
PLS-Prediction and Confirmation of Hydrojuglone Glucoside as the Antitrypanosomal Constituent of Juglans Spp.



Abstract
:1. Introduction
2. Results and Discussion

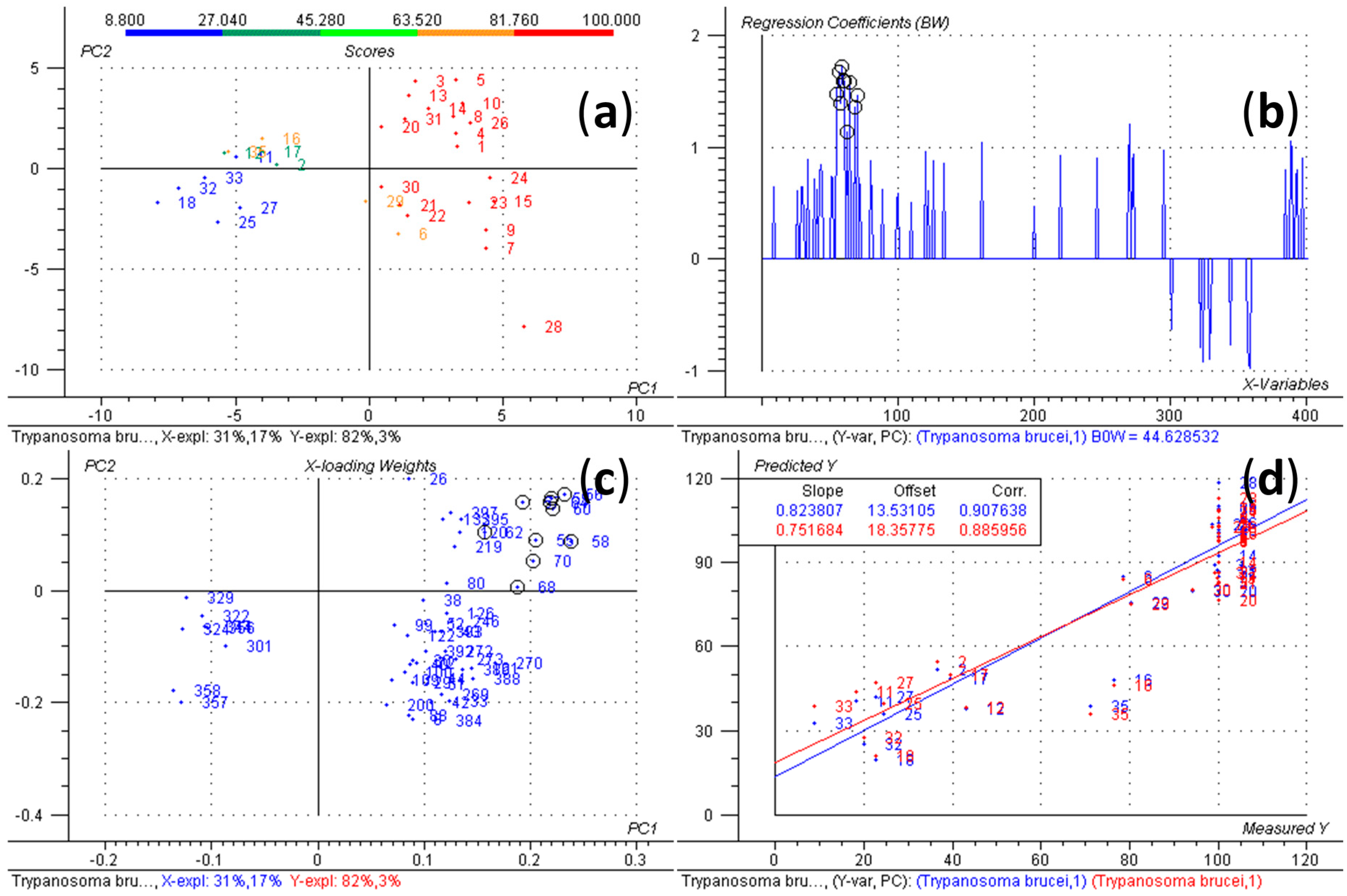{kind=link}
{kind=link}
| No. | Species | Plant Part | State | Drying | Extraction | % Inhibition at 2 μg/mL Against | % Inhibition at 10 μg/mL Against | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TB | TC | LD | TB | TC | LD | ||||||
| 1 | JR | Male flower | Green | FD | D | 83.0 | 6.8 | 21.5 | 100.0 | 0.9 | 29.9 |
| 2 | JR | Pericarp | Green | FD | E | 4.7 | 7.9 | 16.7 | 36.7 | 11.7 | 21.1 |
| 3 | JR | Bark | n.a. | FD | D | 99.5 | 5.9 | 16.6 | 99.1 | 12.5 | 54.4 |
| 4 | JC2 | Leaf | Green | FD | D | 68.9 | 0.0 | 0.0 | 100.0 | 7.7 | 0.0 |
| 5 | JC1 | Pericarp | Green | FD | D | 23.2 | 2.6 | 24.6 | 100.0 | 0.9 | 23.3 |
| 6 | JR | Leaf | Green | FD | F | 15.9 | 0.0 | 24.9 | 78.4 | 0.0 | 23.6 |
| 7 | JN | Leaf | Green | FD | D | 45.2 | 0.4 | 26.4 | 100.0 | 5.7 | 24.2 |
| 8 | JR | Pericarp | Green | FD | G | 20.0 | 5.3 | 25.5 | 100.0 | 9.3 | 27.4 |
| 9 | JN | Leaf | Green | FD | F | 85.6 | 0.0 | 28.3 | 100.0 | 10.3 | 40.8 |
| 10 | JR | Pericarp | Green | FD | D | 24.1 | 0.0 | 24.1 | 100.0 | 0.0 | 28.7 |
| 11 | JR | Male flower | Senescent | FD | D | 9.7 | 11.1 | 23.5 | 18.2 | 5.5 | 20.3 |
| 12 | JN | Pericarp | Senescent | None | A | 5.9 | 0.0 | 19.1 | 43.0 | 23.8 | 18.4 |
| 13 | JC2 | Pericarp | Green | FD | D | 15.5 | 0.0 | 16.3 | 99.8 | 13.3 | 23.2 |
| 14 | JR | Pericarp | Green | FD | D | 26.1 | 4.4 | 16.6 | 100.0 | 23.5 | 24.5 |
| 15 | JC1 | Leaf | Senescent | FD | D | 95.6 | 0.2 | 2.6 | 100.0 | 3.0 | 0.0 |
| 16 | JN | Pericarp | Green | None | A | 15.1 | 0.0 | 23.9 | 76.5 | 15.7 | 14.3 |
| 17 | JN | Pericarp | Senescent | None | B | 12.3 | 5.1 | 22.5 | 39.4 | 10.1 | 19.3 |
| 18 | JR | Leaf | Senescent | FD | C | 9.5 | 11.8 | 25.5 | 22.7 | 10.5 | 27.6 |
| 19 | JR | Leaf | Green | FD | C | 73.4 | 0.0 | 26.9 | 100.0 | 8.3 | 57.9 |
| 20 | JN | Pericarp | Green | None | B | 42.7 | 0.0 | 24.7 | 100.0 | 12.9 | 23.5 |
| 21 | JR | Leaf | Senescent | FD | B | 30.8 | 0.0 | 24.5 | 100.0 | 11.9 | 27.7 |
| 22 | JR | Leaf | Green | FD | D | 51.8 | 3.1 | 21.1 | 100.0 | 1.7 | 27.7 |
| 23 | JR | Leaf | Green | FD | D | 17.2 | 9.1 | 18.0 | 98.7 | 4.0 | 19.8 |
| 24 | JC1 | Leaf | Green | FD | D | 60.7 | 6.8 | 15.6 | 100.0 | 0.0 | 23.9 |
| 25 | JR | Leaf | Senescent | AD | D | 2.5 | 8.5 | 16.4 | 24.6 | 18.7 | 25.2 |
| 26 | JR | Pericarp | Green | FD | B | 43.4 | 5.6 | 12.1 | 100.0 | 4.4 | 50.8 |
| 27 | JR | Leaf | Green | AD | D | 13.9 | 6.9 | 26.5 | 22.8 | 4.7 | 22.1 |
| 28 | JN | Leaf | Senescent | FD | D | 76.3 | 8.1 | 26.2 | 100.0 | 0.0 | 20.1 |
| 29 | JC2 | Leaf | Senescent | FD | D | 15.3 | 13.0 | 25.9 | 80.4 | 15.2 | 18.6 |
| 30 | JR | Leaf | Green | FD | B | 15.2 | 6.7 | 27.6 | 94.3 | 15.4 | 18.2 |
| 31 | JN | Pericarp | Green | FD | D | 54.4 | 9.7 | 22.7 | 100.0 | 0.0 | 29.0 |
| 32 | JN | Leaf | Green | FD | C | 8.7 | 12.1 | 24.3 | 19.9 | 11.7 | 38.7 |
| 33 | JC1 | Leaf | Green | FD | C | 5.7 | 8.4 | 20.8 | 8.8 | 0.0 | 34.9 |
| 34 | JR | Pericarp | Green | FD | C | 69.4 | 8.7 | 29.2 | 100.0 | 18.1 | 100.0 |
| 35 | JN | Pericarp | Green | FD | C | 5.7 | 4.8 | 17.3 | 71.0 | 6.0 | 27.8 |
| 36 | JN | Pericarp | Green | AD | C | 100.0 | 0.0 | 20.7 | 100.0 | 16.1 | 69.3 |

| Mass Spectrum of (1) | Bucket Table | |||
|---|---|---|---|---|
| m/z (Rel. Int.) | Ion | No. | Name | Loading Weight on PC1 |
| 699.1985 (7) | [2M + Na]+ | 54 | 10.8 min: 699.19 m/z | n.a. |
| 677.2157 (10) | [2M + H]+ | 67 | 10.9 min: 677.21 m/z | n.a. |
| 501.1665 (11) | [2M − C10H8O3 + H]+ | 64 | 10.9 min: 501.18 m/z | 0.218 |
| 377.0669 (2) | [M + K]+ | 62 | 10.9 min: 377.10 m/z | 0.157 |
| 361.0930 (9) | [M + Na]+ | 61 | 10.9 min: 361.10 m/z | n.a. |
| 356.1378 (3) | [M + NH4]+ | 60 | 10.9 min: 356.15 m/z | 0.221 |
| 339.1117 (43) | [M + H]+ | 70 | 11.0 min: 339.12 m/z | 0.202 |
| 321.1000 (4) | fragments of [M + H]+ | 59 | 10.9 min: 321.11 m/z | 0.220 |
| 303.0907 (7) | 58 | 10.9 min: 303.10 m/z | 0.238 | |
| 285.0785 (2) | 57 | 10.9 min: 285.09 m/z | 0.192 | |
| 243.0685 (3) | 56 | 10.9 min: 243.08 m/z | 0.231 | |
| 201.0576 (3) | 55 | 10.9 min: 201.06 m/z | 0.205 | |
| 177.0570 (100) | 68 | 11.0 min: 177.06 m/z | 0.187 | |

| Compound | IC50 (μM) | SI | |||||
|---|---|---|---|---|---|---|---|
| TB | TC | LD | L6 | TB | TC | LD | |
| Hydrojuglone glucoside (1) | 6.12 | 169.40 | 16.65 | 122.48 | 20.0 | 0.7 | 7.4 |
| Juglone | 1.62 | >100 | 2.02 | 21.03 | 13.0 | <0.1 | 10.4 |
| 1,4-Naphthoquinone | 0.58 | 9.24 | 2.38 | 6.39 | 11.1 | 0.7 | 2.7 |
| Lawson | 101.15 | 37.36 | 1.99 | 20.63 | 0.2 | 0.6 | 10.3 |
| 2,2′-bis-(3-Hydroxy-1,4-naphthoquinone) | 68.79 | 236.99 | 7.34 | 61.56 | 0.9 | 0.3 | 8.4 |
| Plumbagin | 0.49 | 3.59 | 0.88 | 2.87 | 5.9 | 0.8 | 3.3 |
| Lapachol | 16.40 | 17.56 | 3.33 | 26.57 | 1.6 | 1.5 | 8.0 |
| Shikonin | 0.03 | 0.27 | 0.12 | 0.06 | 2.1 | 0.2 | 0.5 |
| Arbutin (1,4-benzoquinone glucoside) | 236.40 | 244.10 | >367 | >367 | >1.5 | >1.5 | n.a |
| Positive controls | 0.005 | 2.33 | 0.18 | 0.014 | |||
3. Experimental Section
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extract Preparation
3.4. LC-MS Measurements
3.5. Antiprotozoal Assays
3.6. Calculation of PLS Models
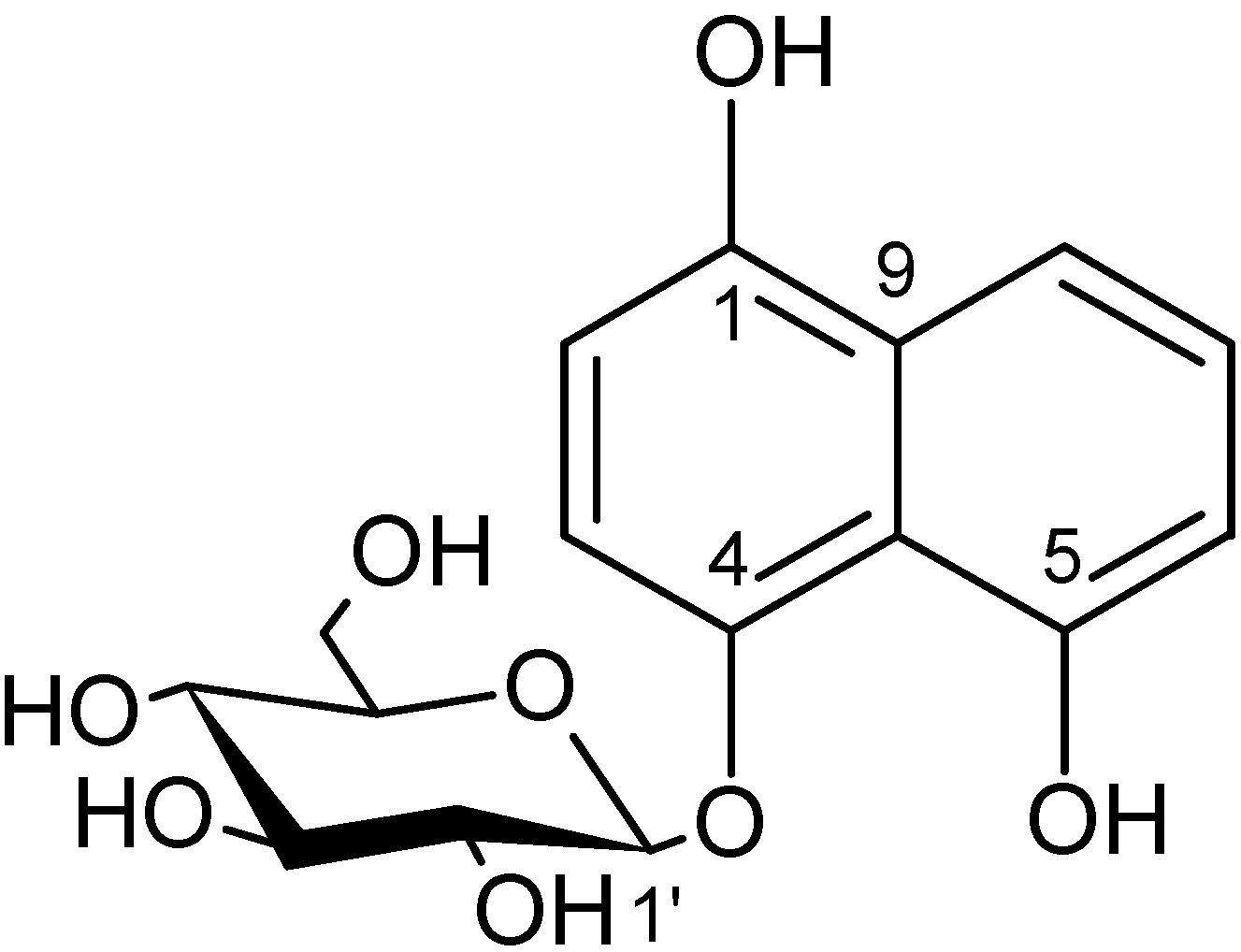
3.7. Isolation and Structure Elucidation of (1)
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads against Protozoan Neglected Diseases—Part I. Curr. Med. Chem. 2012, 19, 2128–2175. [Google Scholar] [PubMed]
- World Health Organization (WHO). Neglected Tropical Diseases. Available online: http://www.who.int/neglected_diseases/diseases/en/ (accessed on 27 February 2015).
- Dai, J.R.; Decosterd, L.A.; Gustafson, K.R.; Cardellina, J.H.; Gray, G.N.; Boyd, M.R. Novel naphthoquinones from Conospermum incurvum. J. Nat. Prod. 1994, 57, 151–156. [Google Scholar] [CrossRef]
- Rajkumar, M.; Chandra, R.H.; Asres, K.; Veeresham, C. Toddalia asiatica (Linn.) Lam.—A Comprehensive Review. Pharmacogn. Rev. 2008, 2, 386–397. [Google Scholar]
- Babula, P.; Adam, V.; Havel, L.; Kizek, L. Noteworthy secondary metabolites naphthoquinones—Their occurrence, pharmacological properties and analysis. Curr. Pharm. Anal. 2009, 5, 47–68. [Google Scholar] [CrossRef]
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads against Protozoan Neglected Diseases—Part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, X.; LaMont, J.T.; Pardee, A.B.; Li, C.J. Selective killing of cancer cells by β-lapachone: Direct checkpoint activation as a strategy against cancer. Proc. Nat. Acad. Sci. USA 2003, 100, 2674–2678. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Sendker, J.; Nahrstedt, A. Generation of primary amide glucosides from cyanogenic glucosides. Phytochemistry 2009, 70, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Y.; Zeng, X.M.; Lu, Y.Y.; Lu, W.J.; Feng, L.L.; Yang, X.Q.; Zheng, Q.P. Senescent Leaves of Artemisia annua are One of the Most Active Organs for Overexpression of Artemisinin Biosynthesis Responsible Genes upon Burst of Singlet Oxygen. Planta Medica 2009, 76, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Binder, R.G.; Benson, M.E.; Flath, R.A. Eight 1,4-Naphthoquinones from Juglans. Phytochemistry 1989, 28, 2799–2801. [Google Scholar] [CrossRef]
- Bailey, N.J.C.; Wang, Y.; Sampson, J.; Davis, W.; Whitcombe, I.; Hylands, P.J.; Croft, S.L.; Holmes, E. Prediction of anti-plasmodial activity of Artemisia annua extracts: Application of 1H-NMR spectroscopy and chemometrics. J. Pharm. Biomed. Anal. 2004, 35, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Taketa, A.T.; Pereda-Miranda, R.; Choi, Y.H.; Verpoorte, R.; Villarreal, M.L. Metabolic Profiling of the Mexican Anxiolytic and Sedative Plant Galphimia glauca using Nuclear Magnetic Resonance Spectroscopy and Multivariate Data Analysis. Planta Med. 2008, 74, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.J.; Wang, Y.Z.; Itagaki, K.; Ma, H.X.; Xin, P.; Zhou, X.G.; Chen, G.Y.; Li, S.; Sun, S.Q. Identification of active compounds from Caesalpinia sappan L. extracts suppressing IL-6 production in RAW 264.7 cells by PLS. J. Ethnopharmacol. 2013, 148, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.M.; Luo, J.G.; Huang, F.; Kong, L.Y. Chemical characteristics combined with bioactivity for comprehensive evaluation of Panax ginseng C.A. Meyer in different ages and seasons based on HPLC-DAD and chemometric methods. J. Pharm. Biomed. Anal. 2014, 89, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Schmidt, S.; Müller-Hannemann, M.; Neumann, S. In silico fragmentation for computer assisted identification of metabolite mass spectra. BMC Bioinform. 2010, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, H.O.; Berger, S.; Braun, S. 13C-NMR-Spektroskopie, 1st ed.; Georg Thieme Verlag: Stuttgart, Germany, 1984; p. 400. [Google Scholar]
- Son, J.K. Isolation and Structure Determination of a New Tetralone Glucoside from the Roots of Juglans mandshurica. Arch. Pharm. Res. 1995, 18, 203–205. [Google Scholar] [CrossRef]
- Duroux, L.; Delmotte, F.M.; Lancelin, J.M.; Kéravis, G.; Jay-Allemand, C. Insight into naphthoquinone metabolism: β-glucosidase-catalysed hydrolysis of hydrojuglone β-d-glucopyranoside. Biochem. J. 1998, 333, 275–283. [Google Scholar] [PubMed]
- Schmidt, T.J.; Nour, A.M.; Khalid, S.A.; Kaiser, M.; Brun, R. Quantitative structure-antiprotozoal activity relationships of sequiterpene lactones. Molecules 2009, 14, 2062–2076. [Google Scholar] [CrossRef] [PubMed]
- Carrol, M.; McCrorie, P. Glycosidases in bloodstream forms of Trypanosoma brucei brucei. Comp. Biochem. Physiol. 1980, 67, 685–688. [Google Scholar] [CrossRef]
- Sample Availability: Sample of compound (1) is available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellendorff, T.; Brun, R.; Kaiser, M.; Sendker, J.; Schmidt, T.J. PLS-Prediction and Confirmation of Hydrojuglone Glucoside as the Antitrypanosomal Constituent of Juglans Spp. Molecules 2015, 20, 10082-10094. https://doi.org/10.3390/molecules200610082
Ellendorff T, Brun R, Kaiser M, Sendker J, Schmidt TJ. PLS-Prediction and Confirmation of Hydrojuglone Glucoside as the Antitrypanosomal Constituent of Juglans Spp. Molecules. 2015; 20(6):10082-10094. https://doi.org/10.3390/molecules200610082
Chicago/Turabian StyleEllendorff, Therese, Reto Brun, Marcel Kaiser, Jandirk Sendker, and Thomas J. Schmidt. 2015. "PLS-Prediction and Confirmation of Hydrojuglone Glucoside as the Antitrypanosomal Constituent of Juglans Spp." Molecules 20, no. 6: 10082-10094. https://doi.org/10.3390/molecules200610082