2.2. Argentatin B Induces Changes on Cell-Cycle Progression
We first determined the concentrations of argentatin B and cisplatin leading to 50% inhibition of cell proliferation (IC
50) for each cell line. Argentatin B IC
50 was 24 μM for HCT-15 and 34 μM for PC-3. Meanwhile, cisplatin IC
50 was determined to be 14 μM for both cell lines. To investigate whether argentatin B inhibits DNA replication we analyzed cell-cycle progression of cells treated with the determined argentatin B IC
50. As a control, cells treated with the cisplatin IC
50 were analyzed. As seen in
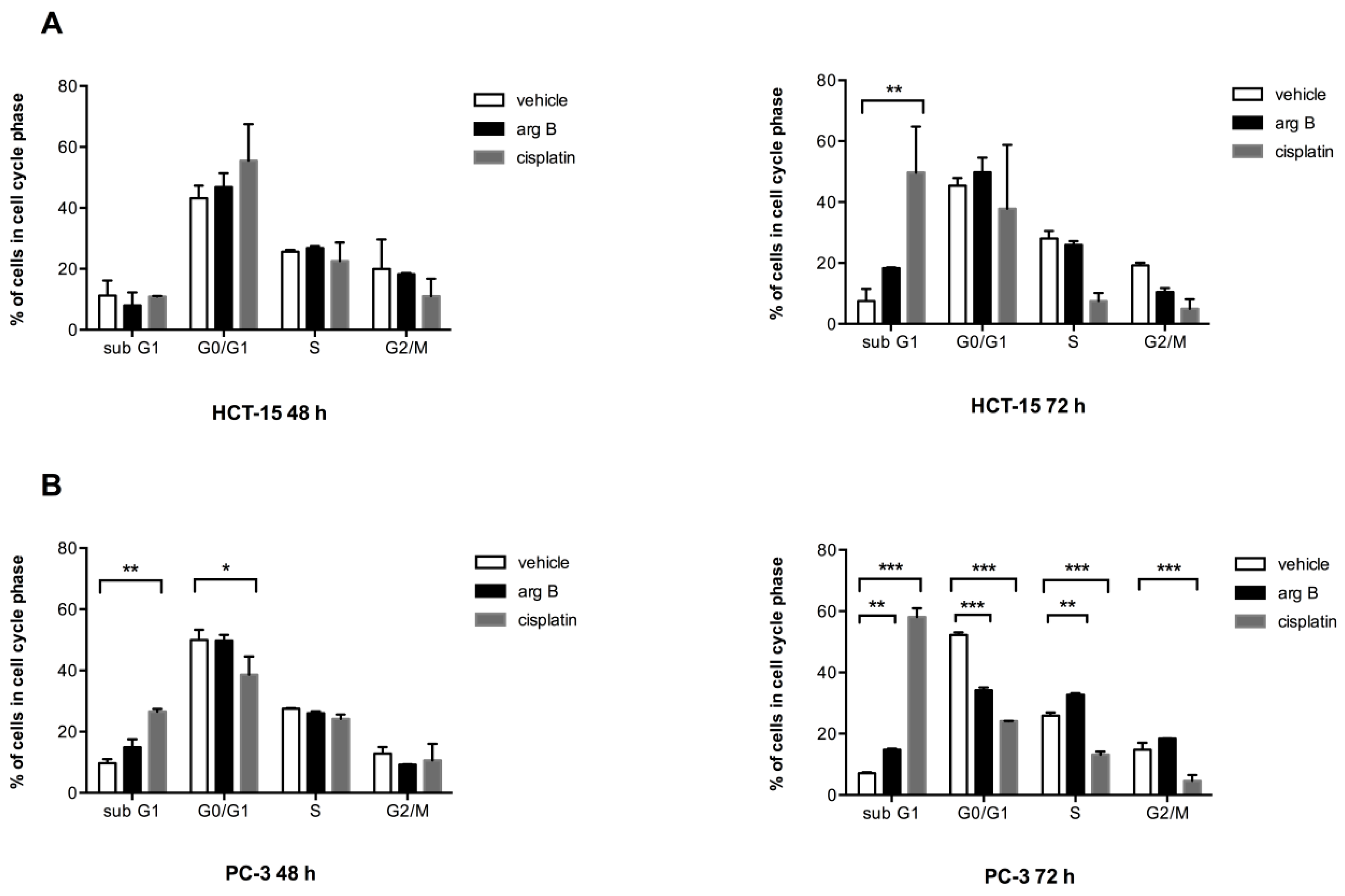
Figure 2A, no significant changes in cell cycle phases were observed in HCT-15 cells after 48 h of treatment. However, cells treated with argentatin B for 72 h showed a tendency to increase the proportion of cells in sub G1, along with a reduction of the number of cells in G2/M, but the proportion of cells in S phase remained unchanged. As expected, 72 h treatment with cisplatin produced a significant increase of the proportion of cells in sub G1 phase. It also reduced the proportion of cells in S and G2/M phases, although this reduction was not significant. On the other hand, changes in PC-3 cell cycle after 48 h were only observed when the cells were incubated with cisplatin (
Figure 2B). Nevertheless, treatment with argentatin B for 72 h induced a significant increment in the proportion of cells in sub G1, along with a reduction of cells in G0/G1. In addition an unexpectedly, marginally significant increase of cells in S phase was observed. Again, the number of cells in G2/M phase was not disturbed by argentatin B. In contrast, cisplatin produced a highly significant increment of cells in sub G1, along with the reduction of cells in G0/G1, S, and G2/M phases (
Figure 2B). These observations seem to suggest that argentatin B does not inhibit DNA replication, neither has a negative effect on cell mitosis.
Figure 2.
Effect of argentatin B on cell cycle progression. HCT-15 (A) and PC-3 (B) cells were incubated with 24 μM or 34 μM argentatin B, respectively, for 48 and 72 h. As a positive control cells were treated with cisplatin 14 μM. Negative control cultures received the carrier solvent (0.2% DMSO). The cells were stained with propidium iodide, and the cell cycle distribution was analyzed by flow cytometry using the BD CellQuest Pro Software. Data represent the average of three independent assays. Error bars indicate the standard error of the mean. * p < 0.05, ** p < 0.001, and *** p < 0.0001 vs. vehicle (one-way ANOVA test, and Tukey-Kramer post-test).
Figure 2.
Effect of argentatin B on cell cycle progression. HCT-15 (A) and PC-3 (B) cells were incubated with 24 μM or 34 μM argentatin B, respectively, for 48 and 72 h. As a positive control cells were treated with cisplatin 14 μM. Negative control cultures received the carrier solvent (0.2% DMSO). The cells were stained with propidium iodide, and the cell cycle distribution was analyzed by flow cytometry using the BD CellQuest Pro Software. Data represent the average of three independent assays. Error bars indicate the standard error of the mean. * p < 0.05, ** p < 0.001, and *** p < 0.0001 vs. vehicle (one-way ANOVA test, and Tukey-Kramer post-test).
2.3. Argentatin B Inhibits Cell Proliferation by Inducing Cell Senescence
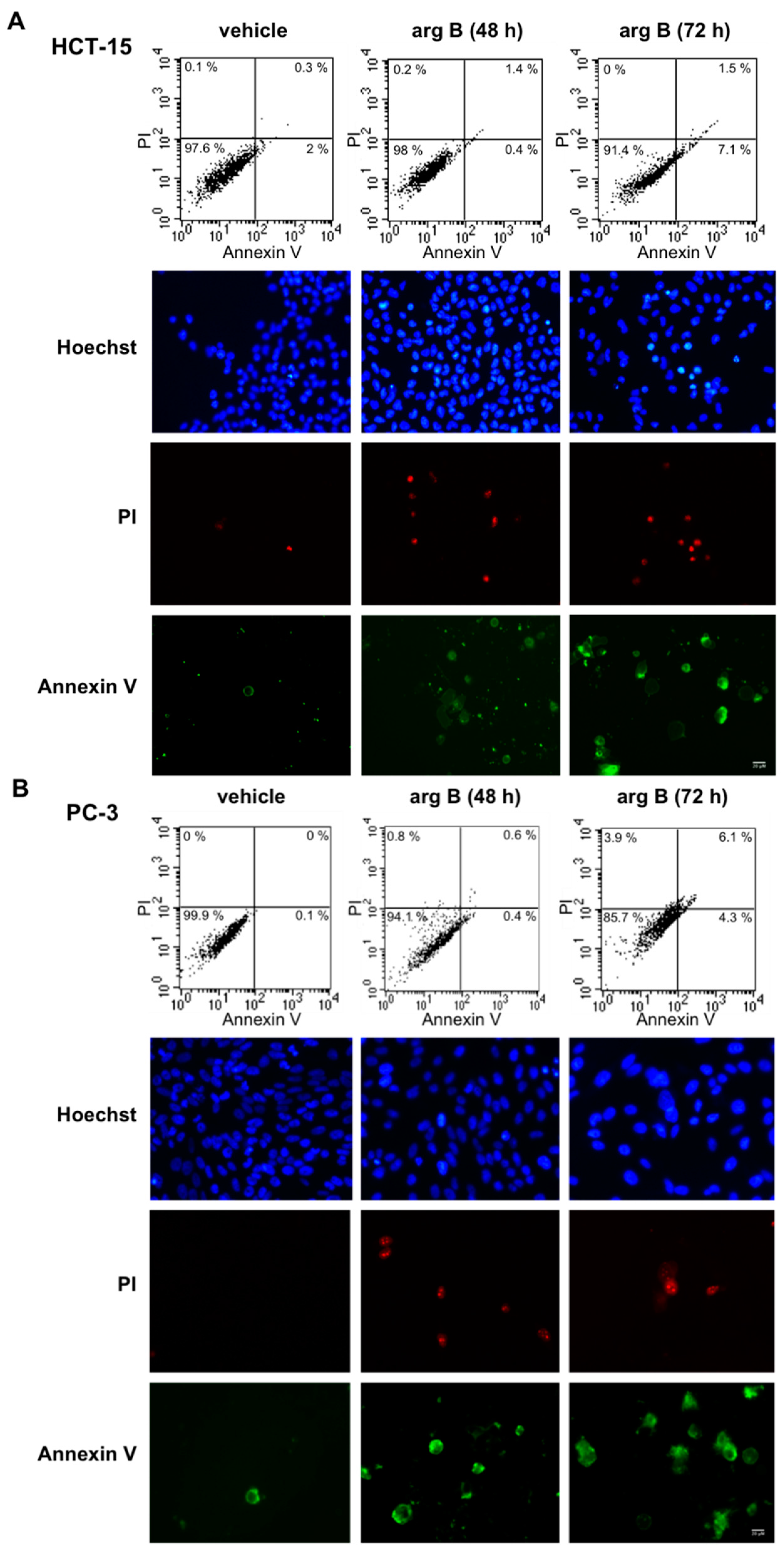
Since argentatin B induced an increase of cells in sub G1, we next investigated whether argentatin B can induce apoptotic cell death. After incubation of HCT-15 and PC-3 cells with argentatin B for 48 and 72 h, cell death was evaluated by staining with annexin V and propidium iodide. As shown in
Figure 3, argentatin B induced a modest increment of apoptotic (7.1%), and necrotic cells (1.5%) after 72 h incubation. Likewise, after 72 h incubation, a slight increment of apoptotic (4.3%), and necrotic (6.1%) PC-3 cells was observed (
Figure 3). These observations indicate that argentatin B is unable to induce a cytotoxic effect. However, we had previously demonstrated that argentatin B inhibits cell proliferation. Therefore, in an attempt to explain the observation mentioned above, we tested the cells for the presence of senescence. As seen in
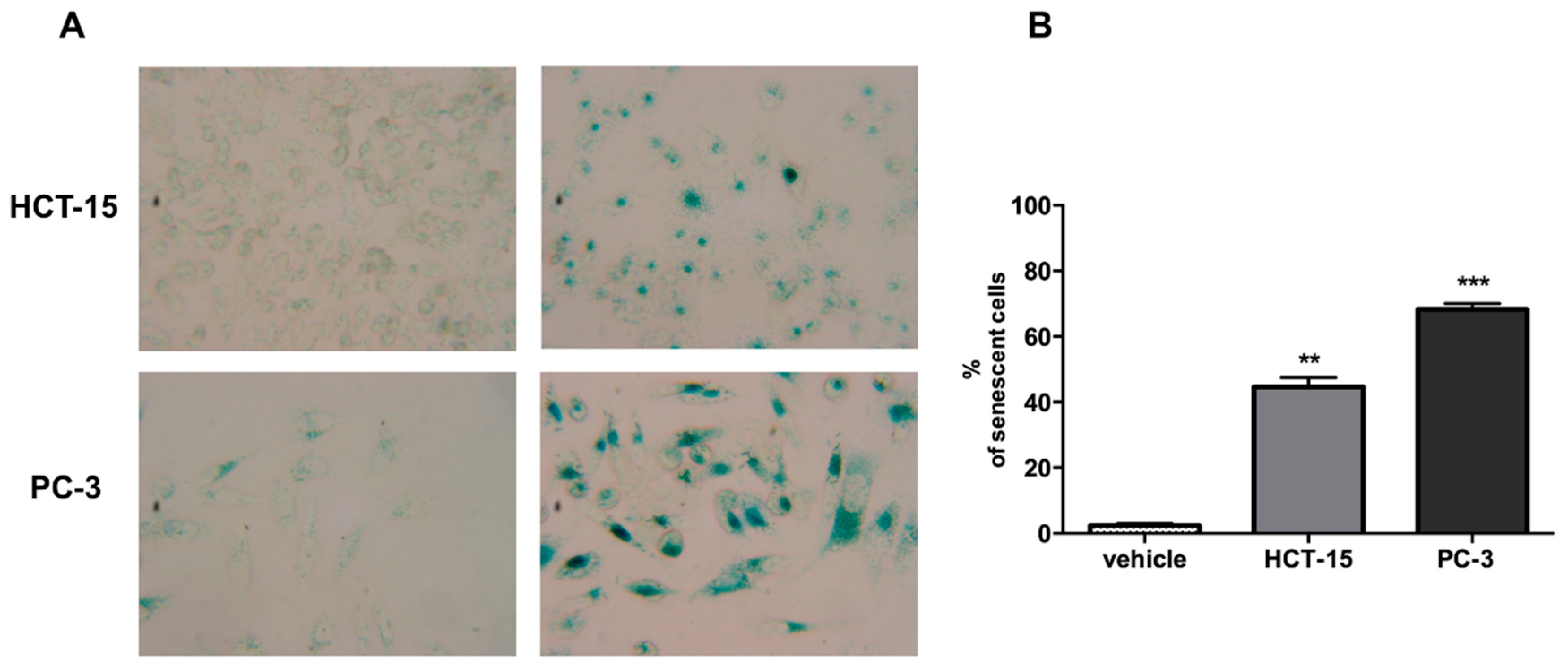
Figure 4A, after incubation with argentatin B for 72 h, both cell lines exhibited phenotypic changes that resemble those observed in cells undergoing senescence, such as flattened morphology and enlarged cell size. When tested for senescence associated-β-galactosidase activity, a proportion of 43% HCT-15, and 66% PC-3 cells showed a positive staining, compared with 2% of untreated controls. These findings suggest that argentatin B inhibits cell proliferation by inducing senescence.
Figure 3.
Effect of argentatin B on cell death. HCT-15 (A); and PC-3 (B) cells were incubated with argentatin B (arg B) for 48 h and 72 h. Cell death was analyzed by labelling with Annexin V and Propidum Iodide (PI). The number of apoptotic and necrotic cells was evaluated by flow cytometry (upper panel). The proportion of viable cells, showing negative annexin and PI staining is depicted in the left lower quadrant. Apoptotic cells, positive annexin, are shown in the right lower quadrant. Necrotic cells, positive annexin and PI staining, are presented in the right upper quadrant. Results are representative figures from three independent tests. Cells stained with Annexin, PI, and Hoechst were also analyzed by fluorescence microscopy (lower panel). Figures are representative micrographs from three independent experiments.
Figure 3.
Effect of argentatin B on cell death. HCT-15 (A); and PC-3 (B) cells were incubated with argentatin B (arg B) for 48 h and 72 h. Cell death was analyzed by labelling with Annexin V and Propidum Iodide (PI). The number of apoptotic and necrotic cells was evaluated by flow cytometry (upper panel). The proportion of viable cells, showing negative annexin and PI staining is depicted in the left lower quadrant. Apoptotic cells, positive annexin, are shown in the right lower quadrant. Necrotic cells, positive annexin and PI staining, are presented in the right upper quadrant. Results are representative figures from three independent tests. Cells stained with Annexin, PI, and Hoechst were also analyzed by fluorescence microscopy (lower panel). Figures are representative micrographs from three independent experiments.
Figure 4.
Argentatin B induces cell senescence at 72 h. (A) Representative micrographs of HCT-15 and PC-3 treated with argentatin B or vehicle (Magnification, ×40); (B) SA-β-gal-positive cells were evaluated by counting more than 100 cells for each treatment. Values presented are the mean of three independent experiments. Error bars indicate the standard error of the mean. ** p < 0.001, and *** p < 0.0001 vs. vehicle (one-way ANOVA test, and Tukey-Kramer post-test)
Figure 4.
Argentatin B induces cell senescence at 72 h. (A) Representative micrographs of HCT-15 and PC-3 treated with argentatin B or vehicle (Magnification, ×40); (B) SA-β-gal-positive cells were evaluated by counting more than 100 cells for each treatment. Values presented are the mean of three independent experiments. Error bars indicate the standard error of the mean. ** p < 0.001, and *** p < 0.0001 vs. vehicle (one-way ANOVA test, and Tukey-Kramer post-test)
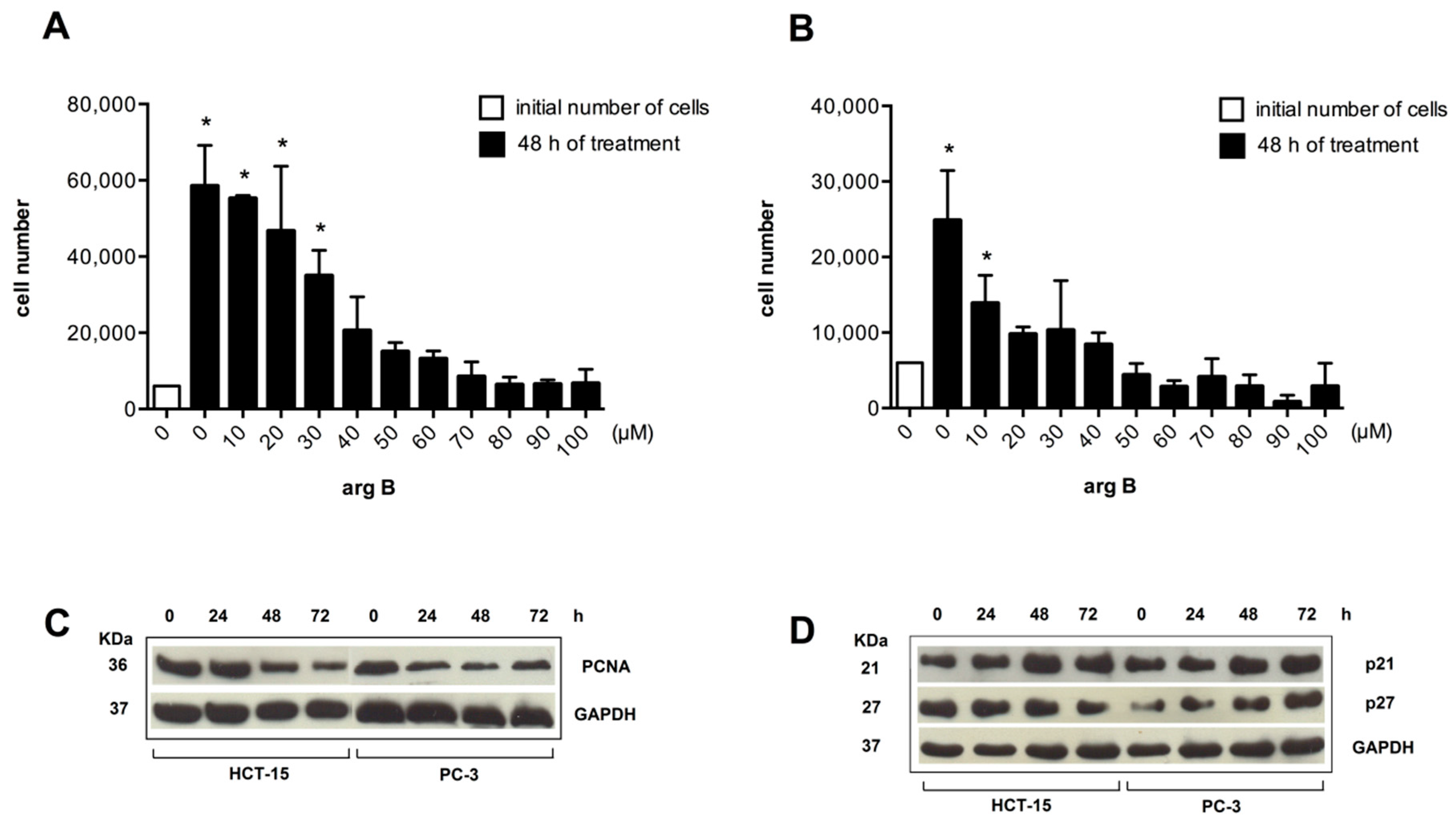
Figure 5.
Argentatin B inhibits cell proliferation and increases expression of senescence-associated proteins. HCT-15 (A), and PC-3 (B) cells were treated with the indicated concentrations of argentatin B for 48 h. Cell numbers were evaluated by using the colorimetric MTT assay. Values presented are the mean of three independent experiments. Error bars indicate the standard error of the mean. * p < 0.05, vs. the number of cells seeded at the beginning of the experiment (initial number of cells) (one-way ANOVA test, and Tukey-Kramer post-test); (C) Western blot analysis of the expression of the proliferation marker PCNA. HCT-15, and PC-3 were incubated with 24 μM or 34 μM argentatin B, respectively for 24, 48, and 72 h. Detection of GAPDH was included as an internal control; (D) Western blot analysis of the expression of negative cell cycle regulators p21 and p27 in response to argentatin. HCT-15 and PC-3 cells were incubated with 24 μM or 34 μM argentatin B, respectively for 24, 48, and 72 h. Detection of GAPDH was included as an internal control.
Figure 5.
Argentatin B inhibits cell proliferation and increases expression of senescence-associated proteins. HCT-15 (A), and PC-3 (B) cells were treated with the indicated concentrations of argentatin B for 48 h. Cell numbers were evaluated by using the colorimetric MTT assay. Values presented are the mean of three independent experiments. Error bars indicate the standard error of the mean. * p < 0.05, vs. the number of cells seeded at the beginning of the experiment (initial number of cells) (one-way ANOVA test, and Tukey-Kramer post-test); (C) Western blot analysis of the expression of the proliferation marker PCNA. HCT-15, and PC-3 were incubated with 24 μM or 34 μM argentatin B, respectively for 24, 48, and 72 h. Detection of GAPDH was included as an internal control; (D) Western blot analysis of the expression of negative cell cycle regulators p21 and p27 in response to argentatin. HCT-15 and PC-3 cells were incubated with 24 μM or 34 μM argentatin B, respectively for 24, 48, and 72 h. Detection of GAPDH was included as an internal control.
![Molecules 20 19757 g005]()
It is known that the main characteristic of senescent cells is the inhibition of proliferation. PCNA expression is a hallmark of cell division. Thus, we analyzed the effect of increasing concentrations of argentatin B on cell proliferation, and its effect on the expression of PCNA. As shown in
Figure 5, argentatin B induced a reduction of cell proliferation in a dose-dependent manner in both, HCT-15 (
Figure 5A), and PC-3 (
Figure 5B) cells. A significant reduction in the number of cells was observed from 30 to 100 μM of argentatin B for HCT-15, and from 10 to 100 μM of argentatin B for PC-3 cells. However, they were never significantly smaller than the initial number of cells seeded, supporting the observation that argentatin B induces the inhibition of proliferation rather than cell death. Accordingly, Western blot analysis of PCNA showed that the expression of this cell division-associated protein decreased when the cells were incubated with argentatin B (
Figure 5C). It is accepted that senescence is promoted by a number of anti-proliferative mechanisms. Classical negative cell cycle regulators, such as p21, and p27, have been associated with the senescent phenotype. Therefore, we next analyzed the expression of p21 and p27 in cell cultures treated with argentatin B. HCT-15 and PC-3 cells were incubated with 24 μM or 34 μM argentatin B, respectively. The expression of p21 and p27 was evaluated at 24, 48, and 72 h by Western blotting. As seen in
Figure 5D, treatment with argentatin B induced an increment of both, p21 and p27 in PC-3 cells after 48 h incubation, and it was persistent for at least 72 h after treatment. In the case of HCT-15, an increment of p21 expression was observed after 48 h treatment and it was persistent until 72 h of argentatin B treatment. In contrast, the level of p27 was not changed in HCT-15 after 24–48 h argentatin B treatment, and a reduction of p27 expression was observed at 72 h of treatment. Taken together, these results indicate that treatment with argentatin B induces the cells to undergo senescence.
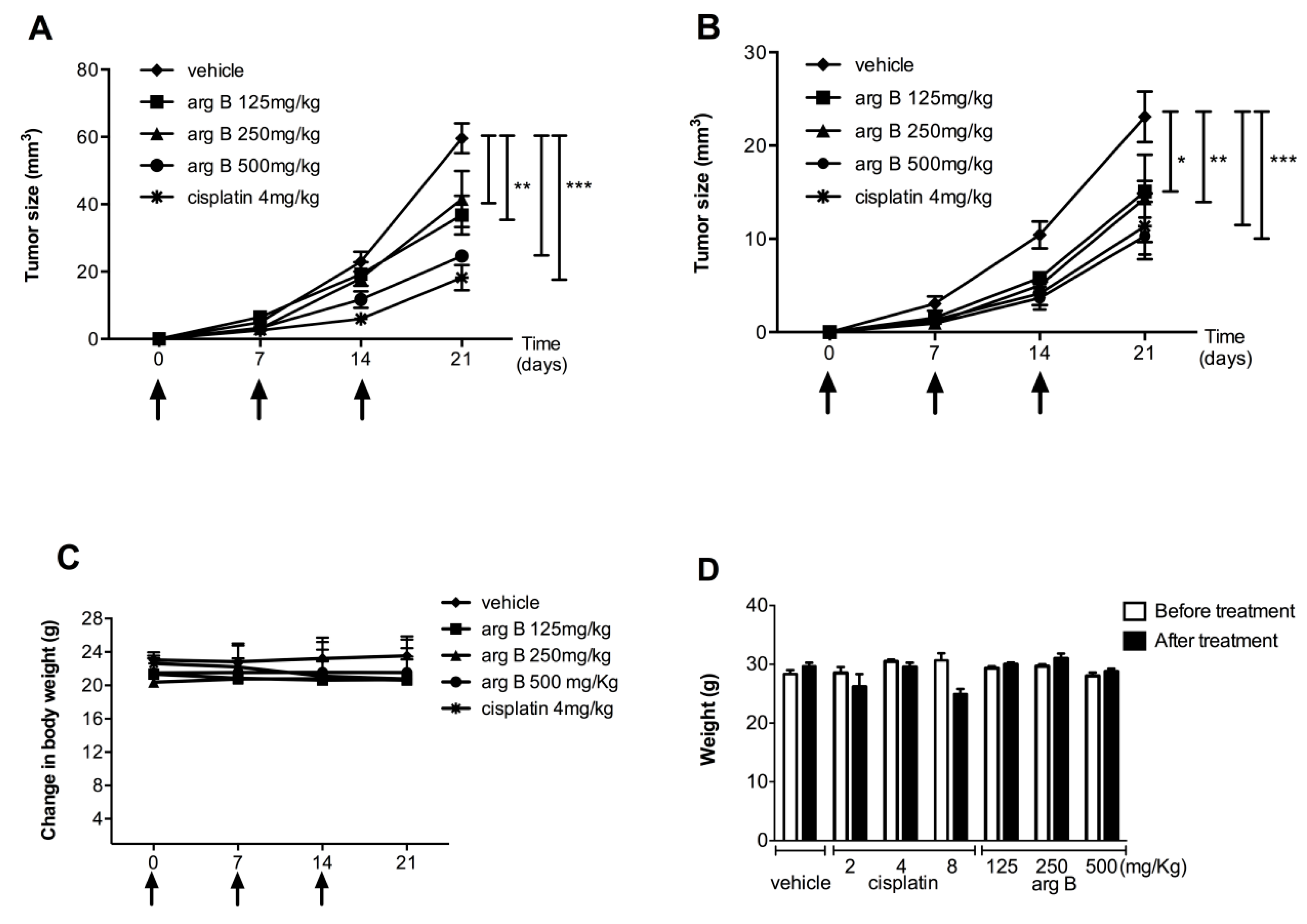
2.4. Argentatin B Reduces Tumor Growth in Vivo
To further examine the antiproliferative effect of argentatin B,
in vivo xenografts using HCT-15 and PC-3 cells were established in mice. Animals were treated with three different concentrations of argentatin B or vehicle (sesame oil). Treatment with cisplatin was used as a positive control. As shown in
Figure 6, the growth of HCT-15 tumors was significantly reduced after treatment with 125 mg/kg (
p < 0.001), 250 mg/kg (
p < 0.001), and 500 mg/kg argentatin B (
p < 0.0001), by day 21 (
Figure 6A). As expected, growing of HCT-15 tumors was also inhibited by cisplatin (
Figure 6A). As seen in
Figure 2B, growing of PC-3 tumors was significantly reduced by treatment with cisplatin (
p < 0.0001), 500 mg/kg (
p < 0.0001), 250 mg/kg (
p < 0.001), and 125 mg/kg argentatin B (
p < 0.05) at the end of the experiment (day 21). The toxicity of treatments is shown in the
Figure 6C, no change in weight was observed, indicating no systemic toxicity with any of the treatments. Our results suggest that argentatin B restricts HCT-15 and PC-3 cells proliferation
in vivo as efficiently as cisplatin. We next examined the potential systemic toxicity of argentatin B in healthy mice. All animals survived the treatment with argentatin B. No significant differences in general appearance, depression of activity, respiratory difficulty, abnormal aggressive behavior, and in mean daily food and water consumption between the experimental groups and the untreated controls were observed. Histopathological analysis showed no evidence of treatment-related pathology. Besides, no significant differences in body weight after and before treatment were observed (
Figure 6D), suggesting that argentatin B is not toxic at the concentrations tested.
2.5. Discussion
Here we have presented evidence that argentatin B inhibits proliferation of HCT-15 and PC-3 cells by inducing senescence. The cytotoxic effect of some cycloartane-type triterpenoids has been associated with a strong pro-apoptotic activity [
16]. Some cycloartane-type triterpenoids have been isolated and tested against cancer cells. Their cytotoxic effect has been reported to be mediated by some potential mechanisms. There is evidence suggesting that triterpenoids inhibit cell proliferation by interfering with DNA replication [
5,
6,
7]. Interestingly, a cycloartane-type triterpenoid isolated from
Commiphora opobalsamum showed a moderate antiproliferative effect on human prostate cancer cells. However, it was able to inhibit the expression of androgen receptors in the cells [
17], suggesting that triterpenoids may modulate some mechanisms involved in the regulation of cell proliferation. Tian
et al. [
18] reported that schisandrolic and isoschisandrolic acids exerted their cytotoxic effect via G0/G1 arrest and subsequent apoptosis. There is evidence showing that cycloartane-type triterpenoids may induce both, cytostatic and cytotoxic effects, increasing the number of cells in sub G1, and arresting cells in S and G2/M phases [
19]. Accordingly, we observed that treatment with argentatin B induced a significant increment in the proportion of cells in sub G1. Accumulation of cells in sub G1, has also been documented to occur as a cellular response to powerful reactive oxygen species, causing oxidative stress that leads the cell to develop a condition of senescence [
20].
Figure 6.
Evaluation of effects of argentatin B in vivo. Antitumor activity of argentatin B was evaluated in nu/nu mice implanted with HCT-15 (A) and PC3 (B) cells. Mice received the indicated concentrations of argentatin B, cisplatin or sesame oil (vehicle) at days 0, 7, and 14. Each point represents the average ± SEM of six animals. Significant differences of * p < 0.05, ** p < 0.001, and *** p < 0.0001 vs. vehicle are shown (one-way ANOVA test, and Tukey-Kramer post-test); (C) Evaluation of body weight change in nu/nu mice; (D) The toxicity of argentatin B was tested in groups of three CD-1 mice. The indicated doses of argentatin B, cisplatin or vehicle were administered intraperitoneally every week. The weight of the mice was recorded every other day. Error bars indicate the standard error of the mean.
Figure 6.
Evaluation of effects of argentatin B in vivo. Antitumor activity of argentatin B was evaluated in nu/nu mice implanted with HCT-15 (A) and PC3 (B) cells. Mice received the indicated concentrations of argentatin B, cisplatin or sesame oil (vehicle) at days 0, 7, and 14. Each point represents the average ± SEM of six animals. Significant differences of * p < 0.05, ** p < 0.001, and *** p < 0.0001 vs. vehicle are shown (one-way ANOVA test, and Tukey-Kramer post-test); (C) Evaluation of body weight change in nu/nu mice; (D) The toxicity of argentatin B was tested in groups of three CD-1 mice. The indicated doses of argentatin B, cisplatin or vehicle were administered intraperitoneally every week. The weight of the mice was recorded every other day. Error bars indicate the standard error of the mean.
It has been shown that tumor growth-inhibitory effects exhibited by natural compounds, such as resveratrol, can be attributed to the induction of cell senescence [
21,
22]. Here we observed that cells treated with argentatin B presented phenotypic changes that resembled those detected in cells undergoing senescence, such as flattened morphology and enlarged cell size. In fact, only after 48 h these changes are evident and at 72 h treatment 66% PC-3, and 43% HCT-15 cells were positive for senescence associated-β-galactosidase activity. Cellular senescence is a complex, multifactorial condition of arrested growth. Senescence-associated signaling pathways activate important cell cycle negative regulators, like p21 which is a strong inhibitor of cyclin-dependent kinases [
23], and p27 which is a negative regulator of G1 progression [
24]. Accordingly, the expression of p27 was elevated in PC-3 cells as a response to argentatin B treatment. However, the level of p27 in HCT-15 remained unchanged during the first 48 h of treatment, and decreased at 72 h. In contrast, a clear elevation of p21 levels was detected in both PC-3, and HCT-15 cells. In accordance with our results, previous reports have demonstrated that inhibitors of proliferation, such as adamantane derivatives [
25], and celecoxib [
26], consistently induce an increment of p21 expression in different colon cancer-derived cell lines. Nevertheless, adamadate derivative DPA produced no change, or even a reduction of p27 in colon cancer-derived Colo 205 and HT-29 cell lines [
25], while celecoxib induced a decrease of p27 levels at short times incubation (0–5 to 8 h), followed by an increasing expression of p27 at 24 h in HCT-15, HCT-29, and Caco-2 cell lines [
26].
Our results showed that argentatin B inhibited cell proliferation, decreased the expression of PCNA (proliferation-related marker), and also induced an increment of p21 expression in HCT-15 and PC-3 cells. It will very interesting to determine if argentatin B affects other proteins involved with cellular senescence such as p16, p53 or proinflammatory molecules.
Since this protein has been associated with a full senescence state, these observations provide evidence to support the fact that argentatin B induces senescence. Cellular senescence is currently considered an important target for cancer treatment, inasmuch as it leads to an irreversible obstruction of the cellular division capacity. Moreover, tumor cells activate senescence mechanisms as a response to chemotherapy and radiotherapy [
27].
Cellular senescence induced by stress is known as premature senescence [
27]. It occurs within a week of exposure to sub lethal stresses. Senescent PC-3 and HCT-15 cells were observed three days after argentatin B treatment, suggesting the induction of premature senescence. There is evidence showing that other triterpenoids can induce senescence. For instance, Chang
et al. [
28] reported that ganoderiol F, a tetracyclic triterpene isolated from
Ganoderma amboinense, induces senescence after 18 days of continuous treatment of HepG2. Similarly, administration of 20–50 μM resveratrol has been reported to produce senescence in lung cancer cells after 10 to 12 days incubation [
21]. In sharp contrast, concentrations of 24 and 34 μM argentatin B can induce senescence in confluent cell cultures in only 48–72 h. Interestingly, commonly used chemotherapeutic drugs, such as cisplatin, doxorubicin, etoposide and other topoisomerase inhibitors are capable of inducing senescence of cancer cells when they used in very low concentrations [
29,
30].
In the present study we observed that argentatin B can inhibit the growth of human colon carcinoma (HCT-15, IC50 24.14 ± 5.58) and human prostate cancer (PC-3, IC50 34.14 ± 3.71 μM) cell lines. We know that triterpenes such ursolic acid and acetyl-boswellic acid with low cytotoxic activities showed an effective antitumor effect in mice xenograft model. With this in mind, we decide to evaluate the antitumor activity of argentatin B in a xenograft mice model using PC-3 and HCT-15 human cell lines.
Our results suggest that argentatin B restricts HCT-15 and PC-3 cells proliferation
in vivo as efficiently as cisplatin. Frequently,
in vivo testing of new antitumor agents is carried out using daily administrations of the drug for 15-days periods. We also demonstrated that three weekly administrations of argentatin-B were enough to induce a significant decrease of tumor growth, and produced no toxic effects on the experimental animals. It has become clear that tumor cells can undergo senescence in response to chemotherapy. Mitotic arrest is normally not tolerated by cells, and it is resolved by cell death. Thus, cytostatic activity can be followed by cell death. In fact, many cytotoxic agents are primarily cytostatic [
31]. Thus, stimulation of this response is nowadays considered a rational approach to cancer treatment. In this work, we have shown that argentatin B can inhibit cell proliferation by inducing cellular senescence. However, the demonstration of cytostasis or cytotoxicity depends on experimental conditions, so it would be important to test argentatin B under a wide range of doses and times schedules, both
in vitro and
in vivo to evaluate its potential pharmaceutical usefulness.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





