
Auraptene Acts as an Anti-Inflammatory Agent in the Mouse Brain


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
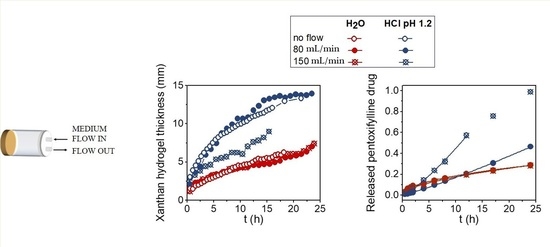
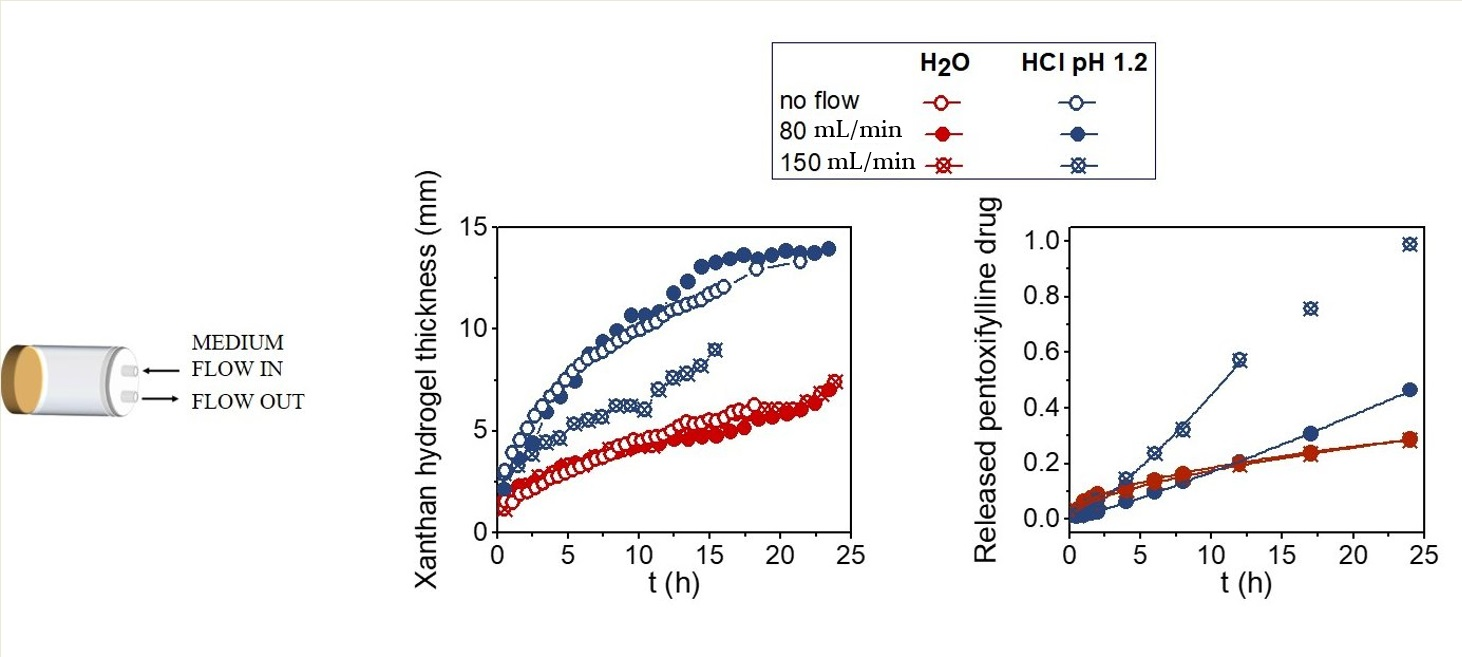
2. Results and Discussion
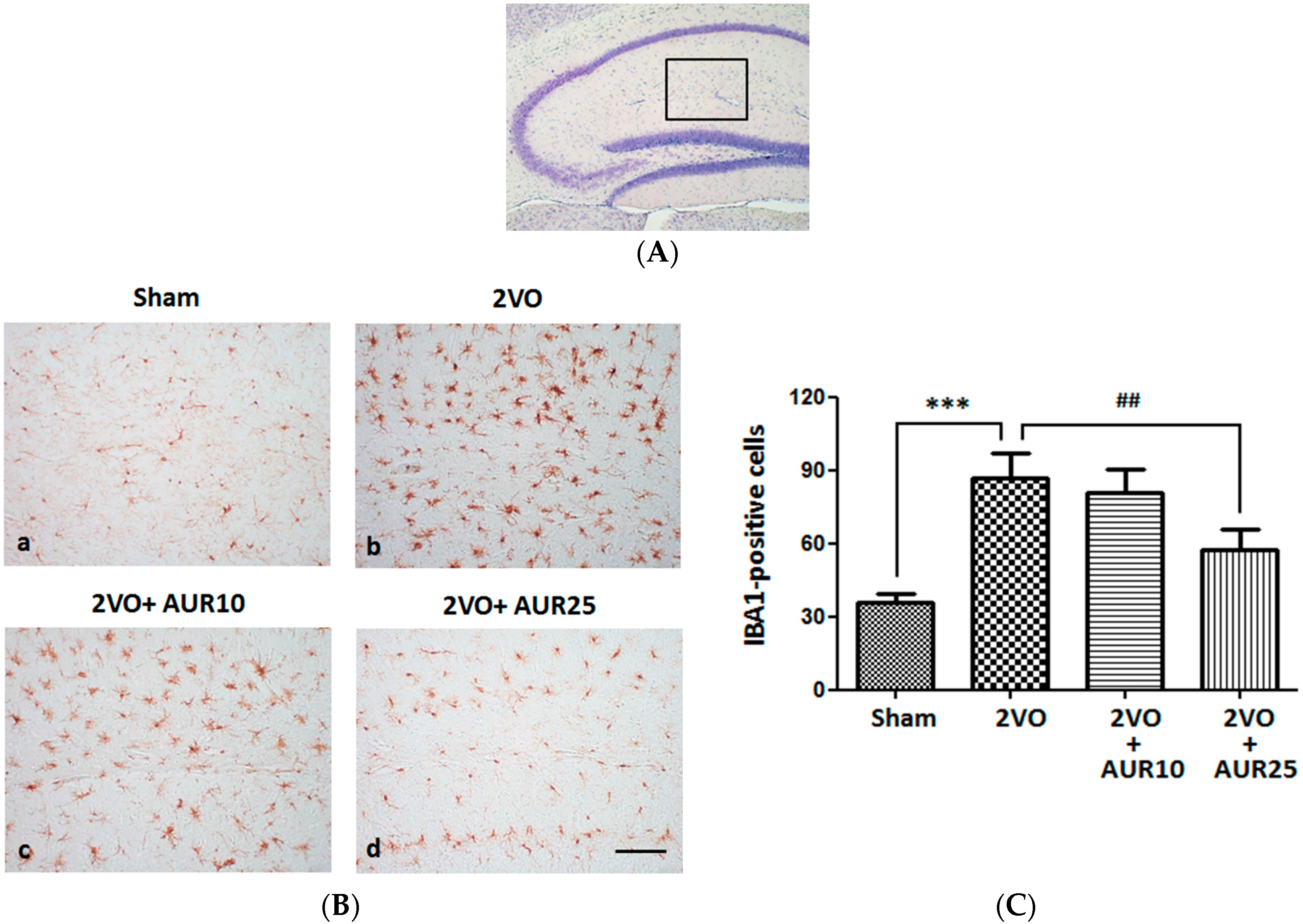
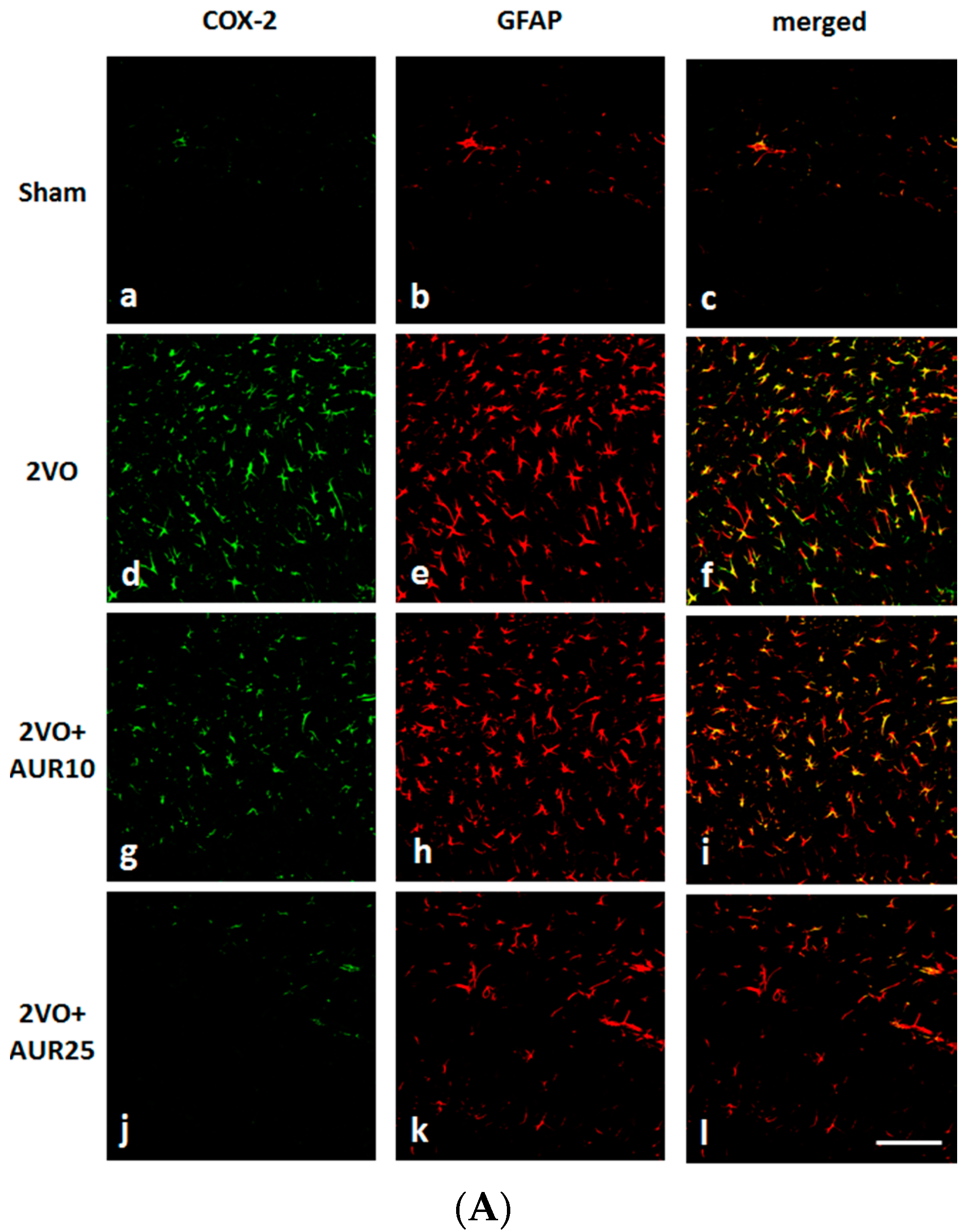
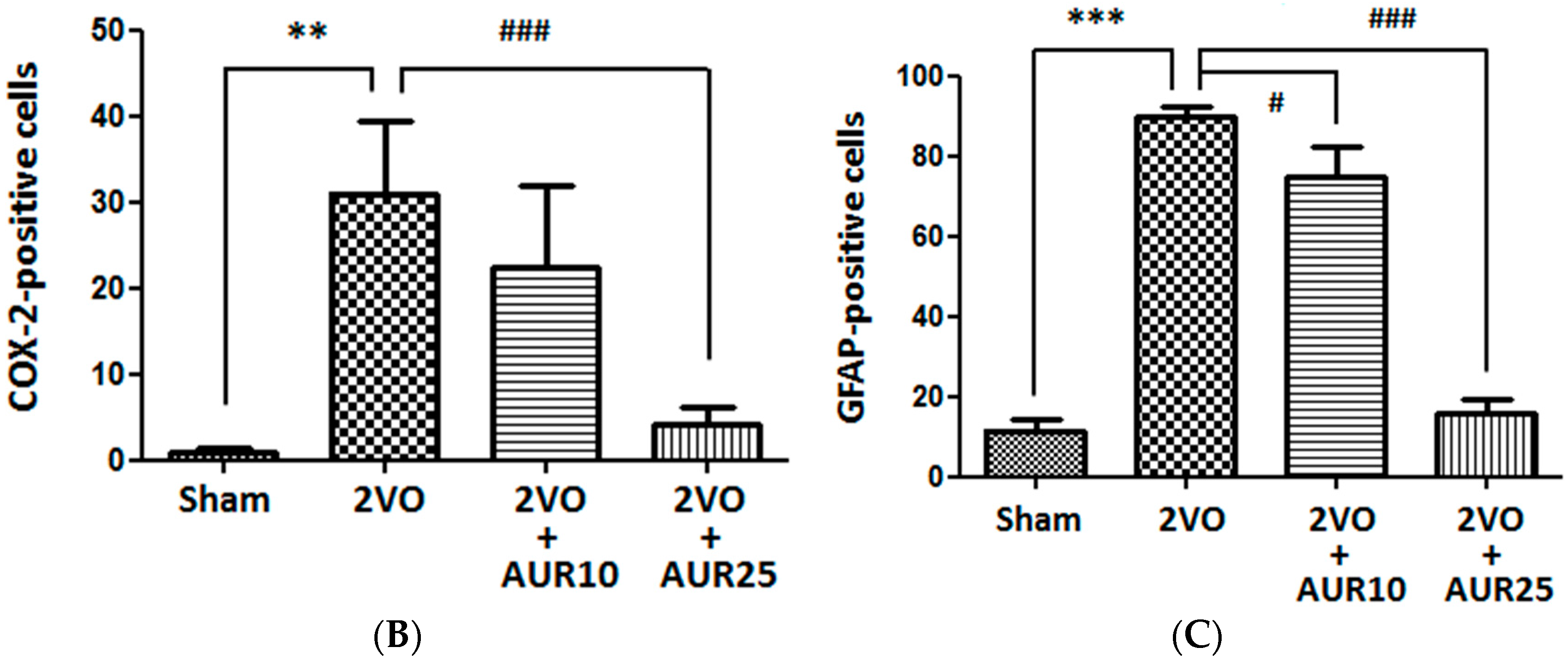
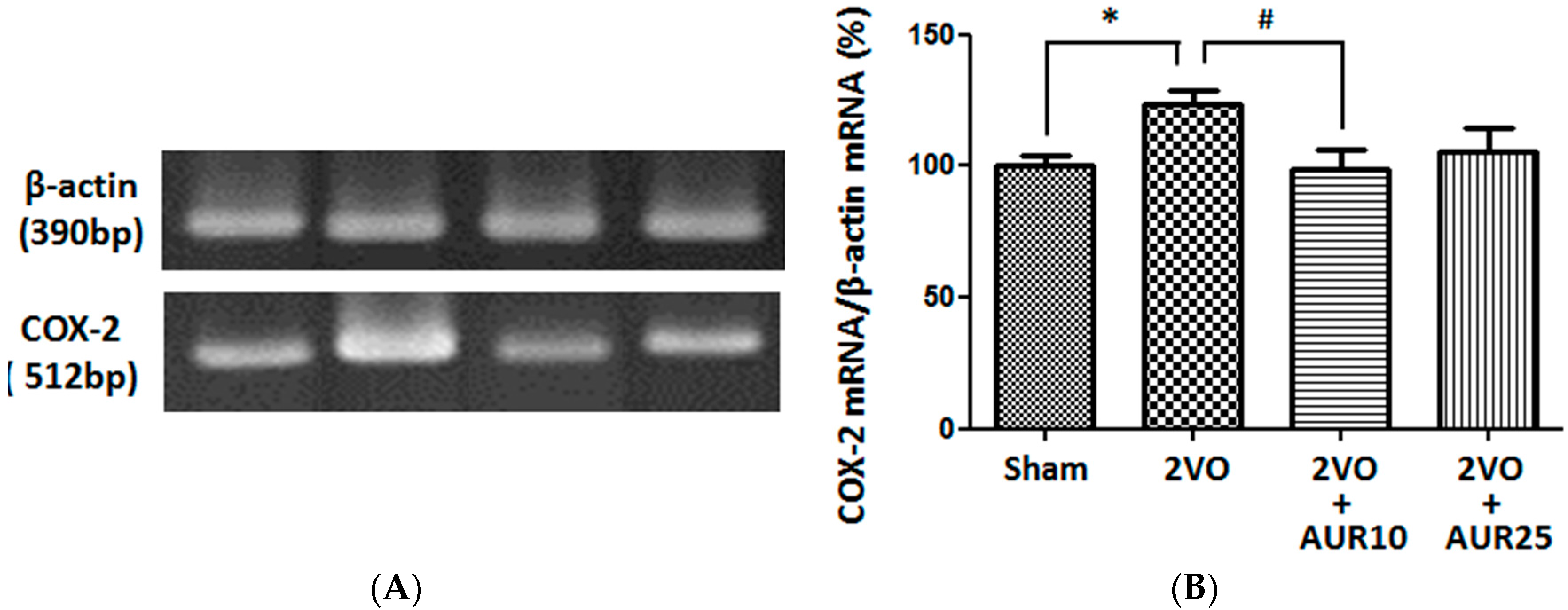
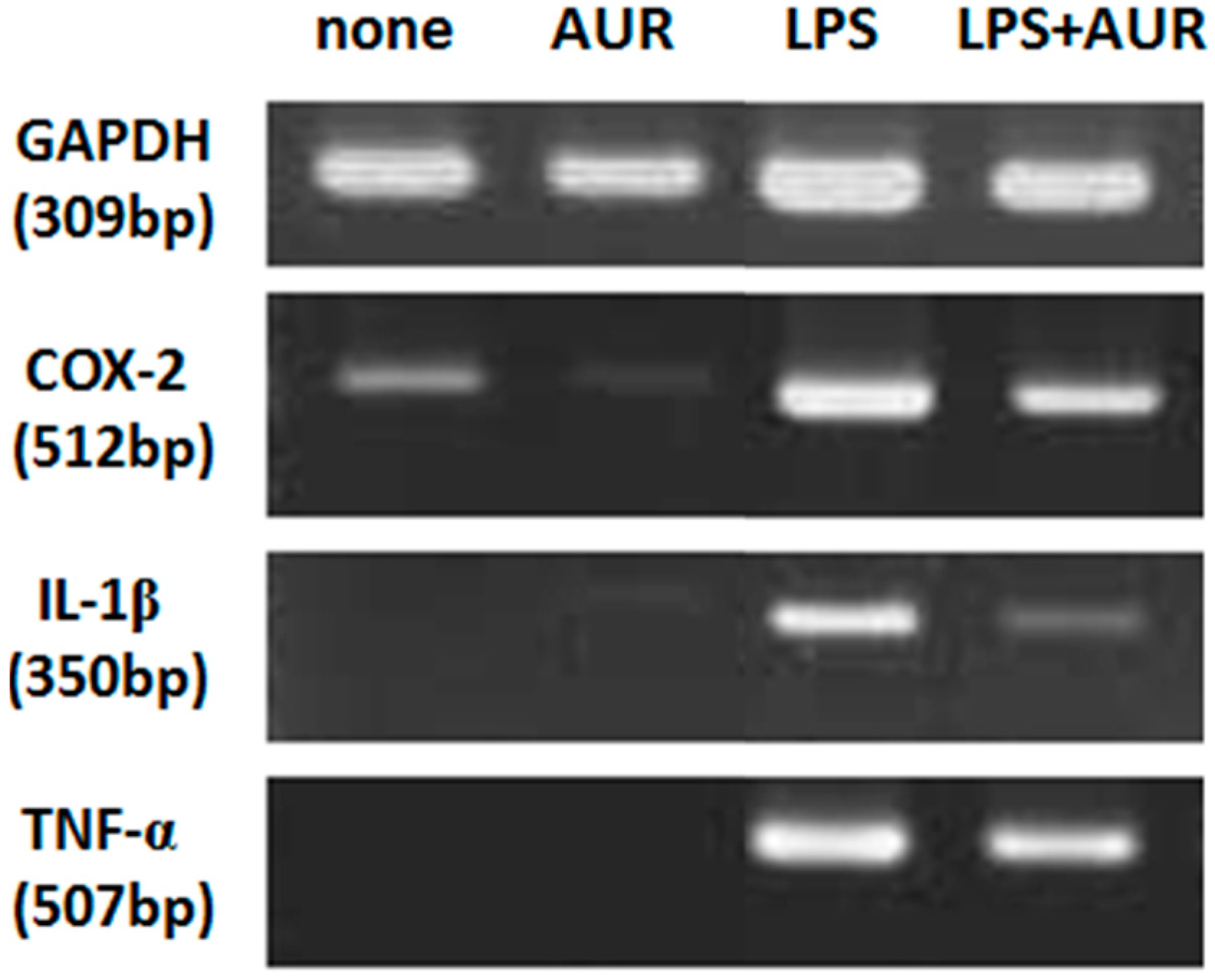
2.1. Suppressive Effects of AUR on Inflammation in the Ischemic Brain





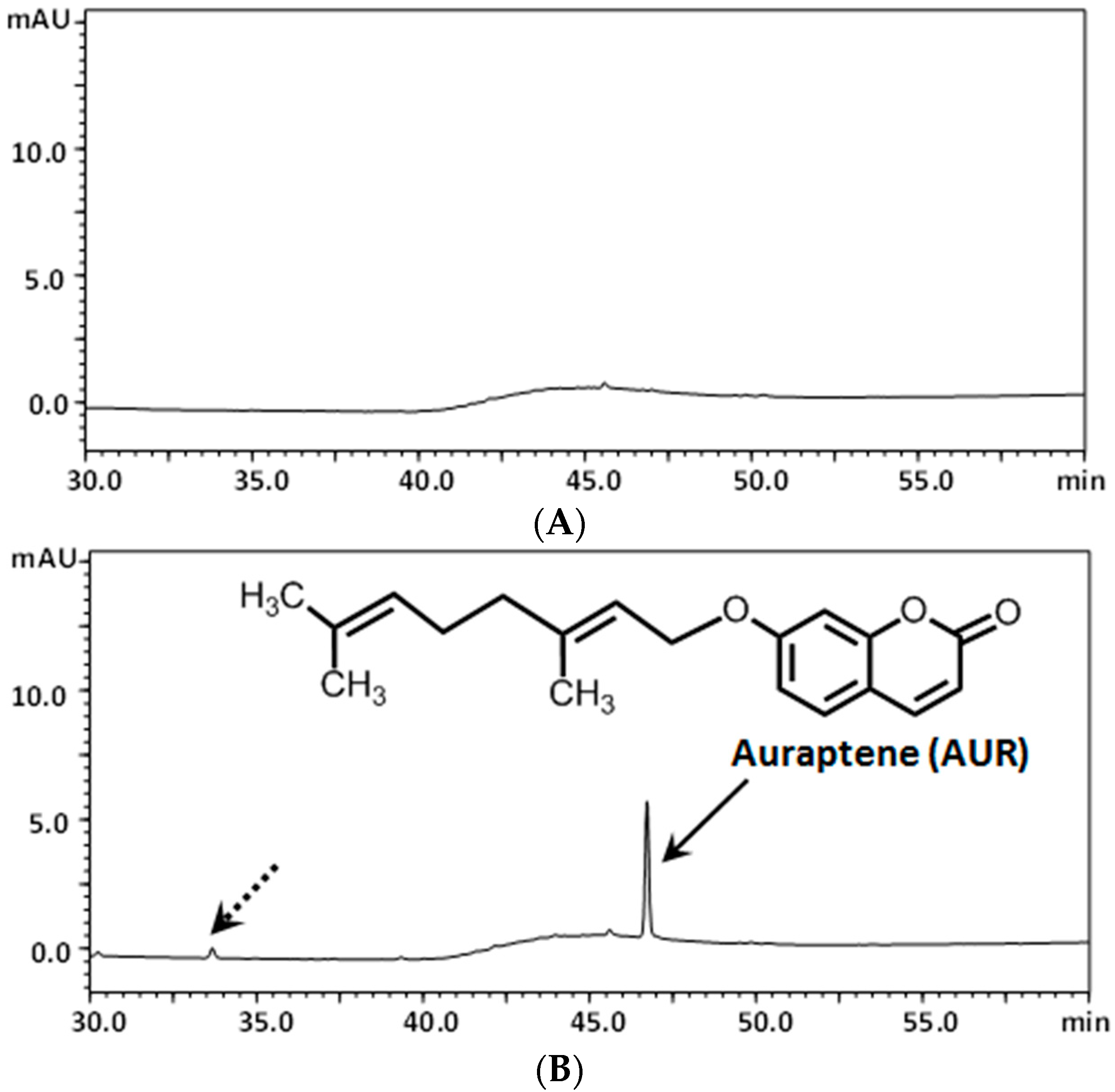
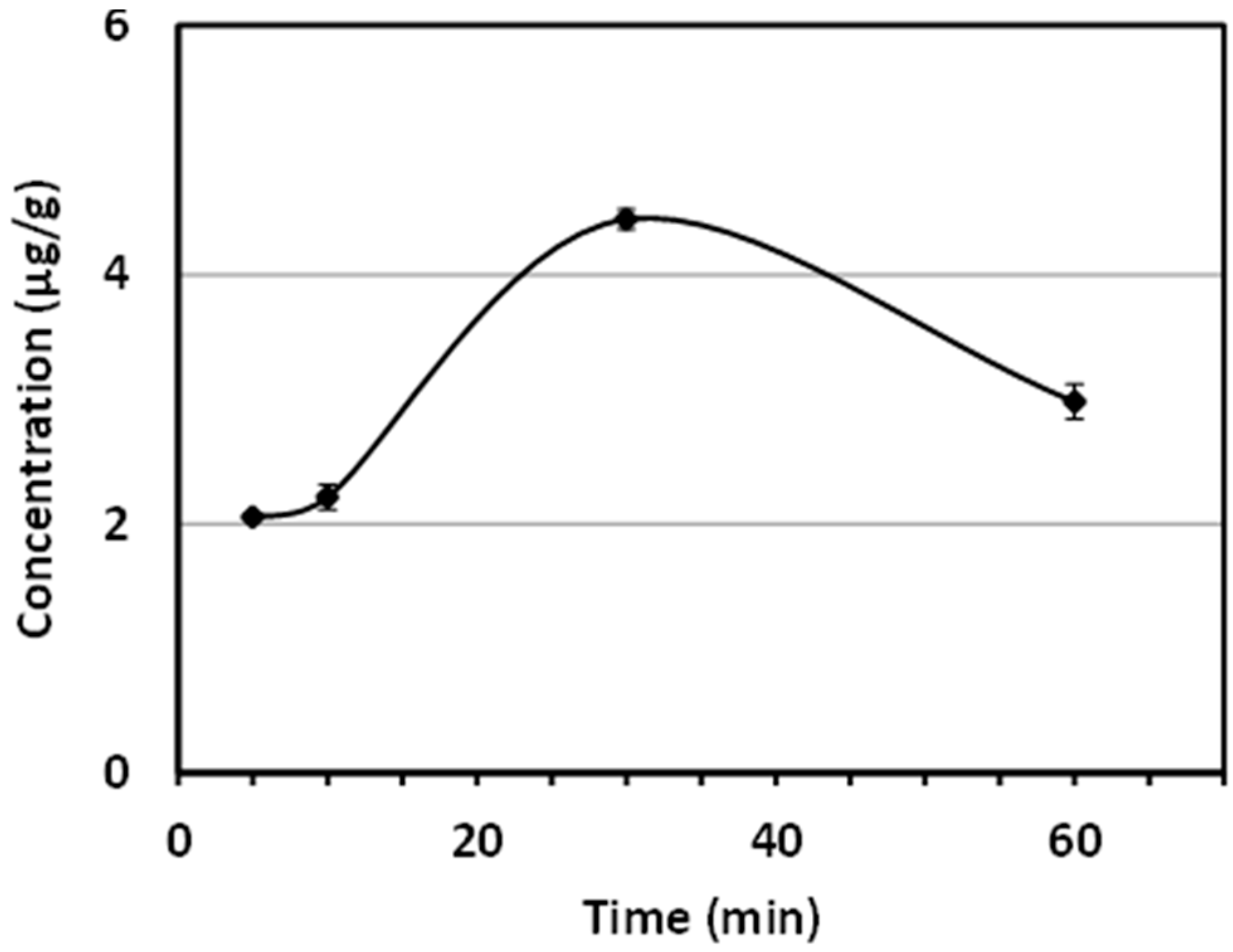
2.2. Determination of AUR in the Mouse Brain after Its i.p. Administration


3. Experimental Section
3.1. Animal Preparation
3.2. Brain Sample Preparation for HPLC/UV
3.3. Assessment of AUR via HPLC/UV
3.4. Procedures for Ischemic Surgery
3.5. Immunohistochemistry
3.6. Cell Culture
3.7. RT-PCR Procedures
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Murakami, A.; Kuki, W.; Takahashi, Y.; Yonei, H.; Nakamura, Y.; Ohto, Y.; Ohigashi, H.; Koshimizu, K. Auraptene, a citrus coumarin, inhibits 12-O-tetradecanoylphorbol-13-acetate-induced tumor promotion in ICR mouse skin, possibly through suppression of superoxide generation in leukocytes. Jpn. J. Cancer Res. 1997, 88, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kawabata, K.; Kakumoto, M.; Hara, A.; Murakami, A.; Kuki, W.; Takahashi, Y.; Yonei, H.; Maeda, M.; Ota, T.; et al. Citrus auraptene exerts dose-dependent chemopreventive activity in rat large bowel tumorigenesis; the inhibition correlates with suppression of cell proliferation and lipid peroxidation and with induction of phase II drug-metabolizing enzymes. Cancer Res. 1998, 58, 2550–2556. [Google Scholar] [PubMed]
- Murakami, A.; Nakamura, Y.; Tanaka, T.; Kawabata, K.; Takahashi, D.; Koshimizu, K.; Ohigashi, H. Suppression by citrus auraptene of phorbol ester- and endotoxin-induced inflammatory responses: Role of attenuation of leukocyte activation. Carcinogenesis 2000, 21, 1843–1850. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yasui, Y.; Ishigamori-Suzuki, R.; Oyama, T. Citrus compounds inhibit inflammation- and obesity-related colon carcinogenesis in mice. Nutr. Cancer 2008, 60 (Suppl. S1), 70–80. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Hirai, S.; Goto, T.; Sakamoto, T.; Takahashi, N.; Yano, M.; Sasaki, T.; Yu, R.; Kawada, T. Auraptene suppresses inflammatory responses in activated RAW264 macrophages by inhibiting p38 mitogen-activated protein kinase activation. Mol. Nutr. Food Res. 2013, 57, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Ma, Z.; Peng, S.; Deng, X. Anti-inflammatory effect of auraptene extracted from Trifoliate orange (Poncitrus Trifoliate) on LPS-stimulated RAW 264.4 cells. Inflammation 2013, 36, 1525–1532. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Shigemori, T.; Ohigashi, H. Zingiberaceous and citrus constituents, 1′-acetoxychavicol acetate, zerumbone, auraptene, and nobiletin, suppress lipopolysaccharide-induced cyclooxygenase-2 expression in RAW264.7 murine macrophages through different modes of action. J. Nutr. 2005, 135, 2987S–2992S. [Google Scholar] [PubMed]
- Okuyama, S.; Minami, S.; Shimada, N.; Makihata, N.; Nakajima, M.; Furukawa, Y. Anti-inflammatory and neuroprotective effects of auraptene, a citrus coumarin, following cerebral global ischemia in mice. Eur. J. Pharmacol. 2013, 699, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Yamamoto, K.; Mori, H.; Toyoda, N.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Sugawara, S.; Sudo, M.; Nakajima, M.; et al. Auraptene in the peels of Citrus kawachiensis (kawachi bankan) ameliorates lipopolysaccharide (LPS)-induced inflammation in the mouse brain. Evid. Based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Matsumura, K.; Yamagata, K.; Watanabe, Y. Induction by lipopolysaccharide of cyclooxygenase-2 mRNA in rat brain; its possible role in the febrile response. Brain Res. 1995, 697, 187–196. [Google Scholar] [CrossRef]
- Weinstein, J.R.; Koerner, I.P.; Möller, T. Microglia in ischemic brain injury. Future Neurol. 2010, 5, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Tanaka, K.; Suzuki, S.; Dembo, T.; Fukuuchi, Y. Enhanced expression of iba1, ionized calcium-binding adapter molecules 1, after transient focal cerebral ischemia in rat brain. Stroke 2001, 32, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Koistinaho, J.; Koponen, S.; Chan, P.H. Expression of cyclooxygenase-2 mRNA after global ischemia is regulated by AMP receptors and glucocorticoids. Stroke 1999, 30, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wu, H.; Wang, J.; Wang, J. Expression and cellular localization of cyclooxygenases and prostaglandin E synthases in the hemorrhagic brain. J. Neuroinflamm. 2011, 8, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.-B.; Voloboueva, L.A.; Xu, L.-J.; Giffard, R.G. Selective dysfunction of hippocampal CA1 astrocytes contributes to delayed neuronal damage after transient forebrain ischemia. J. Neurosci. 2007, 27, 4253–4260. [Google Scholar] [CrossRef] [PubMed]
- Katsuta, K.; Umemura, K.; Ueyama, N.; Matsuoka, N. Pharmacological evidence for a correlation between hippocampal CA1 cell damage and hyperlocomotion following global cerebral ischemia in gerbils. Eur. J. Pharmacol. 2003, 467, 103–109. [Google Scholar] [CrossRef]
- Okuyama, S.; Shimada, N.; Kaji, M.; Morita, M.; Miyoshi, K.; Minami, S.; Amakura, Y.; Yoshimura, M.; Yoshida, T.; Watanabe, S.; et al. Heptamethoxyflavone, a citrus flavonoid, enhances brain-derived neurotrophic factor production and neurogenesis in the hippocampus following cerebral global ischemia in mice. Neurosci. Lett. 2012, 528, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Soromou, L.W.; Zhang, Z.; Li, R.; Chen, N.; Guo, W.; Huo, M.; Guan, S.; Lu, J.; Deng, X. Regulation of inflammatory cytokines in lipopolysaccharide-stimulated RAW 264.7 murine macrophage by 7-O-methyl-naringenin. Molecules 2012, 17, 3574–3585. [Google Scholar] [CrossRef] [PubMed]
- Bramlett, H.M.; Dietrich, W.D. Pathophysiology of cerebral ischemia and brain trauma: Similarities and differences. J. Cereb. Blood Flow Metab. 2004, 24, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Ladecola, D.; Anrather, J. The immunology of stroke: From mechanisms to translation. Nat. Med. 2012, 17, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tang, X.N.; Yenari, M.A. The inflammatory response in stroke. J. Neuroimmunol. 2007, 184, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.P.; Praticò, D. Novel therapeutic opportunities for Alzheimer’s disease: Focus on nonsteroidal anti-inflammatory drugs. FASEB J. 2005, 19, 1592–1601. [Google Scholar] [CrossRef] [PubMed]
- Nitta, A.; Ito, M.; Fukumitsu, H.; Ohmiya, M.; Ito, H.; Sometani, A.; Nomoto, H.; Furukawa, Y.; Furukawa, S. 4-Methylcatechol increases brain-derived neurotrophic factor content and mRNA expression in cultured brain cells and in rat brain in vivo. J. Pharmcol. Exp. Ther. 1999, 291, 1276–1283. [Google Scholar]
- Furukawa, Y.; Watanabe, S.; Okuyama, S.; Nakajima, M. Neurotrophic effect of citrus auraptene: Neuritogenic activity in PC12 cells. Int. J. Mol. Sci. 2012, 13, 5338–5347. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Makihata, N.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Nakajima, M.; Furukawa, Y. Oenothein B suppresses lipopolysaccharide (LPS)-induced inflammation in the mouse brain. Int. J. Mol. Sci. 2013, 14, 9767–9778. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okuyama, S.; Morita, M.; Kaji, M.; Amakura, Y.; Yoshimura, M.; Shimamoto, K.; Ookido, Y.; Nakajima, M.; Furukawa, Y. Auraptene Acts as an Anti-Inflammatory Agent in the Mouse Brain. Molecules 2015, 20, 20230-20239. https://doi.org/10.3390/molecules201119691
Okuyama S, Morita M, Kaji M, Amakura Y, Yoshimura M, Shimamoto K, Ookido Y, Nakajima M, Furukawa Y. Auraptene Acts as an Anti-Inflammatory Agent in the Mouse Brain. Molecules. 2015; 20(11):20230-20239. https://doi.org/10.3390/molecules201119691
Chicago/Turabian StyleOkuyama, Satoshi, Mayu Morita, Miki Kaji, Yoshiaki Amakura, Morio Yoshimura, Koji Shimamoto, Yu Ookido, Mitsunari Nakajima, and Yoshiko Furukawa. 2015. "Auraptene Acts as an Anti-Inflammatory Agent in the Mouse Brain" Molecules 20, no. 11: 20230-20239. https://doi.org/10.3390/molecules201119691