
Indole Alkaloids from Catharanthus roseus: Bioproduction and Their Effect on Human Health


Abstract
:1. Introduction
2. Pharmacological Activities
2.2. Antidiabetic and Antioxidant Properties
2.3. Potential Effects of Yohimbine on Erectile Dysfunction
2.4. Potential Effects of Ajmalicine and Serpentine as Anti-Neuro-Inflammatory Agents
2.5. Potential Effects of Ajmalicine on Vascular Disorders
2.6. Antitumour Properties
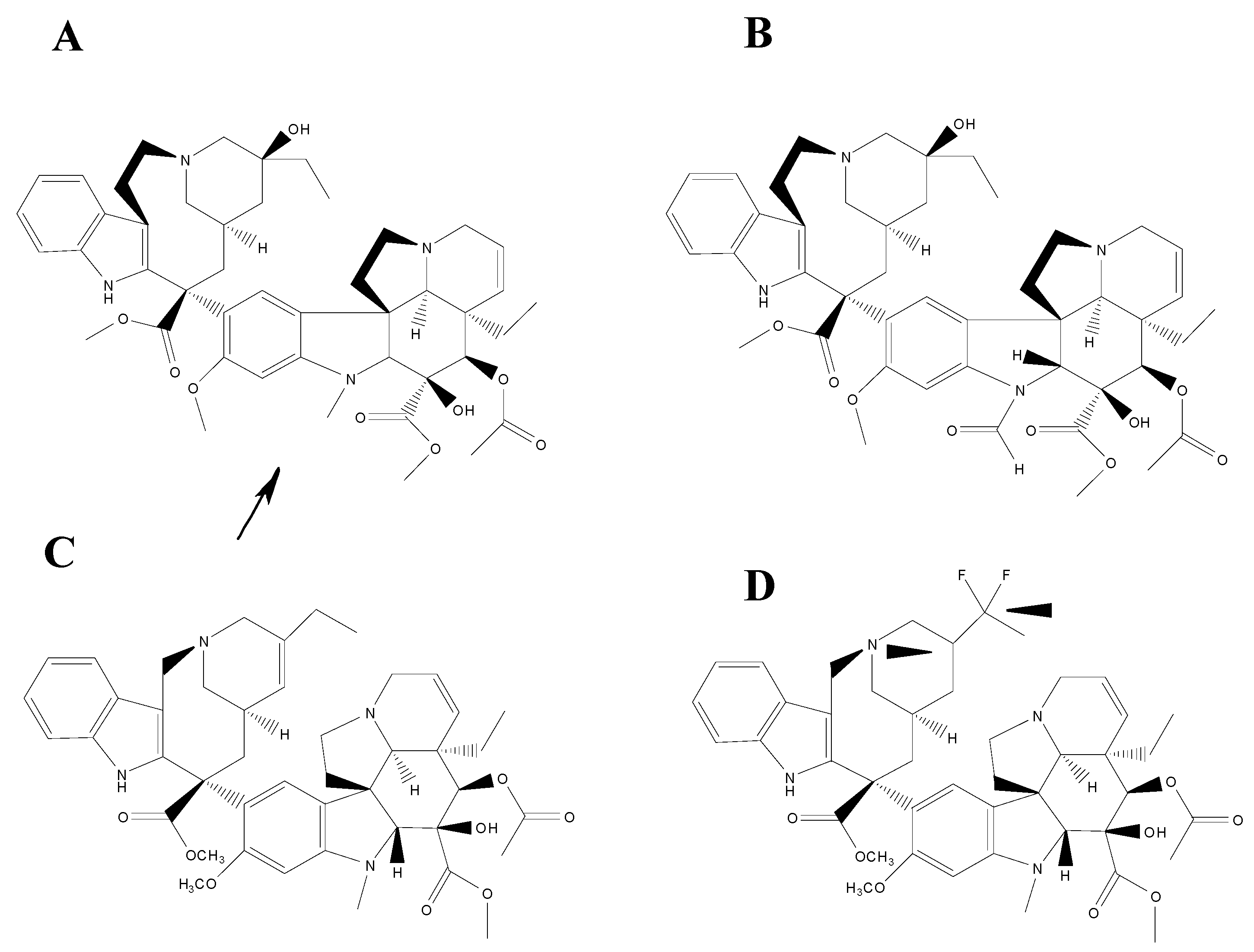
2.6.1. Semi-Synthetic Derivatives of Dimeric Alkaloids
2.6.2. Mechanism of Action of Dimeric Alkaloids

3. Keynotes Based on the Use of C. roseus Plants as Biofactories for Enhancing the Production of Terpenoid Indole Alkaloids
4. Empirical Strategies for Improving TIA Production
4.1. Empirical Optimization of the Culture Medium and Culture Conditions

{kind=link}
{kind=link}
| Material | Strategy | Total TIAs | Observations | Refs |
|---|---|---|---|---|
| Compact callus cluster | Medium supplemented with mannitol (250 mM) | 4.13-fold increase (75 mg/L) | 5.05-fold increase in catharanthine (19.7 mg/L) | [65] |
| 2.77-fold increase in serpentine (13.6 mg/L) | ||||
| 4.54-fold increase in ajmalicine (42.3 mg/L) | ||||
| Medium supplemented with KCl (4g/L) | 3.15-fold increase (57.8 mg/L) | 3.15-fold increase in catharanthine (12.3 mg/L) | ||
| 2.42-fold increase in serpentine (11.9 mg/L) | ||||
| 3.62-fold increase in ajmalicine (33.5 mg/L) | ||||
| Compact callus cluster | Medium supplemented with sucrose (50 g/L) | 1.16-fold increase (46.7 mg/L) | 1.25-fold increase in serpentine (10 mg/L) | [68] |
| 1.20-fold increase in ajmalicine (14.5 mg/L) | ||||
| Suspension cultured cells | High cell density (200 g FW/L) | ND | 120-fold increase in ajmalicine (60 mg/L) | [73] |
| Immobilized cells | High cell density (100 g FW/L) | ND | 2-fold increase in ajmalicine (120 mg/L) | [71] |
| Immobilized cells | Elicitation with Phytophthora cactorum | ND | 45-fold increase in ajmalicine (90 mg/L) | [72] |
| Shoot culture | MS medium supplemented with plant growth regulators | ND | The concentrations of 8.90 µM BA and 2.85 µM IAA increased the production of ajmalicine (0.85 g/L) | [74] |
| Callus | MS medium supplemented with plant growth regulators | ND | The concentrations of 2.21 µM BA and 5.7 µM IAA increased the production of catharanthine (0.12 mg/g DW) | [68] |
| ajmalicine (0.35 mg/g DW) | ||||
| vindoline (0.19 mg/g DW) | ||||
| serpentine (0.53 mg/g DW) | ||||
| Immobilized cells | Variation of O2 and CO2 concentration | ND | 1.1-fold increase in ajmalicine (275 g/L ) | [75] |
| Suspension cultured cells | Feeding with loganin and triptamine | ND | 17.73-fold increase in strictosidine (53.19 µmo/g DW) | [76] |
| 6.4-fold increase in ajmalicine (3.2 µmol/g DW) | ||||
| Compact callus cluster | Feeding with succinic acid (10 mM) | 4.86-fold increase (73 mg/L) | 3.5-fold increase in catharanthine (7 mg/L) | [65] |
| 7.5-fold increase in serpentine (15 mg/L) | ||||
| 16-fold increase in ajmalicine (32 mg/L) | ||||
| Feeding with tryptamine (3.12 mM) | 3.86-fold increase (58 mg/L) | 2.5-fold increase in catharanthine (5 mg/L) | ||
| 7-fold increase in serpentine (14 mg/L) | ||||
| 15.5-fold increase in ajmalicine (31 mg/L) | ||||
| Feeding with tryptophan (2.44 mM) | 5.53-fold increase (68 mg/L) | 2.5-fold increase in catharanthine (5 mg/L) | ||
| 6-fold increase in serpentine (12 mg/L) | ||||
| 14-fold increase in ajmalicine (28 mg/L) | ||||
| Hairy root culture | Feeding with geraniol (0.5 mM) | ND | 1.5-fold increase in tabersonine (1.4 mg/g DW) | [77] |
| Hairy root culture | Elicitation with sodium nitroprusside (0.1 mM) | 1.42-fold increase (3.7 mg/g DW) | 2-fold increase in lochnericine (1 mg/g DW) | [78] |
| 2.3-fold increase in tabersonine (0.7 mg/g DW) | ||||
| 2-fold increase in ajmalicine (0.7 mg/g DW) | ||||
| Suspension cultured cells | Elicitation with MeJA (101.9 µM) | 1.33-fold increase (2.2 mg/g DW) | 2-fold increase in tabersonine (3.8 mg/g DW) | [79] |
| Suspension cultured cells | Elicitation with MeJA (100 µM) | ND | 27.44-fold increase in ajmalicine (137.2 mg/L) | [80] |
| 11.12-fold increase in catharanthine (55.6 mg/L) | ||||
| Hairy root culture | Elicitation with MeJA (250 µM) | 1.32-fold increase (49 mg/L) | 7-fold increase in ajmalicine (6.34 mg/g DW) | [81] |
| 2.9-fold increase in serpentine (1.71 mg/g DW) | ||||
| 3-fold increase in ajmaline (12 mg/g DW) | ||||
| 3-fold increase in catharanthine (4.34 mg/g DW) | ||||
| Suspension cultured cells | Elicitation with Trichoderma viride | ND | 7.9-fold increase in ajmalicine (0.166 mg/g DW) | [82] |
| Suspension cultured cells | Elicitation with the protein of Phytophthora boehmeriae (BP90) | ND | 4-fold increase in catharanthine (20 mg/L) | [83] |
| Suspension cultured cells | Elicitation with CDs | ND | 40-fold increase in ajmalicine (200 mg/L) | [80] |
| 17-fold increase in catharanthine (85 mg/L) | ||||
| Suspension cultured cells | Elicitation with UV-B light | ND | 3-fold increase in catharanthine (0.12 mg/g DW) | [84] |
| 117.6-fold increase in vindoline (0.06 mg/g DW) | ||||
| Suspension cultured cells | Elicitation with UV-C light | ND | 18-fold increase in ajmalicine (90 mg/L) | [73] |
| 10-fold increase in catharanthine (50 mg/L) | ||||
| Plant | Elicitation with chromium (50 µM) | ND | 1.5-fold increase in vincristine (2 µg/g DW) | [35] |
| 2.16-fold increase in vinblastine (2.25 µg/g DW) | ||||
| Suspension cultured cells | Elicitation with Aspergillum niger mycelium and tetramethyl ammonium bromide | 3.84-fold increase (96 mg/L) | 21-fold increase in ajmalicine (63 mg/L) | [85] |
| 17-fold increase in catharanthine (17 mg/L) | ||||
| Elicitation with malate and sodium alginate | 3.28-fold increase (82 mg/L) | 13.6-fold increase in ajmalicine (41 mg/L) | ||
| 26-fold increase in catharanthine (26 mg/L) | ||||
| Suspension cultured cells | Elicitation with MeJA and CDs | ND | 90-fold increase in ajmalicine (450 mg/L) | [73] |
| 31-fold increase in catharanthine (155 mg/L) | ||||
| Elicitation with MeJA, CDs and UV-C light | ND | 2.3-fold increase in ajmalicine (1040 mg/L) (85 mg/g DW) | ||
| 1.26-fold increase in catharanthine (196 mg/L) (10 mg/g DW) | ||||
| Suspension cultured cells | Overexpression of STR | 24.6 fold in increase (123 mg/L) | _ | [86] |
| Suspension cultured cells | Overexpression of TDC and feeding with loganin and secologanin | 125 fold in increase (625 mg/L) | _ | [87] |
| Hairy root culture | Overexpression of DAT | ND | 4-fold increase in hörhammericine (0.16 mg/g DW) | [88] |
| Hairy root culture | Overexpression of CrPrx | 1.5-fold increase (85 mg/g DW) | 5-fold increase in serpentine (3.7 mg/g DW) | [89] |
| 3-fold increase in ajmalicine (0.35 mg/g DW) | ||||
| Hairy root culture | Overexpression of DXS | NV | 1.66-fold increase in ajmalicine (1.5 mg/g DW) | [90] |
| 1.66-fold increase in lochnericine (1 mg/g DW) | ||||
| Overexpression of ASα | NV | 1.25-fold increase in lochnericine (2.5 mg/g DW) | ||
| Overexpression of G10H/DXS | 0.0072 mg/g DW | 1.35-fold increase in tabersonine (0.9 mg/g DW) | ||
| 1.15-fold increase in lochnericine (1.4 mg/g DW) | ||||
| Overexpression of ASα/DXS | 0.015 mg/g DW | 1.16-fold increase in tabersonine (1.7 mg/g DW) | ||
| 1.18-fold increase in lochnericine (2 mg/g DW) | ||||
| Leaves | Transient overexpression of GPPS | ND | 1.6-fold increase in vindoline (2.5 mg/g DW) | [91] |
| Tobacco cell cultures | Overexpression of TDC/STR | ND | Enhancement in strictosidine (5.3 mg/L) | [92] |
| Morinda citrifolia cell cultures | Overexpression of TDC/STR | ND | Enhancement in strictosidine (21.2 mg/L) | |
| Cinchona officinalis hairy root culture | Overexpression of TDC/STR | ND | Enhancement in strictosidine (1.95 mg/g FW) | [93] |
| Saccharomyces cerevisiae | Overexpression of STR/SGD | ND | Enhancement in strictosidine (2000 mg/L) | [94] |
| Hairy root culture | Overexpression of transcription factor CrWRKY1 | ND | 3-fold increase in serpentine (0.291 mg/g DW) | [95] |
| 10-fold increase in ajmalicine (0.015 mg/g DW) | ||||
| Leaves | Transient overexpression of transcription factor CrMPK3 | ND | 3.52-fold increase in serpentine (0.061 mg/g DW) | [96] |
| 2.66-fold increase in vindoline (4.1 mg/g DW) | ||||
| 1.44-fold increase in catharanthine (1.3 mg/g DW) | ||||
| 2-fold increase in vincristine (1.75 mg/g DW) | ||||
| Hairy root culture | Overexpression of transcription factor ORCA3 | ND | 2.5-fold increase in catharanthine (5.6 mg/g DW) | [97] |
| Hairy root culture | Overexpression of transcription factor ORCA2 | ND | 2-fold increase in catharanthine (4.8 mg/g DW) | [98] |
| Transgenic plant | Overexpression of ORCA3 and G10H | ND | 3.03-fold increase in vindoline (2.1 mg/g DW) | [99] |
| 2.29-fold increase in catharanthine (4.6 mg/g DW) | ||||
| 6.30-fold increase in ajmalicine (0.315 mg/g DW) | ||||
| 1.08-fold increase in anhydrovinblastine (10.2 mg/g DW) | ||||
| 10.2-fold increase in vinblastine (0.27 mg/g DW) |
4.2. Feeding with Precursors and Elicitation as Empirical Strategies for Increasing the Production of TIAs
5. Rational Approaches to the Biotechnological Production of TIAs

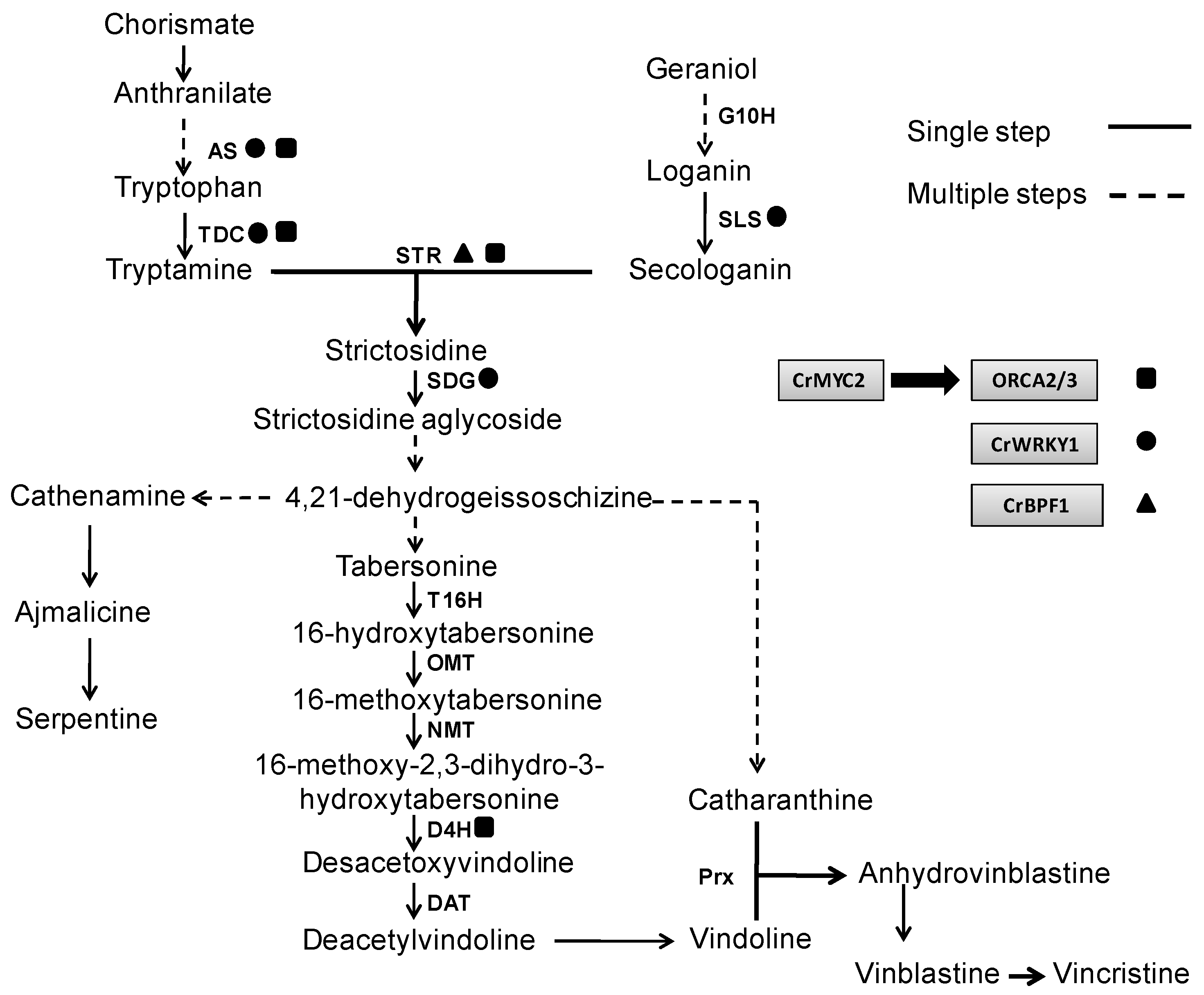
5.1. Elicitor Effects on the Expression of Genes Involved in TIA Metabolism
5.2. Metabolic Engineering to Improve the Production of TIAs
5.2.1. Overexpression of Genes which Regulate TIA Biosynthesis
5.2.2. Overexpression of Transcription Factors which Regulate TIA Biosynthesis
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van der Heijden, R.; Jacobs, D.I.; Snoeijer, W.; Hallard, D.; Verpoorte, R. The Catharanthus alkaloids: Pharmacognosy and biotechnology. Curr. Med. Chem. 2004, 11, 1241–1253. [Google Scholar]
- Rowinsky, E. The Vinca alkaloids. In Cancer Medicine, 6th ed.; Kufe, D.W., Pollock, R.E., Weichselbaum, R.R., Bast, R.C., Gansler, T.S., Holland, J.F., Frei, E., Eds.; BC Decker Publisher: Hamilton, ON, Canada, 2003; Volume 2. [Google Scholar]
- Schutz, F.A.; Bellmunt, J.; Rosenberg, J.E.; Choueiri, T.K. Vinflunine: Drug safety evaluation of this novel synthetic vinca alkaloid. Expert Opin. Drug Saf. 2011, 10, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Bennouna, J.; Delord, J.P.; Campone, M.; Nguyen, L. Vinflunine: A new microtubule inhibitor agent. Clin. Cancer Res. 2008, 14, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Grellier, P.; Sinou, V.; Garrea de Loubresse, N.; Bylen, E.; Boulard, Y.; Schrevel, J. Selective and reversible effects of vinca alkaloids on Trypanosoma cruzi epimastigote forms: Blockage of cytokinesis without inhibition of the organelle duplication. Cell Motil. Cytoskeleton. 1999, 42, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; de Luca, V. ATP-binding cassette transporter controls leaf surface secretion of anticancer drug components in Catharanthus roseus. Proc. Natl. Acad. Sci. USA 2013, 110, 15830–15835. [Google Scholar] [CrossRef] [PubMed]
- Roepke, J.; Salim, V.; Wu, M.; Thamm, A.M.; Murata, J.; Ploss, K.; Boland, W.; de Luca, V. Vinca drug components accumulate exclusively in leaf exudates of Madagascar periwinkle. Proc. Natl. Acad. Sci. USA 2010, 107, 15287–15292. [Google Scholar] [CrossRef] [PubMed]
- Naaranlahti, T.; Auriola, S.; Lapinjoki, S.P. Growth related dimerization of vindoline and catharanthine in Catharanthus roseus and effect of wounding on the process. Phytochemistry 1991, 30, 1451–1453. [Google Scholar] [CrossRef]
- Balsevich, J.; Bishop, G. Distribution of catharanthine, vindoline and 3',4'-anhydrovinblastine in the aerial parts of some Catharanthus roseus plants and the significance thereof in relation to alkaloid production in cultured cells. In Primary and Secondary Metabolism of Plant Cell Cultures II, 1st ed.; Kurz, W.G.W., Ed.; Springer-Verlag: Berlin, Germany, 1989; pp. 149–153. [Google Scholar]
- Goodbody, A.E.; Watson, C.D.; Chapple, C.C.S.; Vukovic, J.; Misawa, M. Extraction of 3',4'-anhydrovinblastine from Catharanthus roseus. Phytochemistry 1988, 27, 1713–1717. [Google Scholar] [CrossRef]
- Sottomayor, M.; Lopéz-Serrano, M.; DiCosmo, F.; Ros Barceló, A. Purification and characterization of α-3',4'-anhydrovinblastine synthase (peroxidase-like) from Catharanthus roseus (L.) G. Don. FEBS Lett. 1998, 428, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Carqueijeiro, I.; Noronha, H.; Duarte, P.; Geros, H.; Sottomayor, M. Vacuolar transport of the medicinal alkaloids from Catharanthus roseus is mediated by a proton-driven antiport. Plant Physiol. 2013, 162, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Verpoorte, R. Manipulating indole alkaloid production by Catharanthus roseus cell cultures in bioreactors: From biochemical processing to metabolic engineering. Phytochem. Rev. 2007, 6, 435–457. [Google Scholar] [CrossRef]
- Koul, M.; Lakra, N.S.; Chandra, R.; Chandra, S. Catharanthus roseus and prospects of its endophytes: A new avenue for production of bioactive metabolites. Int. J. Pharm. Sci. Res. 2013, 4, 2705–2716. [Google Scholar]
- Nayak, B.S.; Pereira, L.M.P. Catharanthus roseus flower extract has wound-healing activity in Sprague Dawley rats. BMC Complement. Altern. Med. 2006, 6, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Özçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Biol. 2011, 49, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Millon, M.J.; Newman, T.A.; Audinot, V.; Cussac, D.; Lejeune, F.; Nicolas, J.P.; Cogé, F.; Galizzi, J.P.; Boutin, J.A. Agonist and antagonist actions of Yohimbine as compared to fluparoxan at alpha (2)-adrenergic receptors (AR). Synapse 2002, 35, 79–95. [Google Scholar] [CrossRef]
- Briones-Martin-Del-Campo, M.; Orta-Zavalza, E.; Juarez-Cepeda, J.; Gutierrez-Escobedo, G.; Cañas-Villamar, I.; Castaño, I.; de Las Peñas, A. The oxidative stress response of the opportunistic fungal pathogen Candida glabrata. Rev. Iberoam. Micol. 2014, 31, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; He, H.P.; Di, Y.T.; Zhang, Y.; Hao, X.J. Catharoseumine, a new monoterpenoid indole alkaloid possessing a peroxy bridge from Catharanthus roseus. Tetrahedron Lett. 2012, 53, 1576–1578. [Google Scholar] [CrossRef]
- Vats, V.; Yadav, S.P.; Biswas, N.R.; Grover, J.K. Anti-cataract activity of Pterocarpus marsupium bark and Trigonella foenum-graecum seeds extract in alloxan diabetic rats. J. Ethnopharmacol. 2004, 93, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Hunag, Y.; Glinka, Y.; Prud’Homme, G.J.; Wang, Q. Gene therapy of diabetes using a novel GLP-1/IgG1-Fc fusion construct normalizes glucose levels in db/db mice. Gene Ther. 2006, 14, 162–172. [Google Scholar] [PubMed]
- Ghosh, S.; Suryawanshi, S.A. Effect of Vinca rosea extracts in treatment of alloxan diabetes in male albino rats. Indian J. Exp. Biol. 2001, 39, 748–759. [Google Scholar] [PubMed]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.C.; Mohd Rais, M.; Awang, K. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules 2013, 18, 9770–9784. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.W.; Worcel, M.; Wyllie, M. Yohimbine: A clinical review. Pharmacol. Ther. 2001, 91, 215–243. [Google Scholar] [CrossRef] [PubMed]
- Lebret, T.; Hervé, J.M.; Gorny, P.; Worcel, M.; Botto, H. Efficacy and safety of a novel combination of L-arginine glutamate and yohimbine hydrochloride: A new oral therapy for erectile dysfunction. Eur. Urol. 2002, 41, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.M.; Rothwell, N.J. Inflammation in central nervous system injury. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; O’Banion, M.K. Inflammatory processes in Alzheimer’s disease. J. Neuroimmunol. 2007, 184, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Scatena, R.; Martorona, G.E.; Bottani, P.; Botta, G.; Pastove, P.; Giardina, B. An update of pharmacological approaches to neurodegenerative diseases. Expert Opin. Investig. Drugs 2007, 16, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Manigandan, V.; Gurudeeban, S.; Satyavani, K.; Ramanathan, T. Molecular docking studies of Rhizophora mucronata alkaloids against neuroinflammatory marker cyclooxygenase 2. Int. J. Biol. Chem. 2014, 8, 91–99. [Google Scholar] [CrossRef]
- Wink, M.; Schmeller, T.; Latz-Brüning, B. Modes of action of allelochemical alkaloids: Interaction with neuroreceptors, DNA, and other molecular targets. J. Chem. Ecol. 1998, 24, 1881–1937. [Google Scholar] [CrossRef]
- Vakil, R.J. Rauwolfia serpentina in the treatment of high blood pressure. A review of the literature. Circulation 1955, 12, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Moudi, M.; Go, R.; Yien, C.Y.S.; Nazre, M. Vinca Alkaloids. Int. J. Prev. Med. 2013, 4, 1231–1235. [Google Scholar] [PubMed]
- Nirmala, M.J.; Samundeeswari, A.; Sankar, P.D. Natural plant resources in anti-cancer therapy-A review. Res. Plant Biol. 2011, 1, 1–14. [Google Scholar]
- Junaid, A.; Sheba, H.K.; Zahid, H.S.; Zohra, F.; Mehpara, M.; Mukthar, A.B.; Sekh, A.N.; Ilah, A.; Iffat, Z.A.; Saeed, A.K.; et al. Catharanthus roseus (L.) G. Don. An important drug: It’s applications and production. Pharm. Glob. IJCP 2010, 4, 1–16. [Google Scholar]
- Rai, V.; Tandon, P.K.; Khatoon, S. Effect of chromium on antioxidant potential of Catharanthus roseus varieties and production of their anticancer alkaloids: Vincristine and vinblastine. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Kutney, J.; Mayer, L.; Schmidt, B. Anhydrovinblastine for the Treatment of Cervical and Lung Cancer. EP0969839 A1.
- Zhu, R.H.; Li, H.D.; Cai, H.L.; Jiang, Z.P.; Xu, P.; Dai, L.B.; Peng, W.X. Validated HILIC–MS/MS assay for determination of vindesine in human plasma: Application to a population pharmacokinetic study. J. Pharm. Biomed. Anal. 2014, 96, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Rhomberg, W.; Eiter, H.; Schmid, F.; Saely, C. Razoxane and vindesine in advanced soft tissue sarcomas: Impact on metastasis, survival and radiation response. Anticancer Res. 2007, 27, 3609–3614. [Google Scholar] [PubMed]
- Gregory, R.K.; Smith, I.E. Vinorelbine—A clinical review. Br. J. Cancer 2000, 82, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Aapro, M.; Finek, J. Oral vinorelbine in metastatic breast cancer: A review of current clinical trial results. Cancer Treat. Rev. 2012, 38, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Bellmunt, J.; Fougeray, R.; Rosenberg, J.E.; von der Maase, H.; Schutz, F.A.; Salhi, Y.; Culine, S.; Choueiri, T.K. Long-term survival results of a randomized phase III trial of vinflunine plus best supportive care versus best supportive care alone in advanced urothelial carcinoma patients after failure of platinum-based chemotherapy. Ann. Oncol. 2013, 24, 1466–1472. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.A.; Lawrence, N.J.; McGown, A.T. Tubulin as a target for anticancer drugs: Agents which interact with the mitotic spindle. Med. Res. Rev. 1998, 18, 259–296. [Google Scholar] [CrossRef] [PubMed]
- Gigant, B.; Wang, C.; Ravelli, R.B.; Roussi, F.; Steinmetz, M.O.; Curmi, P.A.; Sobel, A.; Knossow, M. Structural basis for the regulation of tubulin by vinblastine. Nature 2005, 435, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Okouneva, T.; Hill, B.T.; Wilson, L.; Jordan, M.A. The effects of vinflunine, vinorelbine, and vinblastine on centromere dynamics. Mol. Cancer Ther. 2003, 2, 427–436. [Google Scholar] [PubMed]
- Gidding, C.E.; Kellie, S.J.; Kamps, W.A.; de Graaf, S.S. Vincristine revisited. Crit. Rev. Oncol. Hematol. 1999, 29, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Takanari, H.; Yosida, T.; Morita, J.; Izutsu, K.; Ito, T. Instability of pleomorphic tubulin paracrystals artificially induced by Vinca alkaloids in tissue-cultured cells. Biol. Cell 1990, 70, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Daly, E.M.; Taylor, R.E. Entropy and enthalpy in the activity of tubulin-based antimitotic agents. Curr. Chem. Biol. 2009, 3, 47–59. [Google Scholar] [CrossRef]
- Sertel, S.; Fu, Y.; Zu, Y.; Rebacz, B.; Konkimalla, B.; Plinkert, P.K.; Krämer, A.; Gertsch, J.; Efferth, T. Molecular docking and pharmacogenomics of Vinca alkaloid and their monomeric precursor, vindoline and catharanthine. Biochem. Pharmacol. 2011, 81, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Coderch, C.; Morreale, A.; Gago, F. Tubulin-based structure-affinity relationship for antimitotic Vinca alkaloid. Anticancer Agents Med. Chem. 2012, 12, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Makarov, A.A.; Tsvetkov, P.O.; Villard, C.; Esquieu, D.; Pourroy, B.; Fahy, J.; Braguer, D.; Peyrot, V.; Lafitte, D. Vinflunine, a novel microtubule inhibitor, suppresses calmodulin interaction with the microtubule-associated protein STOP. Biochemistry 2007, 46, 14899–14906. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkov, F.O.; Kulikova, A.A.; Devred, F.; Zernii, E.; Lafitte, D.; Makarov, A.A. Thermodynamics of calmodulin and tubulin binding to the vinca-alkaloid vinorelbine. Mol. Biol. 2011, 45, 697–702. [Google Scholar] [CrossRef]
- Barbier, P.; Tsvetkov, P.O.; Breuzard, G.; Devred, F. Deciphering the molecular mechanism of anti-tubulin plant derived drugs. Phytochem. Rev. 2013, 13, 157–169. [Google Scholar] [CrossRef]
- Deus, B.; Zenk, M.H. Exploitation of plant cells for the production of natural compounds. Biotechnol. Bioeng. 1982, 24, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Kurz, W.G.W.; Chatson, K.B.; Constabel, F. Biosynthesis and accumulation of indole alkaloids in cell suspension cultures of Catharanthus roseus cultivars. In Primary and Secondary Metabolism of Plant Cell Cultures, 1st ed.; Neumann, K.H., Barz, W., Reinhard, E., Eds.; Springer-Verlag: Berlin, Germany, 1985; Volume 5, pp. 143–153. [Google Scholar]
- Ganapathi, B.; Kargi, F. Recent Advances in Indole Alkaloid Production by Catharanthus roseus (Periwinkle). J. Exp. Bot. 1990, 41, 259–267. [Google Scholar] [CrossRef]
- Mannonen, L.; Toivonen, L.; Kauppinen, V.C. Effects of long term preservation on growth and productivity of Panax ginseng and Catharanthus roseus cell culture. Plant Cell Rep. 1990, 9, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Bachiri, Y.; Gazeau, C.; Hansz, J.; Morisset, C.; Dereuddre, J. Successful cryopreservation of suspension cells by encapsulation dehydration. Plant Cell Tissue Organ Cult. 1995, 43, 241–248. [Google Scholar]
- El-Sayed, M.; Verpoorte, R. Catharanthus terpenoid indole alkaloids: Biosynthesis and regulation. Phytochem. Rev. 2007, 6, 277–305. [Google Scholar] [CrossRef]
- Endo, T.; Goodbody, A.E.; Misawa, M. Alkaloid production in root and shoot cultures of Catharanthus roseus. Planta Med. 1987, 53, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.K.; Shasany, A.K.; Verma, R.K.; Gupta, M.M.; Mathur, A.K.; Khanuja, P.S.P. Influence of cellular differentiation and elicitation on intermediate and late steps of terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Protoplasma 2010, 24, 35–47. [Google Scholar] [CrossRef]
- Bhadra, R.; Vani, S.; Shanks, J.V. Production of indole alkaloids by selected hairy root lines of Catharanthus roseus. Biotechnol. Bioeng. 1993, 41, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Kwak, S.S.; Choi, C.Y.; Liu, J.R. An interchangeable system of hairy root and cell suspension cultures of Catharanthus roseus for indole alkaloid production. Plant Cell Rep. 1995, 15, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Palazón, J.; Cusidó, R.M.; Gonzalo, J.; Bonfill, M.; Morales, C.; Pinol, T. Relation between the amount of rolC gene product and indole alkaloid accumulation in Catharanthus roseus transformed root cultures. J. Plant Physiol. 1998, 153, 712–718. [Google Scholar] [CrossRef]
- Van der Heijden, R.; Verpoorte, R.; ten Hoopen, H.J.G. Cell and tissue cultures of Catharanthus roseus (L.) G. Don: A literature survey. Plant Cell Tissue Organ Cult. 1989, 18, 231–280. [Google Scholar] [CrossRef]
- Zhao, J.; Hu, Q.; Guo, Y.Q.; Zhu, W.H. Effects of stress factors, bioregulators, and synthetic precursors on indole alkaloid production in compact callus clusters cultures of Catharanthus roseus. Appl. Microbiol. Biotechnol. 2001, 55, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Liu, Z.P.; Liu, L.; Liu, C. Effects of NaCl on the growth and alkaloid content of Catharanthus roseus seedlings. J. Appl. Ecol. 2008, 19, 2143–2148. [Google Scholar]
- Schlatmann, J.E.; Koolhaas, C.M.A.; Vinke, J.L.; ten Hoopen, H.J.G.; Heijnen, J.J. The role of glucose in ajmalicine production by Catharanthus roseus cell cultures. Biotechnol. Bioeng. 1995, 47, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhu, W.H.; Hu, Q.; He, X.W. Enhanced indole alkaloid production in suspension compact callus clusters of Catharanthus roseus: Impacts of plant growth regulators and sucrose. Plant Growth Reg. 2001, 33, 33–41. [Google Scholar] [CrossRef]
- Jung, K.H.; Kwak, S.S.; Kim, S.W.; Lee, H.; Choi, C.Y.; Liu, J.R. Improvement of the catharanthine productivity in hairy root cultures of Catharanthus roseus by using monosaccharides as a carbon source. Biotechnol. Lett. 1992, 14, 695–700. [Google Scholar] [CrossRef]
- Almagro, L.; López-Pérez, A.J.; Pedreño, M.A. New method to enhance ajmalicine production in Catharanthus roseus cell cultures based on the use of cyclodextrins. Biotechnol. Lett. 2010, 33, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.T.; Shuler, M.L. The effect of inoculum density and conditioned medium on the production of ajmalicine and catharanthine from immobilized Catharanthus roseus cells. Biotechnol. Bioeng. 2000, 67, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Asada, M.; Shuler, M.L. Stimulation of ajmalicine production and excretion from Catharanthus roseus: Effects of adsorption in situ, elicitors, and alginate immobilization. Appl. Microbiol. Biotechnol. 1989, 30, 475–481. [Google Scholar]
- Almagro, L.; Sabater-Jara, A.B.; Belchi-Navarro, S.; Fernandez-Perez, F.; Bru, R.; Pedreño, M.A. Effect of UV light on secondary metabolite biosynthesis in plant cell cultures elicited with cyclodextrins and methyljasmonate. In Plants and Environment, 1st ed.; Vasanthaiah, H.K.N., Kambiranda, D., Eds.; InTech Janeza Trdine: Rijeka, Croatia, 2011; Volume 1, pp. 115–136. [Google Scholar]
- Satdive, R.K.; Fulzele, D.P.; Eapen, S. Studies on Production of Ajmalicine in Shake Flasks by Multiple Shoot Cultures of Catharanthus roseus. Biotechnol. Prog. 2003, 19, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Lee-Parson, C.W.T.; Shuler, M.L. Sparge gas composition affects biomass and ajmalicine production from immobilized cell cultures of Catharanthus roseus. Enzym. Microb. Technol. 2005, 37, 424–434. [Google Scholar] [CrossRef]
- El-Sayed, M.; Verpoorte, R. Effect of phytohormones on growth and alkaloid accumulation by a Catharanthus roseus cell suspension cultures fed with alkaloid precursors tryptamine and loganin. Plant Cell Tissue Organ Cult. 2002, 68, 265–270. [Google Scholar] [CrossRef]
- Morgan, J.A.; Shanks, J.V. Determination of metabolic rate-limitations by precursor feeding in Catharanthus roseus hairy root cultures. J. Biotechnol. 2000, 79, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Peebles, C.A.; Shanks, J.V.; San, K.Y. Effect of sodium nitroprusside on growth and terpenoid indole alkaloid production in Catharanthus roseus hairy root cultures. Biotechnol. Prog. 2011, 27, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Goldhaber-Pasillas, G.D.; Mustafa, N.R.; Verpoorte, R. Jasmonic Acid Effect on the Fatty Acid and Terpenoid Indole Alkaloid Accumulation in Cell Suspension Cultures of Catharanthus roseus. Molecules 2014, 19, 10242–10260. [Google Scholar] [CrossRef] [PubMed]
- Almagro, L.; Gutierrez, J.; Pedreño, M.A.; Sottomayor, M. Synergistic and additive influence of cyclodextrins and methyl jasmonate on the expression of the terpenoid indole alkaloid pathway genes and metabolites in Catharanthus roseus cell cultures. Plant Cell Tissue Organ Cult. 2014, 119, 543–551. [Google Scholar] [CrossRef]
- Ruiz-May, E.; Galaz-Ávalos, R.M.; Loyola-Vargas, V.M. Differential secretion and accumulation of terpene indole alkaloids in hairy roots of Catharanthus roseus treated with methyl jasmonate. Mol. Biotechnol. 2009, 41, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Namdeo, A.; Patil, S.; Fulzele, D.P. Influence of fungal elicitors on production of ajmalicine by cell cultures of Catharanthus roseus. Biotechnol. Prog. 2002, 18, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chen, Z.; Lu, L.; Jin, H.; Sun, L.; Yu, Q.; Xu, H.; Yang, F.; Fu, M.; Li, S.; et al. Interaction between abscisic acid and nitric oxide in PB90 induced catharanthine biosynthesis of Catharanthus roseus cell suspension cultures. Biotechnol. Prog. 2013, 29, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Ramani, S.; Jayabaskaran, C. Enhanced catharanthine and vindoline production in suspension cultures of Catharanthus roseus by ultraviolet-B light. J. Mol. Signal. 2008, 3, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhu, W.H.; Hu, Q. Enhanced catharanthine production in Catharanthus roseus cell cultures by combined elicitor treatment in shake flasks and bioreactors. Enzym. Microb. Technol. 2001, 28, 673–681. [Google Scholar] [CrossRef]
- Canel, C.; Lopes-Cardoso, M.I.; Whitmer, S.; van der Fits, L.; Pasquali, G.; van der Heijden, R.; Hoge, J.H.C.; Verpoorte, R. Effects of over-expression of strictosidine synthase and tryptophan decarboxylase on alkaloid production by cell cultures of Catharanthus roseus. Planta 1998, 205, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, S.; van der Heijden, R.; Verpoorte, R. Effect of precursor feeding on alkaloid accumulation by a tryptophan decarboxylase over-expressing transgenic cell line T22 of Catharanthus roseus. J. Biotechnol. 2002, 96, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Magnotta, M.; Murata, J.; Chen, J.; de Luca, V. Expression of deacetylvindoline-4-O-acetyltransferase in Catharanthus roseus hairy roots. Phytochemistry 2007, 68, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Jaggi, M.; Kumar, S.; Sinha, A.K. Overexpression of an apoplastic peroxidase gene CrPrx in transgenic hairy root lines of Catharanthus roseus. Appl. Microbiol. Biotechnol. 2011, 90, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Peebles, C.; Sander, G.W.; Hughes, E.H.; Peacock, R.; Shanks, J.V.; San, K.Y. The expression of 1-deoxy-d-xylulose synthase and geraniol-10-hydroxylase or anthranilate synthase increases terpenoid indole alkaloid accumulation in Catharanthus roseus hairy roots. Metab. Eng. 2011, 13, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Smita, S.S.; Singh, A.K.; Shanker, K.; Nagegowda, D.A. Heteromeric and homomeric geranyl diphosphate synthases from Catharanthus roseus and their role in monoterpene indole alkaloid biosynthesis. Mol. Plant 2013, 6, 1531–1549. [Google Scholar] [CrossRef] [PubMed]
- Hallard, D.; van der Heijden, R.; Verpoorte, R.; Lopez-Cardoso, I.; Pasquali, G.; Memelink, J.; Hoge, J.H.C. Suspension cultured transgenic cells of Nicotiana tabacum expressing tryptophan decarboxylase and strictosidine synthase cDNAs from Catharanthus roseus produce strictosidine upon feeding of secologanin. Plant Cell Rep. 1997, 17, 50–54. [Google Scholar] [CrossRef]
- Hallard, D. Transgenic Plant Cells for the Production of Indole Alkaloids. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 2000. [Google Scholar]
- Verpoorte, R.; Contin, A.; Memelink, J. Biotechnology for production of plant secondary metabolites. Phytochem. Rev. 2002, 1, 13–25. [Google Scholar] [CrossRef]
- Suttipanta, N.; Pattanaik, S.; Kulshrestha, M.; Patra, B.; Singh, S.K.; Yuan, L. The transcription factor CrWRKY1 positively regulates the terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 2011, 157, 2081–2093. [Google Scholar] [CrossRef] [PubMed]
- Raina, S.K.; Wankhede, D.P.; Jaggi, M.; Singh, P.; Jalmi, S.K.; Raghuram, B.; Sheikh, A.H.; Sinha, A.K. CrMPK3, a mitogen activated protein kinase from Catharanthus roseus and its possible role in stress induced biosynthesis of monoterpenoid indole alkaloids. BMC Plant Biol. 2012, 12, 134. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.X.; Liu, D.H.; Wang, Y.L.; Cui, L.J.; Ren, W.W.; Sun, F. Overexpression of transcriptional factor ORCA3 increases the accumulation of catharanthine and vindoline in Catharanthus roseus hairy roots. Russ. J. Plant Physiol. 2011, 58, 415–422. [Google Scholar] [CrossRef]
- Liu, D.H.; Ren, W.W.; Cui, L.J.; Zhang, L.D.; Sun, X.F.; Tang, K.X. Enhanced accumulation of catharanthine and vindoline in Catharanthus roseus hairy roots by overexpresion of transcriptional factor ORCA2. Afr. J. Biotechnol. 2011, 10, 3260–3268. [Google Scholar] [CrossRef]
- Pan, Q.; Wang, Q.; Yuan, F.; Xing, S.; Zhao, J.; Choi, Y.H.; Verpoorte, R.; Tian, Y.; Wang, G.; Tang, K. Overexpression of ORCA3 and G10H in Catharanthus roseus plants regulated alkaloid biosynthesis and metabolism revealed by NMR-metabolomics. PLoS One 2012, 7, e43038. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, S.; Verpoorte, R.; Canel, C. Influence of auxins on alkaloid accumulation by a transgenic cell line of Catharanthus roseus. Plant Cell Tissue Organ Cult. 1998, 53, 135–141. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, W.H.; Hu, Q. Effects of light and plant growth regulators on the biosynthesis of vindoline and other indole alkaloids in Catharanthus roseus callus cultures. Plant Growth Reg. 2001, 33, 43–49. [Google Scholar] [CrossRef]
- Lee, C.W.T.; Shuler, M.L. Different shake flask closures alter gas phase composition and ajmalicine production in Catharanthus roseus cell suspensions. Biotechnol. Technol. 1991, 5, 173–178. [Google Scholar] [CrossRef]
- El-Sayed, M.; Choi, Y.H.; Fréderichrich, M.; Roytrakul, S.; Verpoorte, R. Alkaloid accumulation in Catharanthus roseus cell suspension cultures fed with stemmadenine. Biotechnol. Lett. 2004, 26, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, S.; van der Heijden, R.; Verpoorte, R. Effect of precursor feeding on alkaloid accumulation by a strictosidine synthase over-expressing transgenic cell line S1 of Catharanthus roseus. Plant Cell Tissue Organ Cult. 2000, 69, 85–93. [Google Scholar] [CrossRef]
- Moreno, P.R.H.; van der Heijden, R.; Verpoorte, R. Effect of terpenoid precursor feeding and elicitation on formation of indole alkaloids in cell suspension cultures of Catharanthus roseus. Plant Cell Rep. 1993, 12, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Krishnamurthy, R. Elicitors in plant tissue culture. J. Pharmacogn. Phytochem. 2013, 2, 60–65. [Google Scholar]
- Seitz, H.U.; Eilert, U.; de Luca, V.; Kurz, W.G.W. Elicitor mediated induction of phenylalanine ammonia lyase and tryptophan decarboxylase; accumulation of phenols and indole alkaloids in cell suspension cultures of Catharanthus roseus. Plant Cell Tissue Organ Cult. 1989, 18, 71–78. [Google Scholar] [CrossRef]
- Kargi, F.; Ganapathi, B. Effects of precursor stimulating agents on formation of indole formation by C. roseus in a biofilm reactor. Enzym. Microb. Technol. 1991, 13, 643–647. [Google Scholar] [CrossRef]
- Hirata, K.; Horiuchi, M.; Asada, M.; Ando, T.; Miyamota, K.; Miura, Y. Stimulation of dimeric alkaloid production by near ultraviolet light in multiple shoot cultures of Catharanthus roseus. Ferment Bioeng. 1992, 74, 222–225. [Google Scholar] [CrossRef]
- Lee-Parsons, C.W.; Ertürk, S.; Tengtrakool, J. Enhancement of ajmalicine production in Catharanthus roseus cell cultures with methyl jasmonate is dependent on timing and dosage of elicitation. Biotechnol. Lett. 2004, 26, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Dong, J. Elicitor-induced nitric oxide burst is essential for triggering catharanthine synthesis in Catharanthus roseus suspension cells. Appl. Microbiol. Biotechnol. 2005, 67, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Memelink, J.; Gantet, P. Transcription factors involved in terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Phytochem. Rev. 2007, 6, 353–362. [Google Scholar] [CrossRef]
- Godoy-Hernández, G.C.; Loyola-Vargas, V.M. Effect of acetylsalicylic acid on secondary metabolism of Catharanthus roseus tumor suspension cultures. Plant Cell Rep. 1997, 16, 287–290. [Google Scholar] [CrossRef]
- Binder, B.Y.; Peebles, C.A.; Shanks, J.V.; San, K.Y. The effects of UV-B stress on the production of terpenoid indole alkaloids in Catharanthus roseus hairy roots. Biotechnol. Prog. 2009, 25, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Courdavault, V.; Lanoue, A.; Mahroug, S.; Guihur, A.; Blanc, N.; Giglioli-Guivarc’h, N.; St-Pierre, B.; Burlat, V. Strictosidine activation in Apocynaceae: Towards a “nuclear time bomb”? BMC Plant Biol. 2010, 10, 182. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Guihur, A.; Poutrain, P.; Héricourt, F.; Mahroug, S.; St-Pierre, B.; Burlat, V.; Courdavault, V. Spatial organization of the vindoline biosynthetic pathway in Catharanthus roseus. J. Plant Physiol. 2011, 168, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.M.R.; Hilliou, F.; Duarte, P.; Pereira, L.G.; Almeida, I.; Leech, M.; Memelink, J.; Barcelo, A.R.; Sottomayor, M. Molecular cloning and characterization of a vacuolar class III peroxidase involved in the metabolism of anticancer alkaloids in Catharanthus roseus. Plant Physiol. 2008, 146, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Van Der Fits, L.; Memelink, J. The jasmonate-inducible AP2/ERF-domain transcription factor ORCA3 activates gene expression via interaction with a jasmonate-responsive promoter element. Plant J. 2001, 25, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Wei, S. Methyl jasmonic acid induced expression pattern of terpenoid indole alkaloid pathway genes in Catharanthus roseus seedlings. Plant Growth Regul. 2010, 61, 243–251. [Google Scholar] [CrossRef]
- Zhou, M.L.; Zhu, X.M.; Shao, J.R.; Wu, Y.M.; Tang, Y.X. Transcriptional response of the catharanthine biosynthesis pathway to methyl jasmonate/nitric oxide elicitation in Catharanthus roseus hairy root culture. Appl. Microbiol. Biotechnol. 2010, 88, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, G.; Goddijn, O.J.; de Waal, A.; Verpoorte, R.; Schilperoort, R.A.; Hoge, J.H.C.; Memelink, J. Coordinated regulation of two indole alkaloid biosynthetic genes from Catharanthus roseus by auxin and elicitors. Plant Mol. Biol. 1992, 18, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Menke, F.L.; Parchmann, S.; Mueller, M.J.; Kijne, J.W.; Memelink, J. Involvement of the octadecanoid pathway and protein phosphorylation in fungal elicitor-induced expression of terpenoid indole alkaloid biosynthetic genes in Catharanthus roseus. Plant Physiol. 1999, 119, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Bru, R.; Sellés, S.; Casado-Vela, J.; Belchí-Navarro, S.; Pedreño, M.A. Modified cyclodextrins are chemically defined glucan inducers of defense responses in grapevine cell cultures. J. Agric. Food Chem. 2006, 54, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Capell, T.; Christou, P. Progress in plant metabolic engineering. Curr. Opin. Biotechnol. 2004, 15, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.A.; Roberts, S.C. Metabolic engineering approaches for production of biochemicals in food and medicinal plants. Curr. Opin. Biotechnol. 2014, 26, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Zárate, R.; Verpoorte, R. Strategies for the genetic modification of the medicinal plant Catharanthus roseus (L.) G. Don. Phytochem. Rev. 2007, 6, 475–491. [Google Scholar] [CrossRef]
- Geerlings, A.; Redondo, F.J.; Contin, A.; Memelink, J.; van der Heijden, R.; Verpoorte, R. Biotransformation of tryptamine and secologanin into plant terpenoid indole alkaloids by transgenic yeast. Appl. Microbiol. Biotechnol. 2001, 56, 420–424. [Google Scholar] [CrossRef]
- Chatel, G.; Montiel, G.; Pre, M.; Memelink, J.; Thiersault, M.; Saint-Pierre, B.; Doireau, P.; Gantet, P. CrMYC1, a Catharanthus roseus elicitor- and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter. J. Exp. Bot. 2003, 54, 2587–2588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hedhili, S.; Montiel, G.; Zhang, Y.; Chatel, G.; Pre, M.; Gantet, P.; Memelink, J. The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J. 2011, 67, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Van der Fits, L.; Memelink, J. ORCA3, a jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 2000, 289, 295–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.T.; Liu, H.; Gao, X.S.; Zhang, H.X. Overexpression of G10H and ORCA3 in the hairy roots of Catharanthus roseus improves catharanthine production. Plant Cell Rep. 2010, 29, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Leopold, A.L.; Sander, G.W.; Shanks, J.V.; Zhao, L.; Gibson, S.I. The ORCA2 transcription factor plays a key role in regulation of the terpenoid indole alkaloid pathway. BMC Plant Biol. 2013, 13, 155. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almagro, L.; Fernández-Pérez, F.; Pedreño, M.A. Indole Alkaloids from Catharanthus roseus: Bioproduction and Their Effect on Human Health. Molecules 2015, 20, 2973-3000. https://doi.org/10.3390/molecules20022973
Almagro L, Fernández-Pérez F, Pedreño MA. Indole Alkaloids from Catharanthus roseus: Bioproduction and Their Effect on Human Health. Molecules. 2015; 20(2):2973-3000. https://doi.org/10.3390/molecules20022973
Chicago/Turabian StyleAlmagro, Lorena, Francisco Fernández-Pérez, and Maria Angeles Pedreño. 2015. "Indole Alkaloids from Catharanthus roseus: Bioproduction and Their Effect on Human Health" Molecules 20, no. 2: 2973-3000. https://doi.org/10.3390/molecules20022973