Immunomodulatory Activity and Partial Characterisation of Polysaccharides from Momordica charantia

Abstract
:1. Introduction
2. Results and Discussion
2.1. Immunomodulatory Activity of MCP in Vivo
2.1.1. Effects of MCP on Spleen and Thymus Indexes

2.1.2. Effect of MCP on Macrophage Phagocytosis

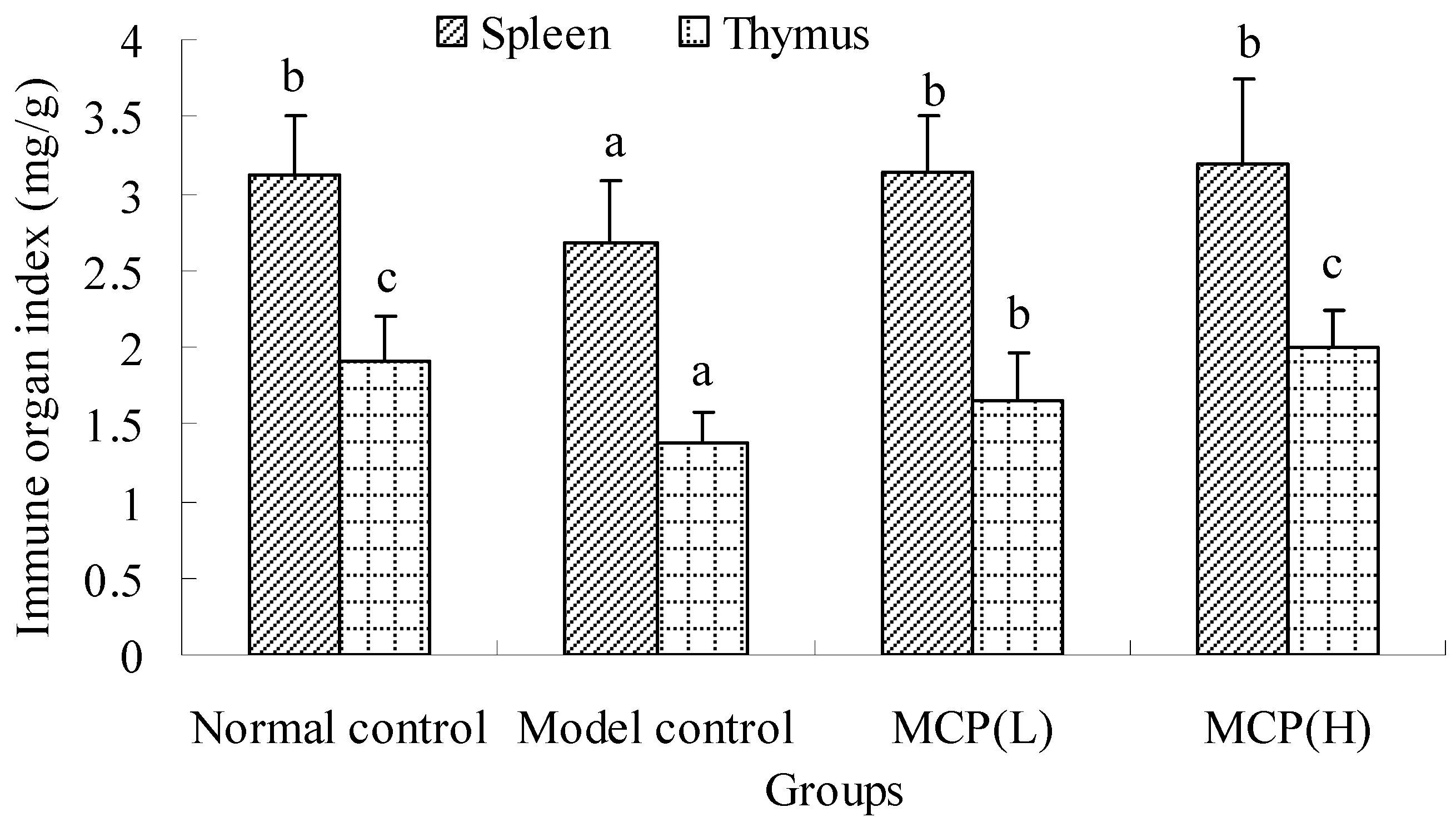{kind=link}
{kind=link}
| Group | Dose (mg·kg−1·d−1) | α | HC50 | PI | NK Activity (%) |
|---|---|---|---|---|---|
| Normal control | — | 5.53 ± 0.95 b | 144.33 ± 11.07 b | 1.001 ± 0.107 b | 43.757 ± 5.160 b |
| Model control | — | 4.22 ± 0.64 a | 123.97 ± 9.53 a | 0.659 ± 0.127 a | 35.382 ± 9.152 a |
| MCP(L) | 150 | 4.97 ± 0.67 b | 136.04 ± 9.02 b | 1.087 ± 0.274 b | 43.478 ± 8.325 b |
| MCP(H) | 300 | 5.04 ± 0.42 b | 158.98 ± 8.93 b | 1.396 ± 0.204 c | 46.397 ± 7.937 b |
2.1.3. Effect of MCP on Haemolysin Production
2.1.4. Effect of MCP on Splenic Lymphocyte Proliferation
2.1.5. Effect of MCP on NK Cell Cytotoxicity
2.2. Effects of MCP1 and MCP2 on Splenic Lymphocyte Proliferation in Vitro
| Dose (µg/mL) | MCP1 | MCP2 | MCP1+ConA | MCP2+ConA |
|---|---|---|---|---|
| 0 | 1.00 ± 0.16 a | 1.00 ± 0.16 a | 1.67 ± 0.34 a∆ | 1.67 ± 0.34 a# |
| 20 | 2.00 ± 0.69 bc | 2.18 ± 0.47 b | 3.29 ± 0.38 b∆ | 2.86 ± 0.36 b |
| 60 | 1.73 ± 0.31 ab | 2.18 ± 0.47 b | 3.36 ± 0.39 bc∆ | 2.32 ± 0.55 b |
| 100 | 3.18 ± 0.42 de | 2.11 ± 0.34 b* | 4.08 ± 0.38 c∆ | 2.32 ± 0.27 b |
| 140 | 2.82 ± 0.42 cd | 2.27 ± 0.63 b | 2.82 ± 0.08 b | 2.77 ± 0.31 b |
| 180 | 3.82 ± 0.72 e | 2.82 ± 0.57 b | 2.77 ± 0.57 b | 3.64 ± 0.28 c |
2.3. Isolation and Purification of MCP

2.4. Characterisation of MCP1 and MCP2
| Composition | MCP1 | MCP2 | ||
|---|---|---|---|---|
| Retention Time (min) | Molar Ratio | Retention Time (min) | Molar Ratio | |
| Ribose | 5.97 | 1.00 | 5.95 | 1.86 |
| Rhamnose | 6.05 | 6.33 | 6.03 | 1.00 |
| Arabinose | 6.11 | 9.07 | 6.14 | 8.92 |
| Xylose | 6.25 | 3.78 | 6.23 | 9.62 |
| Mannose | 8.05 | 4.71 | 8.05 | 34.18 |
| Glucose | 8.12 | 27.28 | 8.12 | 44.20 |
| Galactose | 8.55 | 19.58 | 8.33 | 23.61 |
| Absorption (cm−1) | Functional Group | Structural Characteristic | |
|---|---|---|---|
| MCP1 | MCP2 | ||
| 3431.7 | 3431.2 | Hydroxyl group (–OH) | O–H stretching vibration |
| 2931.9 | 2925.8 | Alkyl group (–CH2-) | C–H stretching vibration |
| 1618 | 1618.5 | Carboxyl group (–C=O or –CHO) | C=O stretching vibration |
| 1419.8 | 1412.7 | Carboxyl group (–COOH) | C–O stretching vibration |
| 1331.5 | Carboxyl group (–COOH) | C=O symmetrical stretching vibration | |
| 1240.7 | Sulphate group (–O–SO3–) | S=O stretching vibration of | |
| 1145.4, 1099.6 | Ether (–C–O–C–) | C–O–C covalent vibration | |
| 1018.8 | 1025.8 | Hydroxyl group (–OH) | O–H bending vibration |
| 954.9 | 955.5 | Pyranose ring | Rolling vibration at the end of the methine |
| 894.2 | 894.7 | β-d-Glucopyranose | |
| 835.2 | 835.3 | α-d-Glucopyranose | |
3. Experimental Section
3.1. Materials and Chemicals
3.2. Preparation of M. charantia Polysaccharides
3.3. Immunomodulatory Activity Analysis
3.3.1. Animals and Cells
| Groups | Normal Saline (mL·kg−·d−1) | Cyclophosphamide (mg·kg−1·d−1) | Normal Saline (mL·kg−1·d−1) | MCP (mg·kg−1·d−1) |
|---|---|---|---|---|
| Intraperitoneal Injection (Days 1–3) | Peroral Administration (Days 4–33) | |||
| Normal control | 5 | – | 5 | – |
| Model control | – | 80 | 5 | – |
| MCP(L) | – | 80 | – | 150 |
| MCP(H) | – | 80 | – | 300 |
3.3.2. Macrophage Phagocytosis Assay
3.3.3. Measurement of Serum Haemolysin
3.3.4. Measurement of Spleen and Thymus Indexes
3.3.5. Splenic Lymphocyte Proliferation Assay
3.3.6. Cytotoxicity Assay of NK Cells
3.4. Characterisation of Polysaccharides
3.4.1. Characterisation and Identification of Monosaccharides
3.4.2. Molecular Weight Measurement
3.4.3. Fourier Transform Infrared Spectrum
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paul, A.; Raychaudhuri, S.S. Medicinal uses and molecular identification of two Momordica charantia varieties—A review. Electron. J. Biol. 2010, 6, 43–51. [Google Scholar]
- Ahmed, I.; Adeghate, E.; Cummings, E.; Sharma, A.; Singh, J. Beneficial effects and mechanism of action of Momordica charantia juice in the treatment of streptozotocin-induced diabetes mellitus in rat. Mol. Cell. Biochem. 2004, 261, 63–70. [Google Scholar] [CrossRef]
- Popovich, D.G.; Li, L.; Zhang, W. Bitter melon (Momordica charantia) triterpenoid extract reduces preadipocyte viability, lipid accumulation and adiponectin expression in 3T3-L1 cells. Food Chem. Toxicol. 2010, 48, 1619–1626. [Google Scholar]
- Umukoro, S.; Ashorobi, R. Evaluation of anti-inflammatory and membrane stabilizing property of aqueous leaf extract of Momordica charantia in rats. Afr. J. Biomed. Res. 2006, 9, 119–124. [Google Scholar]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 10, 881–890. [Google Scholar] [CrossRef]
- Akihisa, T.; Higo, N.; Tokuda, H.; Ukiya, M.; Akazawa, H.; Tochigi, Y.; Kimura, Y.; Suzuki, T.; Nishino, H. Cucurbitane-type triterpenoids from the fruits of Momordica charantia and their cancer chemopreventive effects. J. Nat. Prod. 2007, 70, 1233–1239. [Google Scholar] [CrossRef]
- Lee-Huang, S.; Huang, P.L.; Nara, P.L.; Chen, H.-C.; Kung, H.-F.; Huang, P.; Huang, H.I.; Huang, P.L. MAP 30: A new inhibitor of HIV-1 infection and replication. FEBS Lett. 1990, 272, 12–18. [Google Scholar] [CrossRef]
- Kobori, M.; Ohnishi-Kameyama, M.; Akimoto, Y.; Yukizaki, C.; Yoshida, M. Alpha eleostearic acid and its dihydroxy derivative are major apoptosis inducing components of bitter gourd. J. Agric. Food Chem. 2008, 56, 10515–10520. [Google Scholar] [CrossRef]
- Bian, X.; Shen, F.; Chen, Y.; Wang, B.; Deng, M.; Meng, Y. PEGylation of alpha-momorcharin: Synthesis and characterization of novel anti-tumor conjugates with therapeutic potential. Biotechnol. Lett. 2010, 32, 883–890. [Google Scholar] [CrossRef]
- Nerurkar, P.; Ray, R.B. Bitter melon: Antagonist to cancer. Pharm. Res. 2010, 27, 1049–1053. [Google Scholar]
- Xu, B.; Dong, Y.; Zhang, H.-H.; Cui, H.-L.; Lu, Q. The antihyperglycemic effects of polysaccharide from Momordica charantia in STZ-induced diabetic mice. Acta Nutr. Sin. 2006, 28, 401–404. [Google Scholar]
- Kalupahana, N.S.; Moustaid-Moussa, N.; Claycombe, K.J. Immunity as a link between obesity and insulin resistance. Mol. Aspects Med. 2012, 33, 26–34. [Google Scholar] [CrossRef]
- Zitvogel, L.; Apetoh, L.; Ghiringhelli, F.; Kroemer, G. Immunological aspects of cancer chemotherapy. Nat. Rev. Immunol. 2008, 8, 59–73. [Google Scholar] [CrossRef]
- Jin, M.; Zhao, K.; Huang, Q.; Xu, C.; Shang, P. Isolation, structure and bioactivities of the polysaccharides from Angelica sinensis (Oliv.) Diels: A review. Carbohydr. Polym. 2012, 89, 713–722. [Google Scholar] [CrossRef]
- Zhu, X.L.; Chen, A.F.; Lin, Z.B. Ganoderma lucidum polysaccharides enhance the function of immunological effector cells in immunosuppressed mice. J. Ethnopharmacol. 2007, 111, 219–226. [Google Scholar] [CrossRef]
- Omarsdottir, S.; Freysdottir, J.; Olafsdottir, E.S. Immunomodulating polysaccharides from the lichen Thamnolia vermicularis var. subuliformis. Phytomedicine 2007, 14, 179–184. [Google Scholar] [CrossRef]
- Mizuno, M.; Shiomi, Y.; Minato, K.-I.; Kawakami, S.; Ashida, H.; Tsuchida, H. Fucogalactan isolated from Sarcodon aspratus elicits release of tumor necrosis factor-α and nitric oxide from murine macrophages. Immunopharmacology 2000, 46, 113–121. [Google Scholar] [CrossRef]
- Han, X.-Q.; Chan, B.C.L.; Yu, H.; Yang, Y.-H.; Hu, S.-Q.; Ko, C.-H.; Dong, C.-X.; Wong, C.-K.; Shaw, P.-C.; Fung, K.-P. Structural characterization and immuno-modulating activities of a polysaccharide from Ganoderma sinense. Int. J. Biol. Macromol. 2012, 51, 597–603. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, L.; Duan, X.; Liao, Z.; Ding, H.; Cheung, P.C. Novel highly branched water-soluble heteropolysaccharides as immunopotentiators to inhibit S-180 tumor cell growth in BALB/c mice. Carbohydr. Polym. 2012, 87, 427–434. [Google Scholar] [CrossRef]
- Ke, C.; Qiao, D.; Luo, J.; Li, Z.; Sun, Y.; Ye, H.; Zeng, X. Immunostimulatory activity and structure of polysaccharide from Streptococcus equi subsp. zooepidemicus. Int. J. Biol. Macromol. 2013, 57, 218–225. [Google Scholar] [CrossRef]
- Mizuno, T. Development of antitumor polysaccharides from mushroom fungi. Foods Food Ingred. J. Jpn. 1996, 167, 69–85. [Google Scholar]
- Wan, Z.-X.; Zhang, F.; Geng, Q.; Wang, P.-Y.; Zhou, H.; Zhang, Y.-M. Effect of orally administered bovine colostrum on cytokine production in vivo and in vitro in immunosuppressed mice. Int. Dairy J. 2010, 20, 522–527. [Google Scholar] [CrossRef]
- Yi, Y.; Liao, S.-T.; Zhang, M.-W.; Zhang, R.-F.; Deng, Y.-Y.; Yang, B.; Wei, Z.-C. Immunomodulatory activity of polysaccharide-protein complex of longan (Dimocarpus longan Lour.) pulp. Molecules 2011, 16, 10324–10336. [Google Scholar] [CrossRef]
- Chen, J.R.; Yang, Z.Q.; Hu, T.J.; Yan, Z.T.; Niu, T.X.; Wang, L.; Cui, D.A.; Wang, M. Immunomodulatory activity in vitro and in vivo of polysaccharide from Potentilla anserina. Fitoterapia 2010, 81, 1117–1124. [Google Scholar] [CrossRef]
- Bao, X.; Liu, C.; Fang, J.; Li, X. Structural and immunological studies of a major polysaccharide from spores of Ganoderma lucidum (Fr.) Karst. Carbohydr. Res. 2001, 332, 67–74. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, H.; Cai, G.; Guan, S.; Tong, H.; Yang, X.; Liu, J. Sulfated modification of the water-soluble polysaccharides from Polyporus albicans mycelia and its potential biological activities. Int. J. Biol. Macromol. 2009, 44, 14–17. [Google Scholar] [CrossRef]
- Iribe, H.; Koga, T. Augmentation of the proliferative response of thymocytes to phytohemagglutinin by the muramyl dipeptide. Cell. Immunol. 1984, 88, 9–15. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, K. Characteristic immunostimulation by MAP, a polysaccharide isolated from the mucus of the loach, Misgurnus anguillicaudatus. Carbohydr. Polym. 2005, 59, 75–82. [Google Scholar] [CrossRef]
- Sun, Y.X.; Liu, J.C. Purification, structure and immunobiological activity of a water-soluble polysaccharide from the fruiting body of Pleurotus ostreatus. Bioresour. Technol. 2009, 100, 983–986. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, H.; Zhang, X.; Tong, H.; Liu, J. Structural elucidation and immunological activity of a polysaccharide from the fruiting body of Armillaria mellea. Bioresour. Technol. 2009, 100, 1860–1863. [Google Scholar] [CrossRef]
- Sun, Y.X.; Liu, J.C. Structural characterization of a water-soluble polysaccharide from the roots of Codonopsis pilosula and its immunity activity. Int. J. Biol. Macromol. 2008, 43, 279–282. [Google Scholar] [CrossRef]
- Yang, C.; He, N.; Ling, X.; Ye, M.; Zhang, C.; Shao, W.; Yao, C.; Wang, Z.; Li, Q. The isolation and characterization of polysaccharides from longan pulp. Sep. Purif. Technol. 2008, 63, 226–230. [Google Scholar] [CrossRef]
- Yi, Y.; Liao, S.T.; Zhang, M.W.; Shi, J.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C. Physicochemical characteristics and immunomodulatory activities of three polysaccharide-protein complexes of longan pulp. Molecules 2011, 16, 6148–6164. [Google Scholar] [CrossRef]
- Zhang, W. Biochemical Technology of Carbohydrate Complexes; Zhejiang University Press: Hangzhou, China, 1994. [Google Scholar]
- Yi, Y.; Zhang, M.-W.; Liao, S.-T.; Zhang, R.-F.; Deng, Y.-Y.; Wei, Z.-C.; Tang, X.-J.; Zhang, Y. Structural features and immunomodulatory activities of polysaccharides of longan pulp. Carbohydr. Polym. 2012, 87, 636–643. [Google Scholar] [CrossRef]
- Xu, R.; Ye, H.; Sun, Y.; Tu, Y.; Zeng, X. Preparation, preliminary characterization, antioxidant, hepatoprotective and antitumor activities of polysaccharides from the flower of tea plant (Camellia sinensis). Food Chem. Toxicol. 2012, 50, 2473–2480. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, N.; Yang, B.; Jiang, Y.; Zhang, G. Structural characterization of water-soluble polysaccharides from Opuntia monacantha cladodes in relation to their anti-glycated activities. Food Chem. 2007, 105, 1480–1486. [Google Scholar]
- Yang, W.; Pei, F.; Shi, Y.; Zhao, L.; Fang, Y.; Hu, Q. Purification, characterization and anti-proliferation activity of polysaccharides from Flammulina velutipes. Carbohydr. Polym. 2012, 88, 474–480. [Google Scholar] [CrossRef]
- Guoling, L.; Zhengxiang, N.; Honghui, G. Isolation and Characterization of Polysaccharides from Momordica charantia L. Fruits. Food Sci. 2010, 31, 5. [Google Scholar]
- Orlovskaya, T. Carbohydrates and lipids from Momordica charantia. Chem. Nat. Compd. 2008, 44, 227–228. [Google Scholar] [CrossRef]
- Thakur, M.; Weng, A.; Fuchs, H.; Sharma, V.; Bhargava, C.S.; Chauhan, N.S.; Dixit, V.K.; Bhargava, S. Rasayana properties of Ayurvedic herbs: Are polysaccharides a major contributor. Carbohydr. Polym. 2012, 87, 3–15. [Google Scholar] [CrossRef]
- Sekhon, B.K.; Sze, D.M.-Y.; Chan, W.K.; Fan, K.; Li, G.Q.; Moore, D.E.; Roubin, R.H. PSP activates monocytes in resting human peripheral blood mononuclear cells: Immunomodulatory implications for cancer treatment. Food Chem. 2013, 138, 2201–2209. [Google Scholar] [CrossRef]
- Leiro, J.M.; Castro, R.; Arranz, J.A.; Lamas, J. Immunomodulating activities of acidic sulphated polysaccharides obtained from the seaweed Ulva rigida C. Agardh. Int. Immunopharmacol. 2007, 7, 879–888. [Google Scholar] [CrossRef]
- Qiao, D.; Luo, J.; Ke, C.; Sun, Y.; Ye, H.; Zeng, X. Immunostimulatory activity of the polysaccharides from Hyriopsis cumingii. Int. J. Biol. Macromol. 2010, 47, 676–680. [Google Scholar]
- Zhang, D.; Wang, C.; Wu, H.; Xie, J.; Du, L.; Xia, Z.; Cai, J.; Huang, Z.; Wei, D. Three sulphated polysaccharides isolated from the mucilage of mud snail, Bullacta exarata philippi: Characterization and antitumour activity. Food Chem. 2013, 138, 306–314. [Google Scholar] [CrossRef]
- Staub, A. Removeal of protein—Sevag method. Methods Carbohydr. Chem. 1956, 5, 5–6. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Pebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Xu, W.Q. Study on The Structure Feature, Biological Activity and Anti-tumor Mechanism of Polysaccharide from Spores of Tremella fuciformis Berk. Ph.D. Thesis, Tianjin University, Tianjin, China, June 2006. [Google Scholar]
- Kowalski, J. Effect of enkephalins and endorphins on cytotoxic activity of natural killer cells and macrophages/monocytes in mice. Eur. J. Pharmacol. 1997, 326, 251–255. [Google Scholar] [CrossRef]
- Chen, X.; Jin, J.; Tang, J.; Wang, Z.; Wang, J.; Jin, L.; Lu, J. Extraction, purification, characterization and hypoglycemic activity of a polysaccharide isolated from the root of Ophiopogon japonicus. Carbohydr. Polym. 2011, 83, 749–754. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the polysaccharides described herein are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Deng, Y.-Y.; Yi, Y.; Zhang, L.-F.; Zhang, R.-F.; Zhang, Y.; Wei, Z.-C.; Tang, X.-J.; Zhang, M.-W. Immunomodulatory Activity and Partial Characterisation of Polysaccharides from Momordica charantia. Molecules 2014, 19, 13432-13447. https://doi.org/10.3390/molecules190913432
Deng Y-Y, Yi Y, Zhang L-F, Zhang R-F, Zhang Y, Wei Z-C, Tang X-J, Zhang M-W. Immunomodulatory Activity and Partial Characterisation of Polysaccharides from Momordica charantia. Molecules. 2014; 19(9):13432-13447. https://doi.org/10.3390/molecules190913432
Chicago/Turabian StyleDeng, Yuan-Yuan, Yang Yi, Li-Fang Zhang, Rui-Fen Zhang, Yan Zhang, Zhen-Cheng Wei, Xiao-Jun Tang, and Ming-Wei Zhang. 2014. "Immunomodulatory Activity and Partial Characterisation of Polysaccharides from Momordica charantia" Molecules 19, no. 9: 13432-13447. https://doi.org/10.3390/molecules190913432