Effects of Drying Methods on Physicochemical and Immunomodulatory Properties of Polysaccharide-Protein Complexes from Litchi Pulp

Abstract
:1. Introduction
2. Results and Discussion
2.1. Preliminary Characterization of Litchi Polysaccharide-Protein Complexes
2.1.1. The Chemical Compositions of Litchi Polysaccharide-Protein Complexes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | LP-VF | LP-VM | LP-HP |
|---|---|---|---|
| Neutral Sugar (%) | 51.51 ± 1.89 c | 38.36 ± 0.84 a | 49.81 ± 2.32 b |
| Uronic Acid (%) | 6.8 ± 0.26 c | 4.23 ± 0.15 a | 4.63 ± 0.35 b |
| Protein (%) | 4.2 ± 0.03 c | 4.03 ± 0.02 a | 4.11 ± 0.04 b |
| Monosaccharides Composition (%) | |||
| Ribose | 2.06 ± 0.35 a | 2.00 ± 0.62 a | 3.43 ± 0.06 b |
| Rhamnose | 0.23 ± 0.04 a | 0.23 ± 0.19 a | 0.38 ± 0.06 a |
| Arabinose | 9.67 ± 1.56 a | 12.79 ± 0.74 b | 27.25 ± 1.09 c |
| Xylose | 0.97 ± 0.28 a | 0.85 ± 0.19 a | 2.04 ± 0.63 c |
| Mannose | 9.69 ± 3.01 a | 12.74 ± 2.98 a | 13.94 ± 0.21 a |
| Glucose | 59.00 ± 4.32 c | 50.75 ± 2.57 b | 11.54 ± 0.8 a |
| Galactose | 17.90 ± 0.18 a | 21.35 ± 2.76 b | 40.08 ± 3.11 c |
| Amino Acids Composition (g/100 g) | |||
| Aspartate | 0.243 ± 0.040 a | 0.257 ± 0.040 a | 0.467 ± 0.095 b |
| Threonine | 0.440 ± 0.030 b | 0.290 ± 0.010 a | 0.378 ± 0.075 b |
| Serine | 0.337 ± 0.042 a | 0.253 ± 0.038 a | 0.450 ± 0.131 b |
| Glutamic | 0.367 ± 0.084 a | 0.363 ± 0.021 a | 0.493 ± 0.040 b |
| Glycine | 0.303 ± 0.032 b | 0.227 ± 0.023 a | 0.283 ± 0.052 b |
| Alanine | 0.423 ± 0.124 a | 0.383 ± 0.462 a | 0.333 ± 0.064 a |
| Valine | 0.487 ± 0.080 b | 0.313 ± 0.015 a | 0.384 ± 0.093 a |
| Methionine | 0.106 ± 0.050 a | 0.062 ± 0.036 a | 0.100 ± 0.020 a |
| Isoleucine | 0.250 ± 0.045 a | 0.200 ± 0.010 a | 0.307 ± 0.068 a |
| Leucine | 0.303 ± 0.045 b | 0.170 ± 0.044 a | 0.273 ± 0.099 ab |
| Tyrosine | 0.087 ± 0.010 a | 0.060 ± 0.014 a | 0.071 ± 0.018 a |
| Phenylalanine | 0.136 ± 0.037 a | 0.097 ± 0.032 a | 0.159 ± 0.072 a |
| Histidine | 0.487 ± 0.076 a | 0.417 ± 0.021 a | 0.460 ± 0.098 a |
| Lysine | 0.393 ± 0.031 a | 0.387 ± 0.072 a | 0.500 ± 0.017 b |
| Arginine | 0.287 ± 0.087 b | 0.075 ± 0.022 a | 0.075 ± 0.005 a |
| Proline | 0.237 ± 0.078 a | 0.24 ± 0.106 a | 0.373 ± 0.071 a |
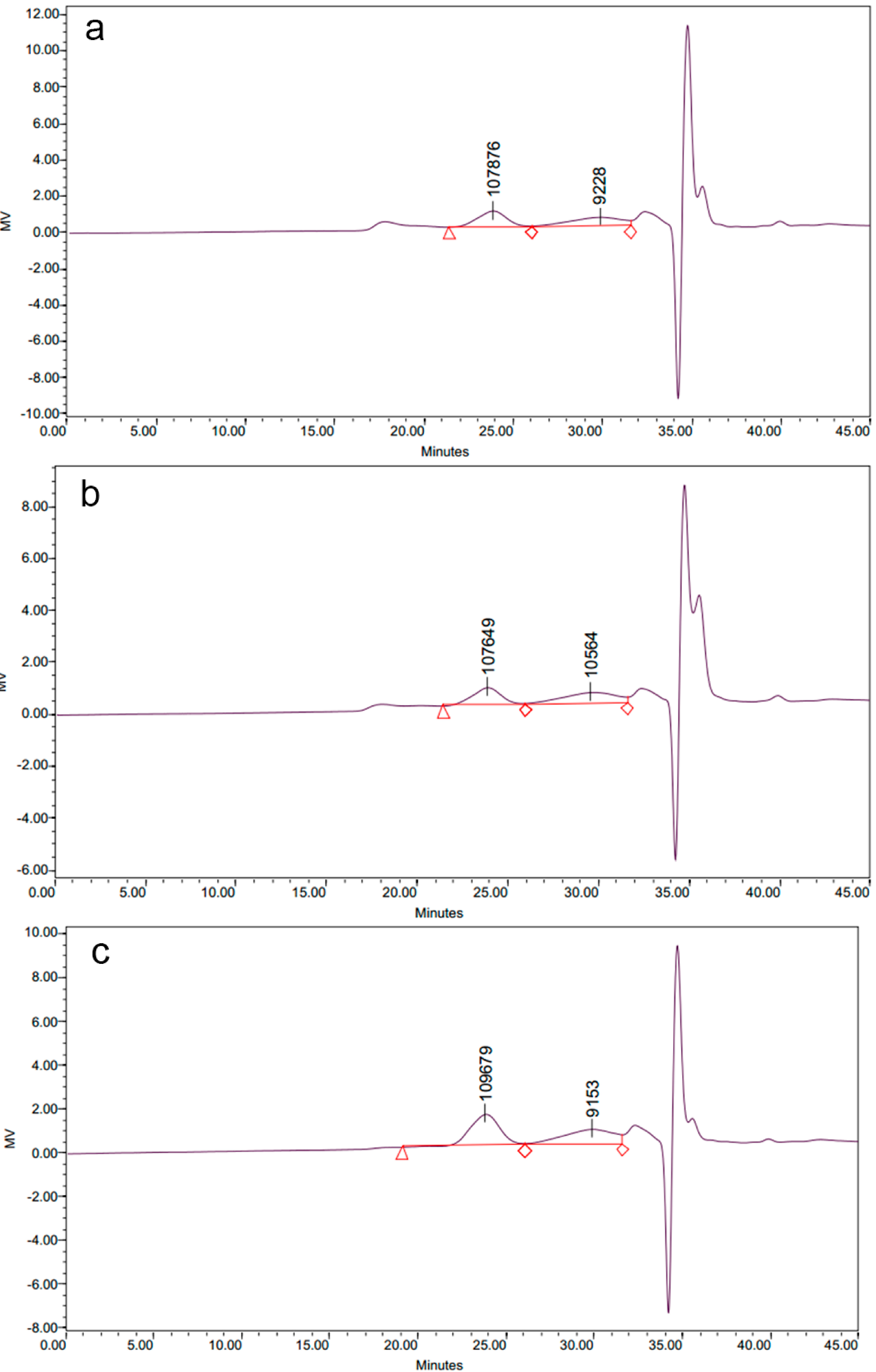
2.1.2. Molecular Weight Distribution of Polysaccharide-Protein Complexes

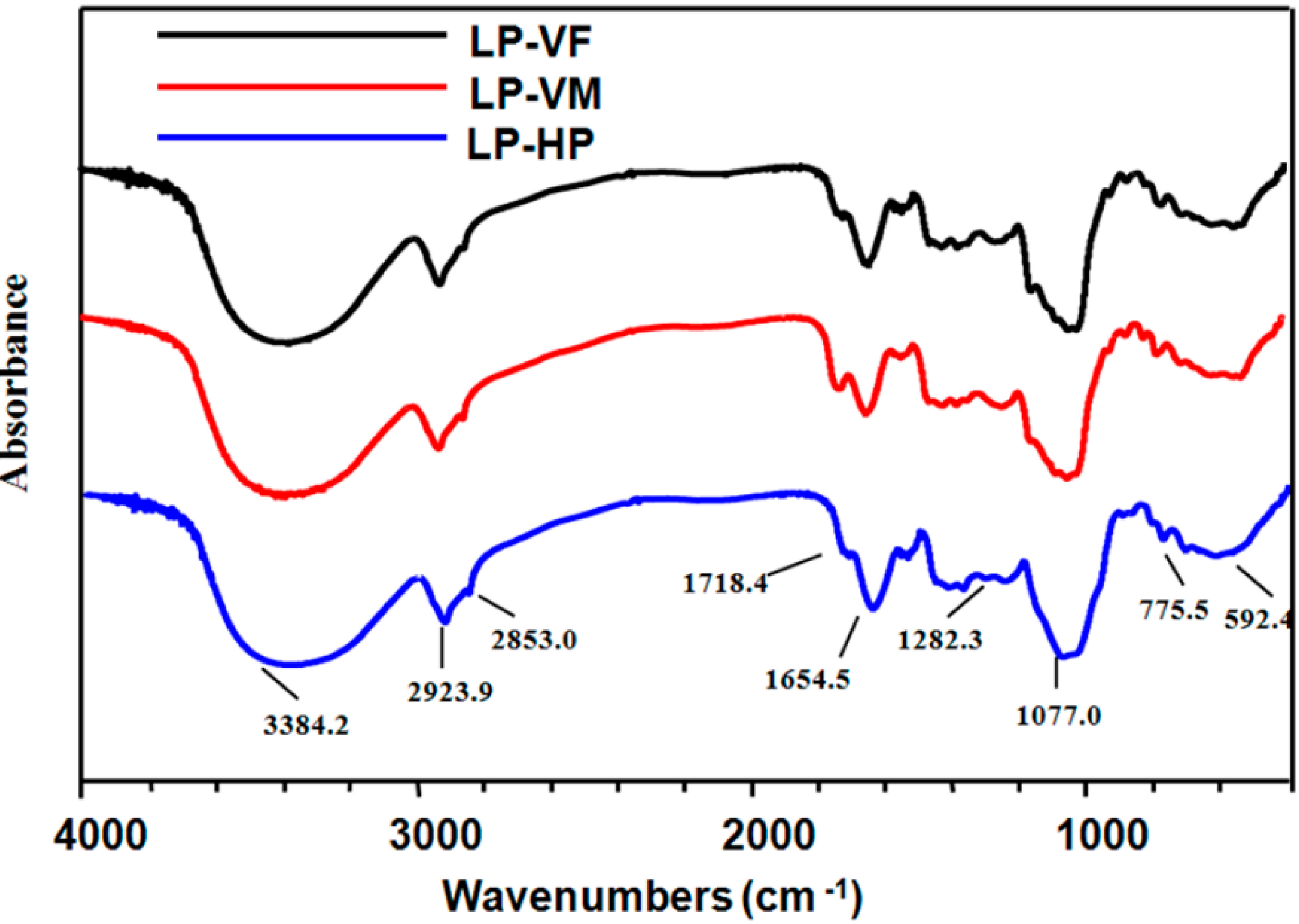
2.1.3. Spectroscopic Characteristics of Polysaccharide-Protein Complexes

2.2. In Vitro Immunostimulatory Activity of Litchi Polysaccharide-Protein Complexes
2.2.1. Effects of Litchi Polysaccharides on Splenocyte Proliferation in Vitro

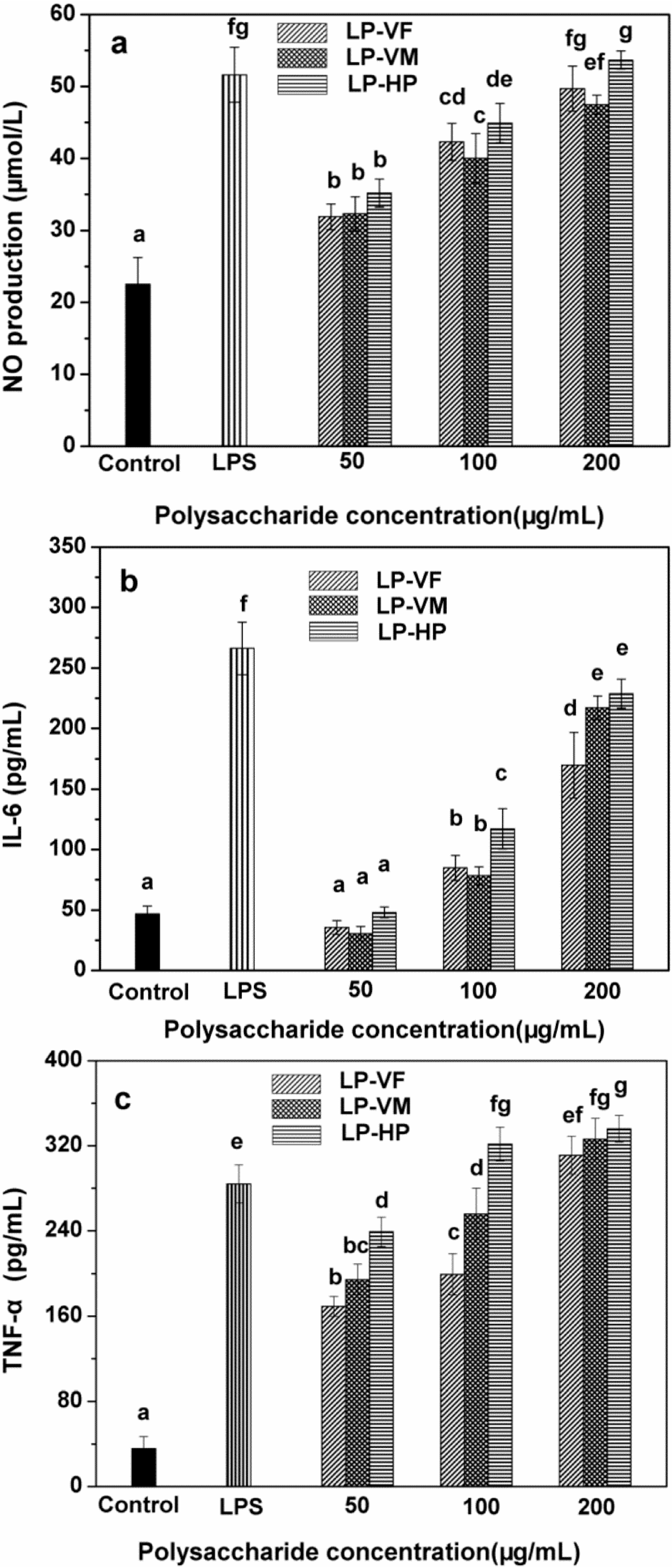
2.2.2. Effects of Litchi Polysaccharide-Protein Complexes on the Production of NO, IL-6 and TNF-α of RAW264.7 Macrophages

2.2.3. Effects of Polymyxin B (PB) on the Biological Activity of Litchi Polysaccharide-Protein Complexes

3. Experimental Section
3.1. Materials and Chemicals
3.1.1. Chemicals and Reagents
3.1.2. Cells and Animals
3.2. Drying Process
3.3. Preparation of Polysaccharide-Protein Complexes
3.4. Preliminary Characterization of Litchi Polysaccharide-Protein Complexes
3.4.1. Analysis of the Chemical Characteristics
3.4.2. Determination of Molecular Weights
3.4.3. Analysis of FT-IR Spectroscopy
3.5. In Vitro Immunostimulatory Activity Assay of Litchi Polysaccharides-Protein Complexes
3.5.1. Determination of Mouse Splenocyte Proliferation

3.5.2. Determination of NO and Cytokine Production of Macrophage
3.5.3. Determination of Endotoxin Contamination of Litchi Polysaccharides-Protein Complexes
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wall, M.M. Ascorbic acid and mineral composition of longan (Dimocarpus longan), lychee (Litchi chinensis) and rambutan (Nephelium lappaceum) cultivars grown in Hawaii. J. Food Compos. Anal. 2006, 19, 655–663. [Google Scholar] [CrossRef]
- Zhang, R.; Zeng, Q.; Deng, Y.; Zhang, M.; Wei, Z.; Zhang, Y.; Tang, X. Phenolic profiles and antioxidant activity of litchi pulp of different cultivars cultivated in Southern China. Food Chem. 2013, 136, 1169–1176. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, B.; Wang, J.; Liu, Y.; Yu, L.; Jiang, Y. Immunomodulatory and anticancer activities of flavonoids extracted from litchi (Litchi chinensis Sonn.) pericarp. Int. Immunopharmacol. 2007, 2, 162–166. [Google Scholar]
- Zhou, Y.; Wang, H.; Yang, R.; Huang, H.; Sun, Y.; Shen, Y.; Lei, H.; Gao, H. Effects of litchi chinensis fruit isolates on prostaglandin E(2) and nitric oxide production in J774 murine macrophage cells. BMC Complement. Altern. Med. 2012, 12. [Google Scholar] [CrossRef]
- Hu, X.Q.; Huang, Y.Y.; Dong, Q.F.; Song, L.Y.; Yuan, F.; Yu, R.M. Structure characterization and antioxidant activity of a novel polysaccharide isolated from pulp tissues of Litchi chinensis. J. Agric. Food Chem. 2011, 21, 11548–11552. [Google Scholar]
- Kong, F.; Zhang, M.; Liao, S.; Yu, S.; Chi, J.; Wei, Z. Antioxidant activity of polysaccharide-enriched fractions extracted from pulp tissue of Litchi chinensis Sonn. Molecules 2010, 4, 2152–2165. [Google Scholar]
- Jing, Y.; Huang, L.; Lv, W.; Tong, H.; Song, L.; Hu, X.; Yu, R. Structure characterization of a novel polysaccharide from pulp tissues of Litchi chinensis and its immunomodulatory activity. J. Agric. Food Chem. 2013, 62, 902–911. [Google Scholar]
- Jiang, Y.; Wang, Y.; Song, L.; Liu, H.; Lichter, A.; Kerdchoechuen, O.; Joyce, D.; Shi, J. Postharvest characteristics and handling of litchi fruit—An overview. Anim. Prod. Sci. 2006, 46, 1541–1556. [Google Scholar]
- Ratti, C. Hot air and freeze-drying of high-value foods: A review. J. Food Eng. 2001, 49, 311–319. [Google Scholar] [CrossRef]
- Mousa, N.; Farid, M. Microwave vacuum drying of banana slices. Dry Technol. 2002, 20, 2055–2066. [Google Scholar] [CrossRef]
- Giri, S.K.; Prasad, S. Drying kinetics and rehydration characteristics of microwave-vacuum and convective hot-air dried mushrooms. J. Food Eng. 2007, 78, 512–521. [Google Scholar] [CrossRef]
- Alves-Filho, O. Combined innovative heat pump drying technologies and new cold extrusion techniques for production of instant foods. Dry. Technol. 2002, 20, 1541–1557. [Google Scholar] [CrossRef]
- Colak, N.; Hepbasli, A. A review of heat-pump drying (HPD): Part 2—Applications and performance assessments. Energ. Convers. Manag. 2009, 50, 2187–2199. [Google Scholar] [CrossRef]
- Femenia, A.; Waldron, K. High-value co-products from plant foods: Cosmetics and pharmaceuticals. In Handbook of Waste Management and Co-product Recovery in Food Processing; CRC Press LLC: Boca Raton, FL, USA, 2007; pp. 484–516. [Google Scholar]
- Ma, L.; Chen, H.; Zhu, W.; Wang, Z. Effect of different drying methods on physicochemical properties and antioxidant activities of polysaccharides extracted from mushroom Inonotus obliquus. Food Res. Int. 2013, 2, 633–640. [Google Scholar]
- Fan, L.; Li, J.; Deng, K.; Ai, L. Effects of drying methods on the antioxidant activities of polysaccharides extracted from Ganoderma lucidum. Carbohyd. Polym. 2012, 87, 1849–1854. [Google Scholar] [CrossRef]
- Huang, F.; Zhang, R.; Yi, Y.; Tang, X.; Zhang, M.; Su, D.; Deng, Y.; Wei, Z. Comparison of physicochemical properties and immunomodulatory activity of polysaccharides from fresh and dried litchi pulp. Molecules 2014, 19, 3909–3925. [Google Scholar] [CrossRef]
- Cui, Z.-W.; Xu, S.-Y.; Sun, D.-W.; Chen, W. Dehydration of concentrated Ganoderma lucidum extraction by combined microwave-vacuum and conventional vacuum drying. Dry. Technol. 2006, 24, 595–599. [Google Scholar] [CrossRef]
- Femenia, A.; Bestard, M.; Sanjuan, N.; Rossello, C.; Mulet, A. Effect of rehydration temperature on the cell wall components of broccoli (Brassica oleracea L. Var. italica) plant tissues. J. Food Eng. 2000, 46, 157–163. [Google Scholar] [CrossRef]
- Tao, Y.; Xu, W. Microwave-assisted solubilization and solution properties of hyperbranched polysaccharide. Carbohydr. Res. 2008, 343, 3071–3078. [Google Scholar] [CrossRef]
- Colin-Henrion, M.; Mehinagic, E.; Renard, C.M.G.C.; Richomme, P.; Jourjon, F. From apple to applesauce: Processing effects on dietary fibres and cell wall polysaccharides. Food Chem. 2009, 117, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Asgar, M.A.; Yamauchi, R.; Kato, K. Modification of pectin in Japanese persimmon fruit during the sun-drying process. Food Chem. 2003, 81, 555–560. [Google Scholar] [CrossRef]
- Mirhosseini, H.; Amid, B.T. Effect of different drying techniques on flowability characteristics and chemical properties of natural carbohydrate-protein Gum from durian fruit seed. Chem. Cent. J. 2013, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Yu, X.; Ma, H.; Liu, S.; Qin, X.; Yagoub, A.E.-G.A.; Owusu, J. Examining of athermal effects in microwave-induced glucose/glycine reaction and degradation of polysaccharide from Porphyra yezoensis. Carbohyd. Polym. 2013, 97, 38–44. [Google Scholar] [CrossRef]
- Zhang, W. Biochemical Technology of Carbohydrate Complexes; Zhejiang University Press: Hangzhou, China, 1994. [Google Scholar]
- Chen, X.; Nie, W.; Yu, G.; Li, Y.; Hu, Y.; Lu, J.; Jin, L. Antitumor and immunomodulatory activity of polysaccharides from Sargassum fusiforme. Food Chem.Toxicol. 2012, 50, 695–700. [Google Scholar] [CrossRef]
- Lo, T.C.; Jiang, Y.H.; Chao, A.L.; Chang, C.A. Use of statistical methods to find the polysaccharide structural characteristics and the relationships between monosaccharide composition ratio and macrophage stimulatory activity of regionally different strains of Lentinula edodes. Anal. Chim. Acta 2007, 584, 50–56. [Google Scholar] [CrossRef]
- Ng, T. A review of research on the protein-bound polysaccharide (polysaccharopeptide, PSP) from the mushroom Coriolus versicolor (basidiomycetes: Polyporaceae). Gen. Pharmacol. 1998, 30, 1–4. [Google Scholar]
- Ren, L.; Perera, C.; Hemar, Y. Antitumor activity of mushroom polysaccharides: A review. Food Funct. 2012, 3, 1118–1130. [Google Scholar] [CrossRef]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Kralovec, J.A.; Metera, K.L.; Kumar, J.R.; Watson, L.V.; Girouard, G.S.; Guan, Y.; Carr, R.I.; Barrow, C.J.; Ewart, H.S. Immunostimulatory principles from Chlorella pyrenoidosa—Part 1: Isolation and biological assessment in vitro. Phytomedicine 2007, 14, 57–64. [Google Scholar]
- Tao, Y.; Zhang, Y.; Zhang, L. Chemical modification and antitumor activities of two polysaccharide—protein complexes from Pleurotus tuber-regium. Int. J. Biol. Macromol. 2009, 45, 109–115. [Google Scholar] [CrossRef]
- Liu, F.; Ooi, V.E.; Liu, W.K.; Chang, S.T. Immunomodulation and antitumor activity of polysaccharide-protein complex from the culture filtrates of a local edible mushroom, Tricholoma lobayense. Gen. Pharmacol. 1996, 27, 621–624. [Google Scholar] [CrossRef]
- Hirano, T. The biology of interleukin-6. Chem. Immunol. 1991, 51, 153–180. [Google Scholar] [CrossRef]
- Kehrl, J.H.; Miller, A.; Fauci, A.S. Effect of tumor necrosis factor α on mitogen-activated human B cells. J. Exp. Med. 1987, 166, 786–791. [Google Scholar] [CrossRef]
- Na, Y.S.; Kim, W.J.; Kim, S.M.; Lee, S.M.; Kim, S.O.; Synytsya, A.; Park, Y. Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens. Int. Immunopharmacol. 2010, 10, 364–370. [Google Scholar] [CrossRef]
- Zhang, C.X.; Dai, Z.R. Immunomodulatory activities on macrophage of a polysaccharide from Sipunculus nudus L. Food Chem. Toxicol. 2011, 49, 2961–2967. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A; Hamilton, J.K.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Mazumder, S.; Morvan, C.; Thakur, S.; Ray, B. Cell wall polysaccharides from chalkumra (Benincasa hispida) fruit: Part I. Isolation and characterization of pectins. J. Agric. Food. Chem. 2004, 52, 3556–3562. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, M.W.; Liao, S.T.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C.; Tang, X.J.; Zhang, Y. Structural features and immunomodulatory activities of polysaccharides of longan pulp. Carbohyd. Polym. 2012, 1, 636–643. [Google Scholar]
- Kim, G.Y.; Lee, J.Y.; Lee, J.O.; Ryu, C.-H.; Choi, B.T.; Jeong, Y.K.; Lee, K.W.; Jeong, S.C.; Choi, Y.H. Partial characterization and immunostimulatory effect of a novel polysaccharide—protein complex extracted from Phellinus linteus. Biosci. Biotech. Bioch. 2006, 70, 1218–1226. [Google Scholar]
- Sample Availability: Samples of polysaccharides are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, F.; Guo, Y.; Zhang, R.; Yi, Y.; Deng, Y.; Su, D.; Zhang, M. Effects of Drying Methods on Physicochemical and Immunomodulatory Properties of Polysaccharide-Protein Complexes from Litchi Pulp. Molecules 2014, 19, 12760-12776. https://doi.org/10.3390/molecules190812760
Huang F, Guo Y, Zhang R, Yi Y, Deng Y, Su D, Zhang M. Effects of Drying Methods on Physicochemical and Immunomodulatory Properties of Polysaccharide-Protein Complexes from Litchi Pulp. Molecules. 2014; 19(8):12760-12776. https://doi.org/10.3390/molecules190812760
Chicago/Turabian StyleHuang, Fei, Yajuan Guo, Ruifen Zhang, Yang Yi, Yuanyuan Deng, Dongxiao Su, and Mingwei Zhang. 2014. "Effects of Drying Methods on Physicochemical and Immunomodulatory Properties of Polysaccharide-Protein Complexes from Litchi Pulp" Molecules 19, no. 8: 12760-12776. https://doi.org/10.3390/molecules190812760