
Derinat Protects Skin against Ultraviolet-B (UVB)-Induced Cellular Damage

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
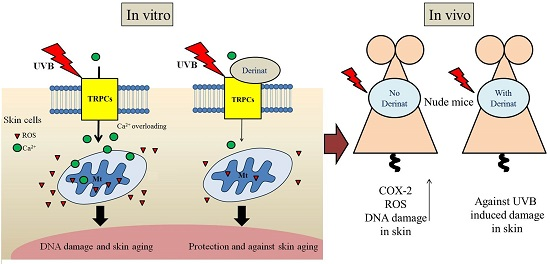
2.1. Derinat Protected Skin Cells from Damage Induced by UVB Irradiation

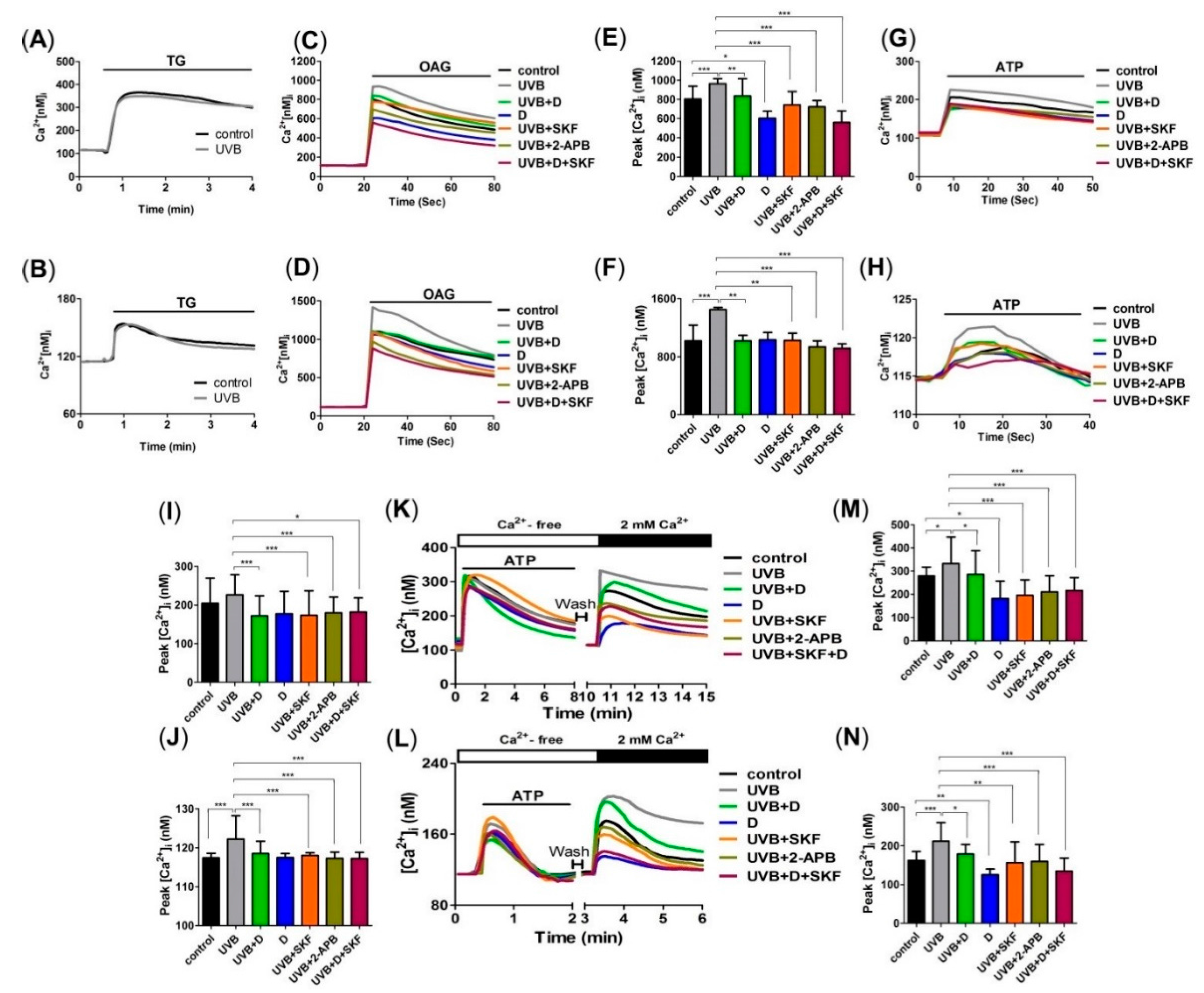
2.2. Derinat Decreased Intracellular Ca2+ Elevation in Skin Cells Exposed to UVB



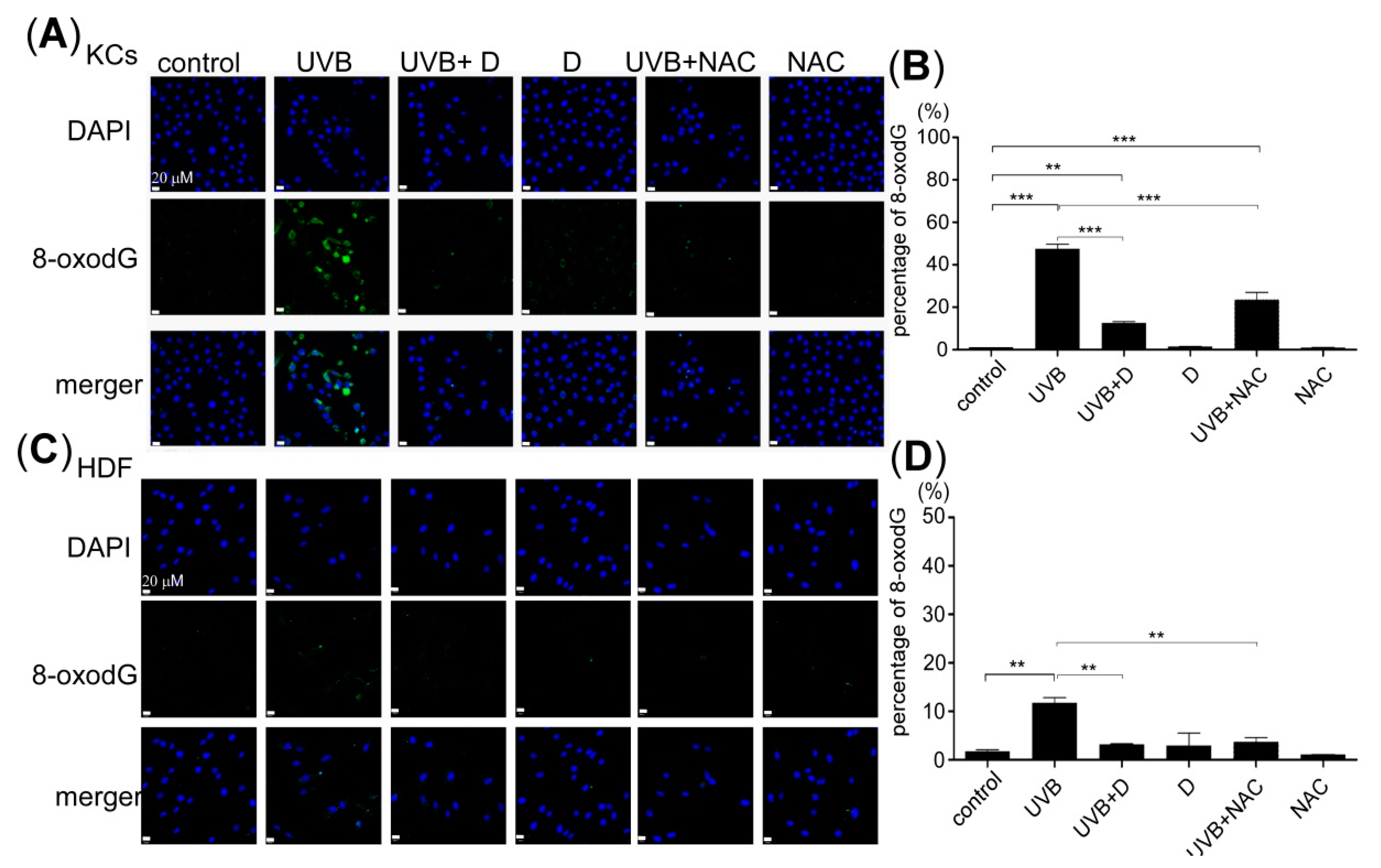
2.3. Derinat Reduced Oxidative Stress Accumulation Following the UVB Irradiation of Skin Cells
2.4. Derinat Protected Skin Cells Against UVB Induced DNA Damage


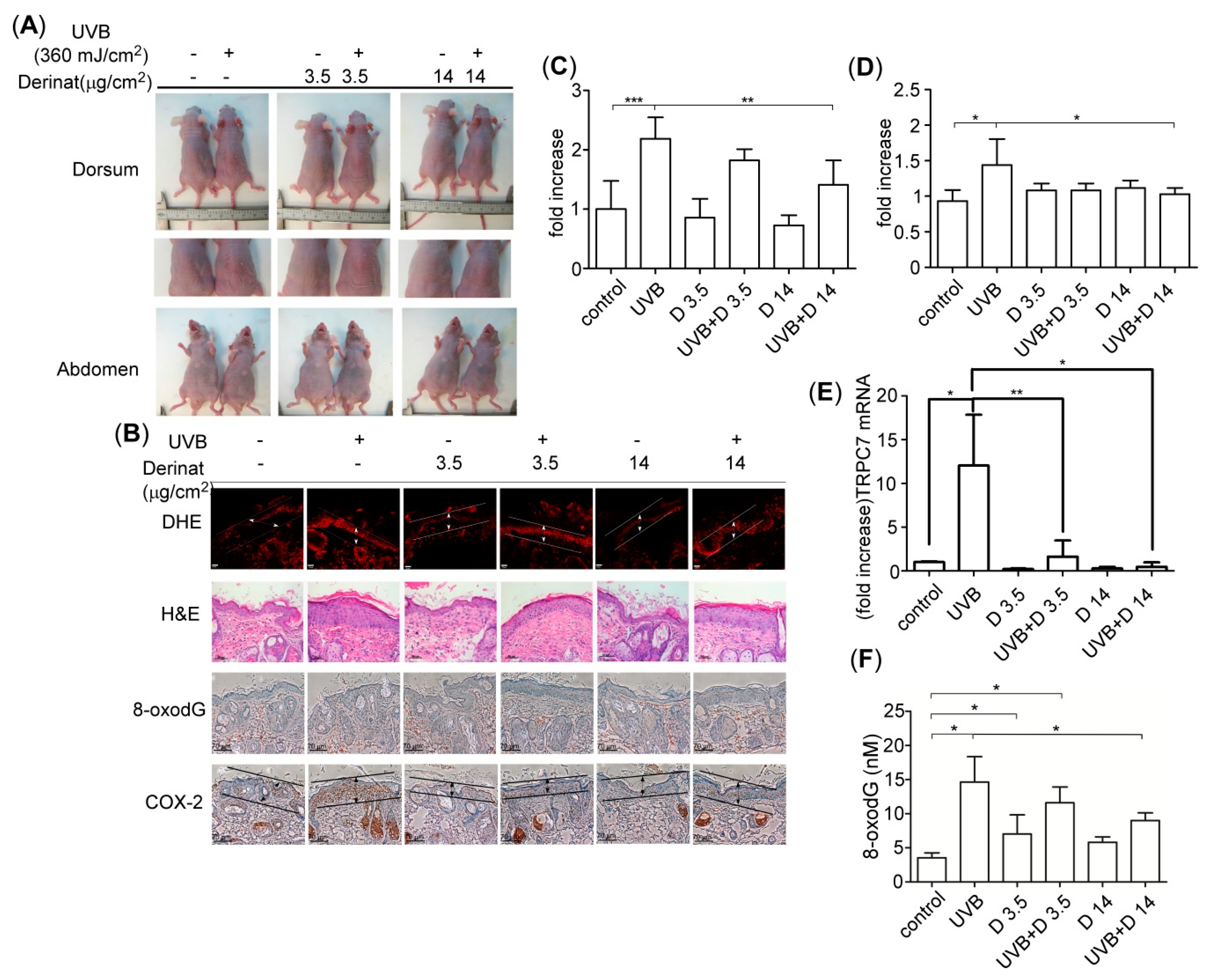
2.5. Derinat Protected BALB/c-nu Mice from UVB-Induced Skin Damage

3. Experimental Section
3.1. Cell Culture
3.2. Cell Viability Assay
3.3. Calcium Imaging
3.4. Immunofluorescence Assay
3.5. Analysis of Intracellular ROS Production in Skin Tissue
3.6. Immunohistochemistry of Skin Tissue
3.7. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Akhalaya, M.Y.; Maksimov, G.V.; Rubin, A.B.; Lademann, J.; Darvin, M.E. Molecular action mechanisms of solar infrared radiation and heat on human skin. Ageing Res. Rev. 2014, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Yang, Y.L.; Yang, Z.; Tu, Y.; Cheng, L.; Chen, B.; Xia, J.P.; Sun, W.L.; Su, Z.L.; He, L.; et al. Perifosine sensitizes UVB-induced apoptosis in skin cells: New implication of skin cancer prevention? Cell. Signal. 2012, 24, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Grether-Beck, S.; Felsner, I.; Koehler, T.; Farwick, M.; Lersch, P.; Rawlings, A.V.; Krutmann, J. Topical ceramides neither enhance UVB-induced apoptosis in normal human keratinocytes nor affect viability in UVB-irradiated reconstructed human epidermis. Exp. Dermatol. 2014, 23, 853–855. [Google Scholar] [CrossRef] [PubMed]
- Kulms, D.; Schwarz, T. Independent contribution of three different pathways to ultraviolet-B-induced apoptosis. Biochem. Pharmacol. 2002, 64, 837–841. [Google Scholar] [CrossRef]
- Darvin, M.E.; Gersonde, I.; Albrecht, H.; Sterry, W.; Lademann, J. In vivo Raman spectroscopic analysis of the influence of UV radiation on carotenoid antioxidant substance degradation of the human skin. Laser Methods Biol. Med. 2006, 16, 833–837. [Google Scholar] [CrossRef]
- He, Y.D.; Liu, Y.T.; Lin, Q.X.; Zhu, J.; Zhang, Y.; Wang, L.Y.; Ren, X.L.; Ye, X.Y. Polydatin suppresses ultraviolet B-induced cyclooxygenase-2 expression in vitro and in vivo via reduced production of reactive oxygen species. Br. J. Dermatol. 2012, 167, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive compounds from natural resources against skin aging. Phytomed. Int. J. Phytother. Phytopharm. 2011, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.P.; Kim, H.G.; Han, E.H.; Choi, J.H.; Park, B.H.; Jung, K.H.; Shin, Y.C.; Jeong, H.G. N-Acetylglucosamine suppress collagenases activation in ultraviolet B-irradiated human dermal fibroblasts: Involvement of calcium ions and mitogen-activated protein kinases. J. Dermatol. Sci. 2011, 63, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Tigges, J.; Haarmann-Stemmann, T.; Vogel, C.F.; Grindel, A.; Hubenthal, U.; Brenden, H.; Grether-Beck, S.; Vielhaber, G.; Johncock, W.; Krutmann, J.; et al. The new aryl hydrocarbon receptor antagonist E/Z-2-benzylindene-5,6-dimethoxy-3,3-dimethylindan-1-one protects against UVB-induced signal transduction. J. Investig. Dermatol. 2014, 134, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Sviatkina, O.I.; Balashov, V.P.; Balykova, L.A.; Shchukin, S.A. Anti-arrhythmia activity of derinat in an experiment. Eksp. Klin. Farmakol. 2004, 67, 22–24. [Google Scholar] [PubMed]
- Wang, Y.N.; Wu, W.; Chen, H.C.; Fang, H. Genistein protects against UVB-induced senescence-like characteristics in human dermal fibroblast by p66Shc down-regulation. J. Dermatol. Sci. 2010, 58, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Gromov, M.I.; Pivovarova, L.P. Use of immunomodulator derinat in the treatment of patients with surgical sepsis in traumatic shock. Vestn. Khir. Im. I. I. Grek. 2002, 161, 45–48. [Google Scholar] [PubMed]
- Gora, N.V.; Bairakova, A.L.; Kozlov, L.V. Enzyme immunoassay of masked complement component C4 deficiency in patients with urogenital Chlamydia infection. Zh. Mikrobiol. Epidemiol. Immunobiol. 2011, 3, 76–80. [Google Scholar] [PubMed]
- Zemskov, A.M.; Zemskov, V.M.; Tokmakov, A.I. Clinical efficacy of immunostimulating drugs in treatment of purulent infections. Khirurgiia 2011, 2, 4–10. [Google Scholar] [PubMed]
- Mangushev, A.R.; Rafailov, V.V.; Svatko, L.G. Clinical efficiency of derinat used to treat chronic adenoiditis in children. Vestn. Otorinolaringol. 2008, 6, 33–34. [Google Scholar] [PubMed]
- Zemskov, A.M.; Kiselev, A.V.; Koveshnikov, V.V. Immune status of patients with exacerbation of chronic suppurative otitis media and its correction. Vestn. Otorinolaringol. 2010, 5, 38–40. [Google Scholar] [PubMed]
- Leonavičienė, L.; Bernotienė, E.; Bradūnaitė, R.; Vaitkienė, D.; Redaitienė, E.; Astrauskas, V. Antiarthritic and hepatoprotective effect of derinat on adjuvant arthritis in rats. Acta Med. Litu. 2006, 13, 236–244. [Google Scholar]
- Paiva, C.N.; Bozza, M.T. Are ROS always detrimental to pathogens? Antioxid. Redox Signal. 2013. [Google Scholar] [CrossRef]
- Okayama, Y. Oxidative stress in allergic and inflammatory skin diseases. Curr. Drug Targets Inflamm. Allergy 2005, 4, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-inducing bacterial species in the stomach. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2010, 59, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H.; Izutsu, Y.; Yahagi, S.; Okano, Y. Reactive oxygen species in HaCaT keratinocytes after UVB irradiation are triggered by intracellular Ca(2+) levels. J. Investig. Dermatol. Symp. Proc. 2009, 14, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.J.; Chu, A.W.; Lu, Z.F.; Pan, M.H.; Che, D.F.; Zhou, X.J. Ultraviolet B-induced alterations of the skin barrier and epidermal calcium gradient. Exp. Dermatol. 2007, 16, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W. L.; Yoshioka, T. Role of TRP channels in the induction of heat shock proteins (Hsps) by heating skin. Biophysics 2015, 11, 25–32. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Montell, C. TRP channels. Ann. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Punnonen, K.; Yuspa, S.H. Ultraviolet light irradiation increases cellular diacylglycerol and induces translocation of diacylglycerol kinase in murine keratinocytes. J. Investig. Dermatol. 1992, 99, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Rae, M.G.; Hilton, J.; Sharkey, J. Putative TRP channel antagonists, SKF 96365, flufenamic acid and 2-APB, are non-competitive antagonists at recombinant human alpha1beta2gamma2 GABA(A) receptors. Neurochem. Int. 2012, 60, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, J.R.; Wang, Y.; Li, C.L.; Lu, D.; Guan, S.M.; Chen, J. Effects of a non-selective TRPC channel blocker, SKF-96365, on melittin-induced spontaneous persistent nociception and inflammatory pain hypersensitivity. Neurosci. Bull. 2012, 28, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. SnapShot: mammalian TRP channels. Cell 2007. [Google Scholar] [CrossRef] [PubMed]
- Fajardo, O.; Meseguer, V.; Belmonte, C.; Viana, F. TRPA1 channels: Novel targets of 1,4-dihydropyridines. Channels 2008, 2, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Tiruppathi, C.; Ahmmed, G.U.; Vogel, S.M.; Malik, A.B. Ca2+ signaling, TRP channels, and endothelial permeability. Microcirculation 2006, 13, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, G.U.; Malik, A.B. Functional role of TRPC channels in the regulation of endothelial permeability. Pflug. Arch. Eur. J. Physiol. 2005, 451, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G.; Voets, T.; Peters, J.A. Transient receptor potential cation channels in disease. Physiol. Rev. 2007, 87, 165–217. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Garland, C.J. Recent developments in vascular endothelial cell transient receptor potential channels. Circ. Res. 2005, 97, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Toth, B. I.; Olah, A.; Szollosi, A.G.; Biro, T. TRP channels in the skin. Br. J. Pharmacol. 2014, 171, 2568–2581. [Google Scholar] [CrossRef] [PubMed]
- Vandael, D.H.; Ottaviani, M.M.; Legros, C.; Lefort, C.; Guerineau, N.C.; Allio, A.; Carabelli, V.; Carbone, E. Reduced availability of voltage-gated sodium channels by depolarization or blockade by tetrodotoxin boosts burst firing and catecholamine release in mouse chromaffin cells. J. Physiol. 2015, 593, 905–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morley, N.; Curnow, A.; Salter, L.; Campbell, S.; Gould, D. N-acetyl-l-cysteine prevents DNA damage induced by UVA, UVB and visible radiation in human fibroblasts. J. Photochem. Photobiol. B Biol. 2003, 72, 55–60. [Google Scholar] [CrossRef]
- Oikawa, S. Sequence-specific DNA damage by reactive oxygen species: Implications for carcinogenesis and aging. Environ. Health Prev. med. 2005, 10, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.E.; Prahalad, A.K.; Williams, G.M. 8-Oxodeoxyguanosine formation in the DNA of cultured cells after exposure to H2O2 alone or with UVB or UVA irradiation. Photochem. Photobiol. 1996, 64, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Chen, B.H.; Wang, L.F.; Chen, J.S. Characterization of chondroitin sulfate and its interpenetrating polymer network hydrogels for sustained-drug release. Int. J. Pharm. 2007, 329, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Mi, F.L.; Liang, H.F.; Wu, Y.C.; Lin, Y.S.; Yang, T.F.; Sung, H.W. pH-sensitive behavior of two-component hydrogels composed of N,O-carboxymethyl chitosan and alginate. J. Biomater. Sci. Polym. Ed. 2005, 16, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Athar, M.; An, K.P.; Morel, K.D.; Kim, A.L.; Aszterbaum, M.; Longley, J.; Epstein, E.H., Jr.; Bickers, D.R. Ultraviolet B(UVB)-induced cox-2 expression in murine skin: An immunohistochemical study. Biochem. Biophys. Res. Commun. 2001, 280, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Chen, B.K.; Wang, Y.S.; Tsai, Y.T.; Chen, W.C.; Chang, W.C.; Hou, M.F.; Wu, Y.C.; Chang, W.C. Involvement of store-operated calcium signaling in EGF-mediated COX-2 gene activation in cancer cells. Cell. Signal. 2012, 24, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.T.; Yu, H.S.; Arbiser, J.L.; Hong, C.H.; Govindarajan, B.; Chai, C.Y.; Shan, W.J.; Lin, Y.F.; Chen, G.S.; Lee, C.H. Enhanced MCP-1 release by keloid CD14+ cells augments fibroblast proliferation: Role of MCP-1 and Akt pathway in keloids. Exp. Dermatol. 2010, 19, e142–e150. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.L.; Tsai, M.H.; Lin, M.W.; Chiu, Y.C.; Lu, J.H.; Chang, C.H.; Yu, H.S.; Yoshioka, T. Differential effects of arsenic on calcium signaling in primary keratinocytes and malignant (HSC-1) cells. Cell Calcium 2012, 52, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.L.; Chung, P.J.; Tsai, M.H.; Chang, C.L.; Liang, C.L. A role for Epstein-Barr viral BALF1 in facilitating tumor formation and metastasis potential. Virus Res. 2012, 163, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Seike, T.; Yutsudo, N.; Ohno, M.; Yamada, H.; Yamaguchi, H.; Sakumi, K.; Yamakawa, Y.; Kido, M.A.; Takaki, A.; et al. Hydrogen in drinking water reduces dopaminergic neuronal loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. PLoS ONE 2009, 4, e7247. [Google Scholar] [CrossRef] [PubMed]
- Cheon, G.J.; Cui, Y.; Yeon, D.S.; Kwon, S.C.; Park, B.G. Mechanisms of motility change on trinitrobenzenesulfonic Acid-induced colonic inflammation in mice. Korean J. Physiol. Pharmacol. 2012, 16, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compound Derinat is available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, W.-L.; Lu, J.-H.; Noda, M.; Wu, C.-Y.; Liu, J.-d.; Sakakibara, M.; Tsai, M.-H.; Yu, H.-S.; Lin, M.-W.; Huang, Y.-B.; et al. Derinat Protects Skin against Ultraviolet-B (UVB)-Induced Cellular Damage. Molecules 2015, 20, 20297-20311. https://doi.org/10.3390/molecules201119693
Hsu W-L, Lu J-H, Noda M, Wu C-Y, Liu J-d, Sakakibara M, Tsai M-H, Yu H-S, Lin M-W, Huang Y-B, et al. Derinat Protects Skin against Ultraviolet-B (UVB)-Induced Cellular Damage. Molecules. 2015; 20(11):20297-20311. https://doi.org/10.3390/molecules201119693
Chicago/Turabian StyleHsu, Wen-Li, Jian-He Lu, Mami Noda, Ching-Ying Wu, Jia-dai Liu, Manabu Sakakibara, Ming-Hsien Tsai, Hsin-Su Yu, Ming-Wei Lin, Yaw-Bin Huang, and et al. 2015. "Derinat Protects Skin against Ultraviolet-B (UVB)-Induced Cellular Damage" Molecules 20, no. 11: 20297-20311. https://doi.org/10.3390/molecules201119693