.1976 | C28H35O15 | 0.0 | 611 | 303 |
| − | 609.1834 | C28H33O15 | 2.5 | 609 | 301 | ||||
| bb | 44.43 | naringenin-O-deoxyhexose(1→2) hexose | nd d | + | 581.1869 | C27H33O14 | −0.2 | 581 | 315, 273, 231, 219, 195, 153, 147 |
| − | 579.1691 | C27H31O14 | −4.0 | 579 | 579, 485, 459, 433, 415, 313, 271, 253, 209, 177, 151, 145, 125 |
2.2.4. Amino Acids
2.2.5. Glycosylated Phenolic Acids
2.2.6. Flavones
2.2.7. Dihydrochalcones
2.3. HPLC-DAD Method Validation
| Phenolic Standard | Number of Calibration Points, n | Wavelength, nm | Linearity Range, µg on-column | Regression Equation a | Correlation Coefficient, r2 |
|---|---|---|---|---|---|
| Maclurin | 6 | 320 | 0.0202–0.2527 | y = 2034.4 x + 1.3312 | 0.9999 |
| Mangiferin b | 7 | 320 | 0.0364–3.6422 | y = 2114.9 x + 1.9834 | 0.9999 |
| Vicenin-2 b | 8 | 320 | 0.0060–0.9008 | y = 1647.0 x – 2.3668 | 0.9995 |
| Aspalathin | 6 | 288 | 0.0301–0.3760 | y = 2305.6 x + 3.9468 | 0.9999 |
| Eriocitrin | 6 | 288 | 0.0200–0.2500 | y = 1607.5 x + 1.7281 | 0.9999 |
| Narirutin | 7 | 288 | 0.0200–0.3340 | y = 1584.9 x + 1.7940 | 0.9999 |
| Hesperidin | 6 | 288 | 0.0776–0.9700 | y = 1631.5 x + 2.1973 | 0.9999 |
2.4. Quantification of Phenolic Compounds
| Nr | Compound | Unfermented Extract | Fermented Extract |
|---|---|---|---|
| a | Maclurin-di-O,C-hexoside a | 0.096 | 0.117 |
| b | Iriflophenone-di-O,C-hexoside b | 6.101 | 5.540 |
| c | Maclurin-3-C-glucoside a | 0.173 | 0.041 |
| A | Unidentified compound c | 0.159 | 0.105 |
| 3 | Iriflophenone-3-C-glucoside | 1.222 | 0.498 |
| k | Tetrahydroxyxanthone-C-hexoside dimer d | nq e | 0.074 |
| l | Tetrahydroxyxanthone di-O,C-hexoside d | 0.190 | 0.080 |
| s | Eriodictyol-O-hexose-O-deoxyhexose f | 0.143 | 0.100 |
| 4 | Mangiferin | 13.791 | 6.966 |
| 5 | Isomangiferin | 1.617 | 0.907 |
| 6 | Vicenin-2 | 0.493 | 0.420 |
| v | Naringenin-O-hexose-O-deoxyhexose g | 0.147 | 0.206 |
| w | Naringenin-O-hexose-O-deoxyhexose g | 0.441 | 0.219 |
| 7 | Eriocitrin | 0.045 | 0.041 |
| x | 3-Hydroxyphloretin-3',5'-di-C-hexoside h | 0.125 | 0.029 |
| y | Tetrahydroxyxanthone-C-hexoside isomer d | 0.109 | 0.053 |
| z | Phloretin-3',5'-di-C-glucoside i | 0.273 | 0.145 |
| 9 | Hesperidin | 0.374 | 0.268 |
3. Experimental Section
3.1. Chemicals
3.2. Sample Preparation
3.3. HPLC-DAD Method Development
3.4. Optimized HPLC-DAD Method
3.5. LC-DAD-ESI-MS and -MS/MSAnalyses
3.6. HPLC-DAD Method Validation
3.7. Quantification of Phenolic Compounds in Freeze-Dried Aqueous Extracts of C. genistoides
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Joubert, E.; Joubert, M.E.; Bester, C.; de Beer, D.; de Lange, J.H. Honeybush (Cyclopia spp.): From local cottage industry to global markets — The catalytic and supporting role of research. S. Afr. J. Bot. 2011, 77, 887–907. [Google Scholar]
- Greenish, H. Cape tea. Pharm. J. 1881, 11, 549–551. [Google Scholar]
- Joubert, E.; Otto, F.; Grüner, S.; Weinreich, B. Reversed-phase HPLC determination of mangiferin, isomangiferin and hesperidin in Cyclopia and the effect of harvesting date on the phenolic composition of C. genistoides. Eur. Food Res. Technol. 2003, 216, 270–273. [Google Scholar]
- Joubert, E.; Manley, M.; Botha, M. Evaluation of spectrophotometric methods for screening of green rooibos (Aspalathus linearis) and green honeybush (Cyclopia genistoides) extracts for high levels of bio-active compounds. Phytochem. Anal. 2008, 19, 169–178. [Google Scholar]
- Joubert, E.; de Beer, D.; Hernández, I.; Munné-Bosch, S. Accummulation of mangiferin, isomangiferin, iriflophenone-3-C-β-glucoside and hesperidin in honeybush leaves (Cyclopia genistoides Vent.) in response to harvest time, harvest interval and seed source. Ind. Crops Prod. 2014, 56, 74–82. [Google Scholar] [CrossRef]
- Vyas, A.; Syeda, K.; Ahmad, A.; Padhye, S.; Sarkar, F.H. Perspectives on medicinal properties of mangiferin. Mini-Rev. Med. Chem. 2012, 12, 412–425. [Google Scholar]
- Kokotkiewicz, A.; Luczkiewicz, M.; Pawlowska, J.; Luczkiewicz, P.; Sowinski, P.; Witkowski, J.; Bryl, E.; Bucinski, A. Isolation of xanthone and benzophenone derivatives from Cyclopia genistoides (L.) Vent. (honeybush) and their pro-apoptotic activity on synoviocytes from patients with rheumatoid arthritis. Fitoterapia 2013, 90, 199–208. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, L.; Ge, D.; Liu, X.; Liu, E.; Wu, C.; Gao, X.; Wang, T. Isolation, structural elucidation, MS profiling, and evaluation of triglyceride accumulation inhibitory effects of benzophenone C-glucosides from leaves of Mangifera indica L. J. Agric. Food Chem. 2013, 61, 1884–1895. [Google Scholar] [CrossRef]
- Verhoog, N.; Joubert, E.; Louw, A. Screening of four Cyclopia (honeybush) species for putative phyto-oestrogenic activity by oestrogen receptor binding assays. S. Afr. J. Sci. 2007, 103, 13–21. [Google Scholar]
- Joubert, E.; Richards, E.S.; van der Merwe, J.D.; de Beer, D.; Manley, M.; Gelderblom, W.C.A. Effect of species variation and processing on phenolic composition and in vitro antioxidant activity of aqueous extracts of Cyclopia spp. (honeybush tea). J. Agric. Food Chem. 2008, 56, 954–963. [Google Scholar] [CrossRef]
- De Beer, D.; Joubert, E. Development of HPLC method for Cyclopia subternata phenolic compound analysis and application to other Cyclopia spp. J. Food Comp. Anal. 2010, 23, 289–297. [Google Scholar] [CrossRef]
- De Beer, D.; Schulze, A.E.; Joubert, E.; de Villiers, A.; Malherbe, C.J.; Stander, M.A. Food ingredient extracts of Cyclopia subternata (honeybush): Variation in phenolic composition and antioxidant capacity. Molecules 2012, 17, 14602–14624. [Google Scholar]
- Schulze, A.E. HPLC Method Development for Characterisation of the Phenolic Composition of Cyclopia subternata and C. maculata Extracts and Chromatographic Fingerprint Analysis for Quality Control. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, December 2013. [Google Scholar]
- De Villiers, A.; Lestremau, F.; Szucs, R.; Gélébart, S.; David, F.; Sandra, P. Evaluation of ultra performance liquid chromatography: Part I. Possibilities and limitations. J. Chromatogr. A 2006, 1127, 60–69. [Google Scholar] [CrossRef]
- Kalili, K.M.; Cabooter, D.; Desmet, G.; de Villiers, A. Kinetic optimisation of the reversed phase liquid chromatographic separation of proanthocyanidins on sub-2 μm and superficially porous phases. J. Chromatogr. A 2012, 1236, 63–76. [Google Scholar] [CrossRef]
- Dolan, J.W. Temperature selectivity in reversed-phase high performance liquid chromatography. J. Chromatogr. A 2002, 965, 195–205. [Google Scholar] [CrossRef]
- Abad-García, B.; Berrueta, L.A.; Garmón-Lobato, S.; Gallo, B.; Vicente, F. A general analytical strategy for the characterization of phenolic compounds in fruit juices by high-performance liquid chromatography with diode array detection coupled to electrospray ionization and triple quadrupole mass spectrometry. J. Chromatogr. A 2009, 1216, 5398–5415. [Google Scholar] [CrossRef]
- Cuyckens, F.; Claeys, M. Mass spectrometry in the structural analysis of flavonoids. J. Mass Spectrom. 2004, 39, 461–461. [Google Scholar] [CrossRef]
- Ito, T.; Kakino, M.; Tazawa, S.; Oyama, M.; Maruyama, H.; Araki, Y.; Hara, H.; Iinuma, M. Identification of phenolic compounds in Aquilaria crassna leaves via liquid chromatography-electrospray ionization mass spectroscopy. Food Sci. Technol. Res. 2012, 18, 259–262. [Google Scholar] [CrossRef]
- Feng, J.; Yang, X.W.; Wang, R.F. Bio-assay guided isolation and identification of α-glucosidase inhibitors from the leaves of Aquilaria sinensis. Phytochemistry 2011, 72, 242–247. [Google Scholar] [CrossRef]
- Malherbe, C.J.; Willenburg, E.; de Beer, D.; Bonnet, S.L.; van der Westhuizen, J.H.; Joubert, E. Iriflophenone-3-C-glucoside from Cyclopia genistoides: Isolation and quantitative comparison of antioxidant capacity with mangiferin and isomangiferin using on-line HPLC antioxidant assays. J. Chromatogr. B 2014, 951–952C, 164–171. [Google Scholar]
- Du, X.G.; Wang, W.; Zhang, Q.Y.; Cheng, J.; Avula, B.; Khan, I.A.; Guo, D.A. Identification of xanthones from Swertia punicea using high-performance liquid chromatography coupled with electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 2913–2923. [Google Scholar] [CrossRef]
- Ichiki, H.; Miura, T.; Kubo, M.; Ishihara, E.; Komatsu, Y.; Tanigawa, K.; Okada, M. New antidiabetic compounds, mangiferin and its glucoside. Biol. Pharm. Bull. 1998, 21, 1389–1390. [Google Scholar]
- Miura, T.; Ichiki, H.; Iwamoto, N.; Kato, M.; Kubo, M.; Sasaki, H.; Okada, M.; Ishida, T.; Seino, Y.; Tanigawa, K. Antidiabetic activity of the rhizoma of Anemarrhena asphodeloides and active components, mangiferin and its glucoside. Biol. Pharm. Bull. 2001, 24, 1009–1011. [Google Scholar]
- Muruganandan, S.; Srinivasan, K.; Gupta, S.; Gupta, P.K.; Lal, J. Effect of mangiferin on hyperglycemia and atherogenicity in streptozotocin diabetic rats. J. Ethnopharmacol. 2005, 97, 497–501. [Google Scholar] [CrossRef]
- Abdel-Mageed, W.M.; Bayoumi, S.A.H.; Chen, C.; Vavricka, C.J.; Li, L.; Malik, A.; Dai, H.; Song, F.; Wang, L.; Zhang, J.; et al. Benzophenone C-glucosides and gallotannins from mango tree stem bark with broad-spectrum anti-viral activity. Bioorg. Med. Chem. 2014, 22, 2236–2243. [Google Scholar] [CrossRef]
- Huang, C.F.; Gan, X.W.; Bai, H.Y.; Ma, L.; Hu, L.H. Schoepfin A, B, C: Three new chalcone C-glycosides from Schoepfia chinensis. Nat. Prod. Res. 2008, 22, 623–627. [Google Scholar]
- Wang, J.N.; Hou, C.Y.; Liu, Y.L.; Lin, L.Z.; Gil, R.R.; Cordell, G.A. Swertifrancheside, an HIV-Reverse transcriptase inhibitor and the first flavone-xanthone dimer, from Swertia franchetiana. J. Nat. Prod. 1994, 57, 211–217. [Google Scholar] [CrossRef]
- Andersen, Ø.M.; Markham, K.R. Flavonoids: Chemistry, Biochemistry and Applications; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Djoukeng, J.D.; Arbona, V.; Argamasilla, R.; Gomez-Cadenas, A. Flavonoid profiling in leaves of citrus genotypes under different environmental situations. J. Agric. Food Chem. 2008, 56, 11087–11097. [Google Scholar] [CrossRef]
- Cuyckens, F.; Rozenberg, R.; de Hoffmann, E.; Claeys, M. Structure characterization of flavonoid O-diglycosides by positive and negative nano-electrospray ionization ion trap mass spectrometry. J. Mass Spectrom. 2001, 36, 1203–1210. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconjugate J. 1988, 5, 397–409. [Google Scholar]
- Louw, A.; Joubert, E.; Visser, K. Phytoestrogenic potential of Cyclopia extracts and polyphenols. Planta Med. 2013, 79, 580–590. [Google Scholar] [CrossRef]
- Natella, F.; Nardini, M.; di Felice, M.; Scaccini, C. Benzoic and cinnamic acid derivatives as antioxidants: Structure−activity relation. J. Agric. Food Chem. 1999, 47, 1453–1459. [Google Scholar]
- Bartsch, M.; Bednarek, P.; Vivancos, P.D.; Schneider, B.; von Roepenack-Lahaye, E.; Foyer, C.H.; Kombrink, E.; Scheel, D.; Parker, J.E. Accumulation of isochorismate-derived 2,3-dihydroxybenzoic 3-O-beta-d-xyloside in arabidopsis resistance to pathogens and ageing of leaves. J. Biol. Chem. 2010, 285, 25654–25665. [Google Scholar] [CrossRef]
- Fayos, J.; Bellés, J.M.; López-Gresa, M.P.; Primo, J.; Conejero, V. Induction of gentisic acid 5-O-β-d-xylopyranoside in tomato and cucumber plants infected by different pathogens. Phytochemistry 2006, 67, 142–148. [Google Scholar] [CrossRef]
- Zhang, S.; Xuan, L. New phenolic constituents from the stems of Spatholobus suberectus. Helv. Chim. Acta 2006, 89, 1241–1245. [Google Scholar] [CrossRef]
- Tsopmo, A.; Muir, A.D. Chemical profiling of lentil (Lens culinaris Medik.) cultivars and isolation of compounds. J. Agric. Food Chem. 2010, 58, 8715–8721. [Google Scholar] [CrossRef]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Lamuela-Raventós, R.M.; Bastida, J.; Viladomat, F.; Codina, C. Identification of phenolic compounds in artichoke waste by high-performance liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2003, 1008, 57–72. [Google Scholar] [CrossRef]
- Lu, Y.; Yeap Foo, L. Flavonoid and phenolic glycosides from Salvia officinalis. Phytochemistry 2000, 55, 263–267. [Google Scholar] [CrossRef]
- Meng, D.; Wu, J.; Zhao, W. Glycosides from Breynia fruticosa and Breynia rostrata. Phytochemistry 2010, 71, 325–331. [Google Scholar] [CrossRef]
- Ferreira, D.; Kamara, B.I.; Brandt, E.V.; Joubert, E. Phenolic compounds from Cyclopia intermedia (honeybush tea). 1. J. Agric. Food Chem. 1998, 46, 3406–3410. [Google Scholar] [CrossRef]
- Kamara, B.I.; Brandt, E.V.; Ferreira, D.; Joubert, E. Polyphenols from honeybush tea (Cyclopia intermedia). J. Agric. Food Chem. 2003, 51, 3874–3879. [Google Scholar] [CrossRef]
- Kamara, B.I.; Brand, D.J.; Brandt, E.V.; Joubert, E. Phenolic metabolites from honeybush tea (Cyclopia subternata). J. Agric. Food Chem. 2004, 52, 5391–5395. [Google Scholar] [CrossRef]
- Wang, M.; Shao, Y.; Li, J.; Zhu, N.; Rangarajan, M.; LaVoie, E.J.; Ho, C.T. Antioxidative phenolic glycosides from sage (Salvia officinalis). J. Nat. Prod. 1999, 62, 454–456. [Google Scholar] [CrossRef]
- Vitalini, S.; Braca, A.; Fico, G. Study on secondary metabolite content of Helleborus niger L. leaves. Fitoterapia 2011, 82, 152–154. [Google Scholar] [CrossRef]
- Beelders, T.; Sigge, G.O.; Joubert, E.; de Beer, D.; de Villiers, A. Kinetic optimisation of the reversed phase liquid chromatographic separation of rooibos tea (Aspalathus linearis) phenolics on conventional high performance liquid chromatographic instrumentation. J. Chromatogr. A 2012, 1219, 128–139. [Google Scholar] [CrossRef]
- Beelders, T.; Sigge, G.O.; Joubert, E.; de Beer, D.; de Villiers, A. Erratum to “Kinetic optimisation of the reversed phase liquid chromatographic separation of rooibos tea (Aspalathus linearis) phenolics on conventional high performance liquid chromatographic instrumentation” [J. Chromatogr. A 1219 (2012) 128–139]. J. Chromatogr. A 2012, 1241, 128. [Google Scholar] [CrossRef]
- Kokotkiewicz, A.; Luczkiewicz, M.; Sowinski, P.; Glod, D.; Gorynski, K.; Bucinski, A. Isolation and structure elucidation of phenolic compounds from Cyclopia subternata Vogel (honeybush) intact plant and in vitro cultures. Food Chem. 2012, 133, 1373–1382. [Google Scholar] [CrossRef]
- Roowi, S.; Crozier, A. Flavonoids in tropical citrus species. J. Agric. Food Chem. 2011, 59, 12217–12225. [Google Scholar] [CrossRef]
- Snyder, L.R.; Kirkland, J.J.; Glajch, J.L. Practical HPLC Method Development, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Beelders, T.; De Beer, D.; Stander, M.A.; Joubert, E. Comprehensive Phenolic Profiling of Cyclopia genistoides (L.) Vent. by LC-DAD-MS and -MS/MS Reveals Novel Xanthone and Benzophenone Constituents. Molecules 2014, 19, 11760-11790. https://doi.org/10.3390/molecules190811760
Beelders T, De Beer D, Stander MA, Joubert E. Comprehensive Phenolic Profiling of Cyclopia genistoides (L.) Vent. by LC-DAD-MS and -MS/MS Reveals Novel Xanthone and Benzophenone Constituents. Molecules. 2014; 19(8):11760-11790. https://doi.org/10.3390/molecules190811760
Chicago/Turabian StyleBeelders, Theresa, Dalene De Beer, Maria A. Stander, and Elizabeth Joubert. 2014. "Comprehensive Phenolic Profiling of Cyclopia genistoides (L.) Vent. by LC-DAD-MS and -MS/MS Reveals Novel Xanthone and Benzophenone Constituents" Molecules 19, no. 8: 11760-11790. https://doi.org/10.3390/molecules190811760