
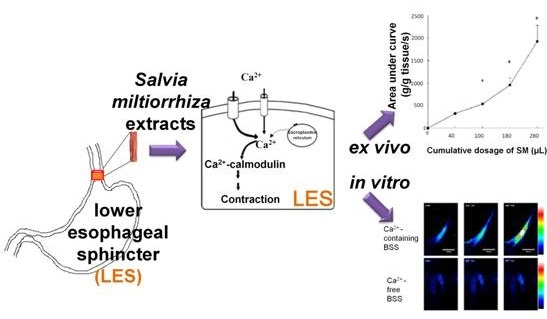
Salvia miltiorrhiza Induces Tonic Contraction of the Lower Esophageal Sphincter in Rats via Activation of Extracellular Ca2+ Influx

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. The Content of Tanshinone IIA (TA) in SM Extracts
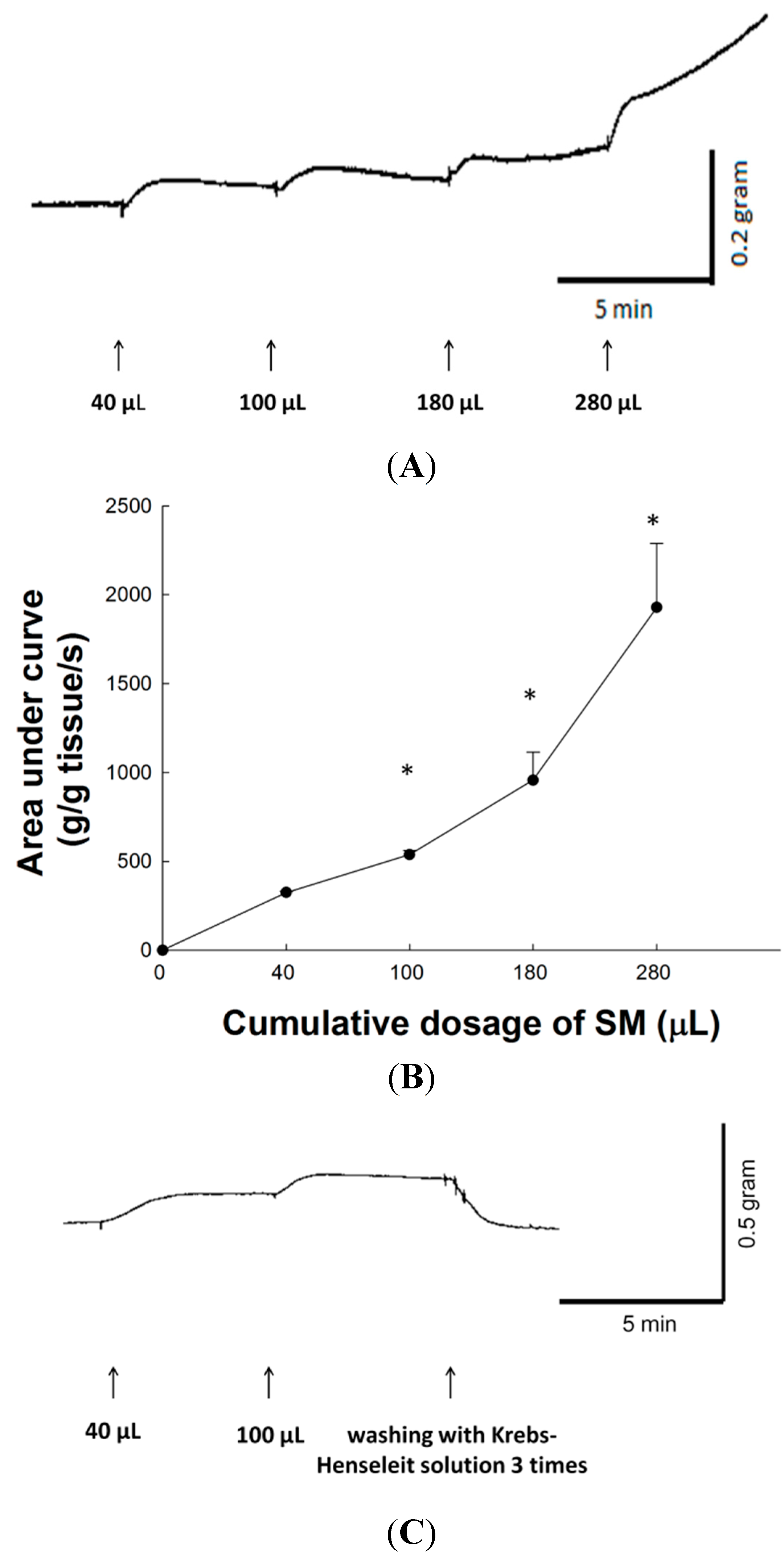
2.2. Dose-Response Effect of SM Extracts on Contraction of LES

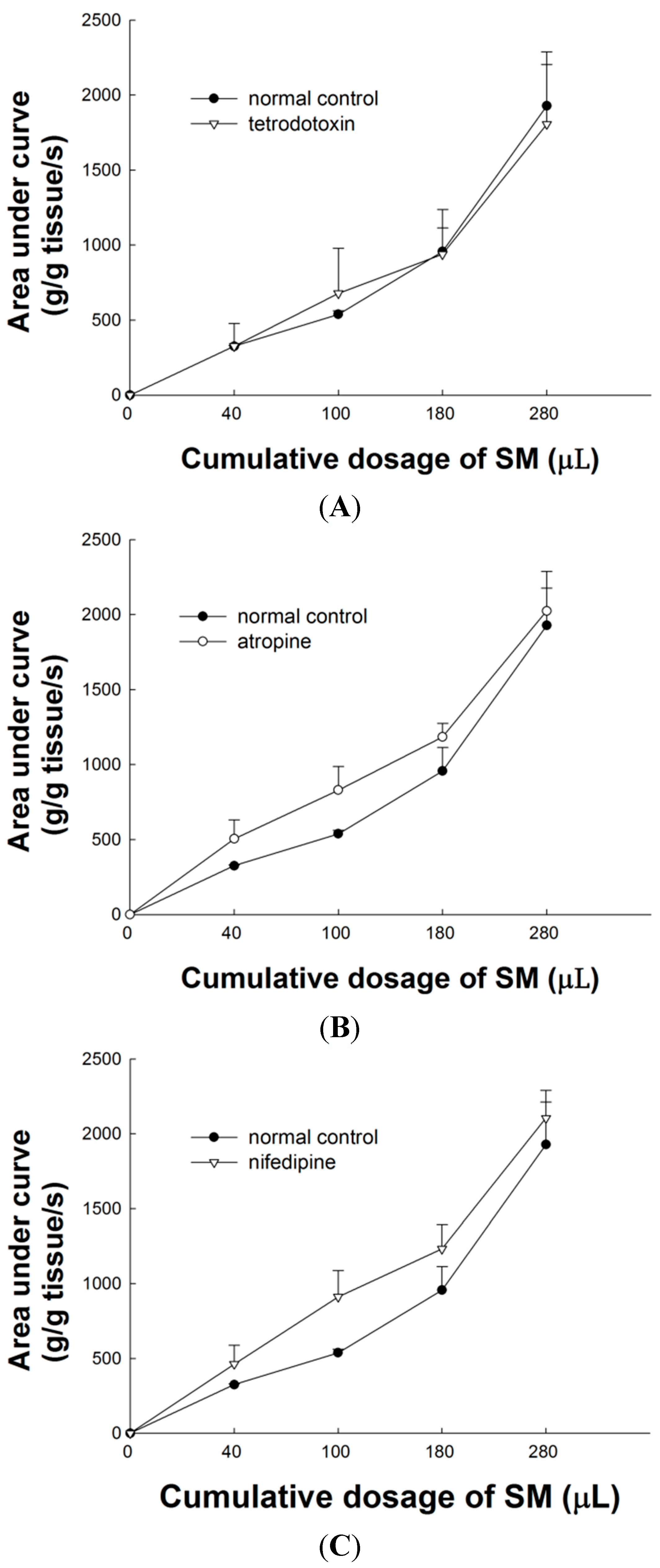
2.3. Effect of Tetrodotoxin (TTX) on SM Extracts-Induced Contraction of LES
2.4. Effect of Atropine on SM Extract-Induced Contraction of LES

2.5. Effect of L-Type Ca2+ Channel Blocker on SM Extract-Induced Contraction of LES
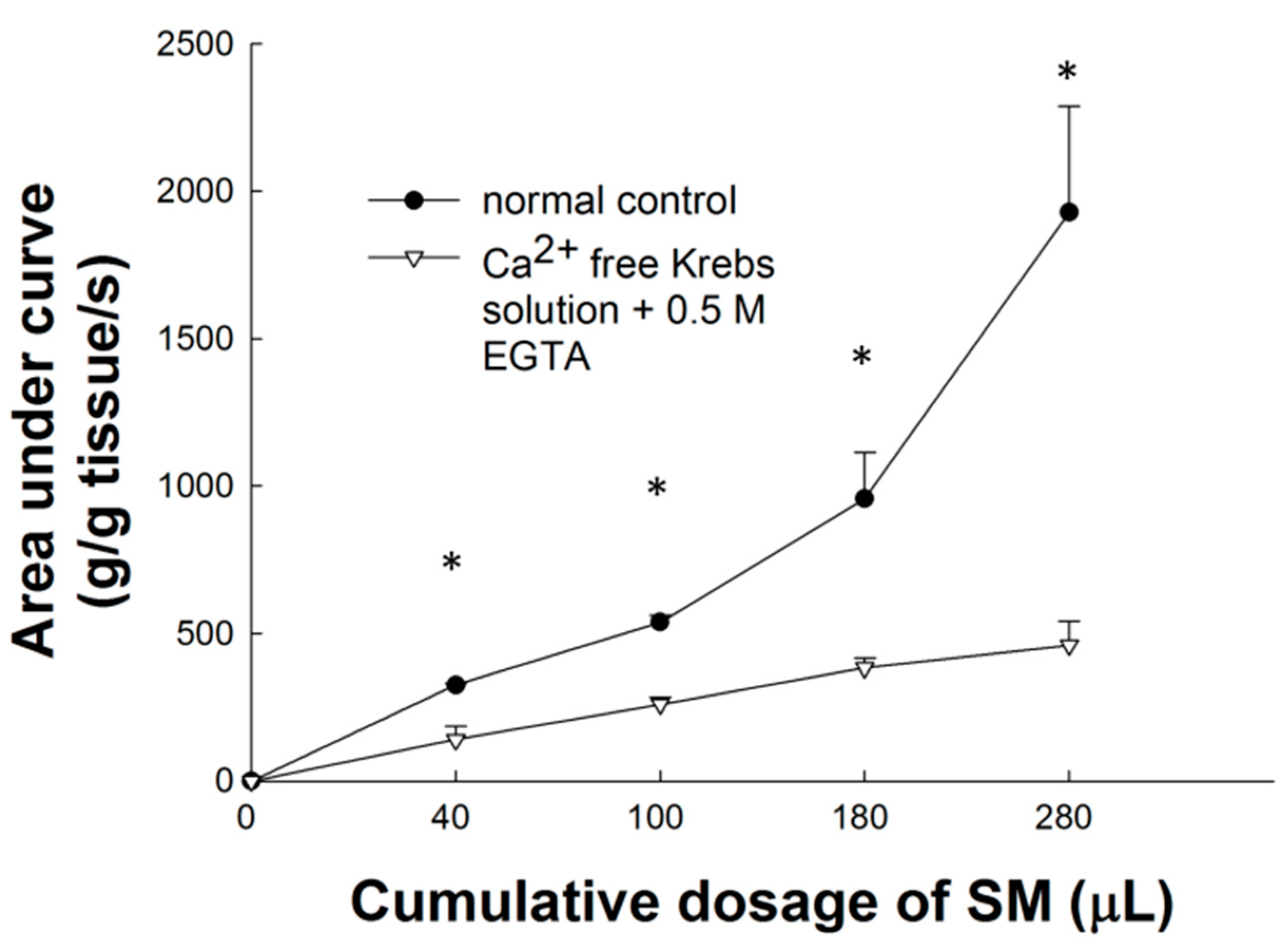
2.6. Effect of Ca2+-Free Krebs-Henseleit Solution plus EGTA on SM Extract-Induced Contraction of LES

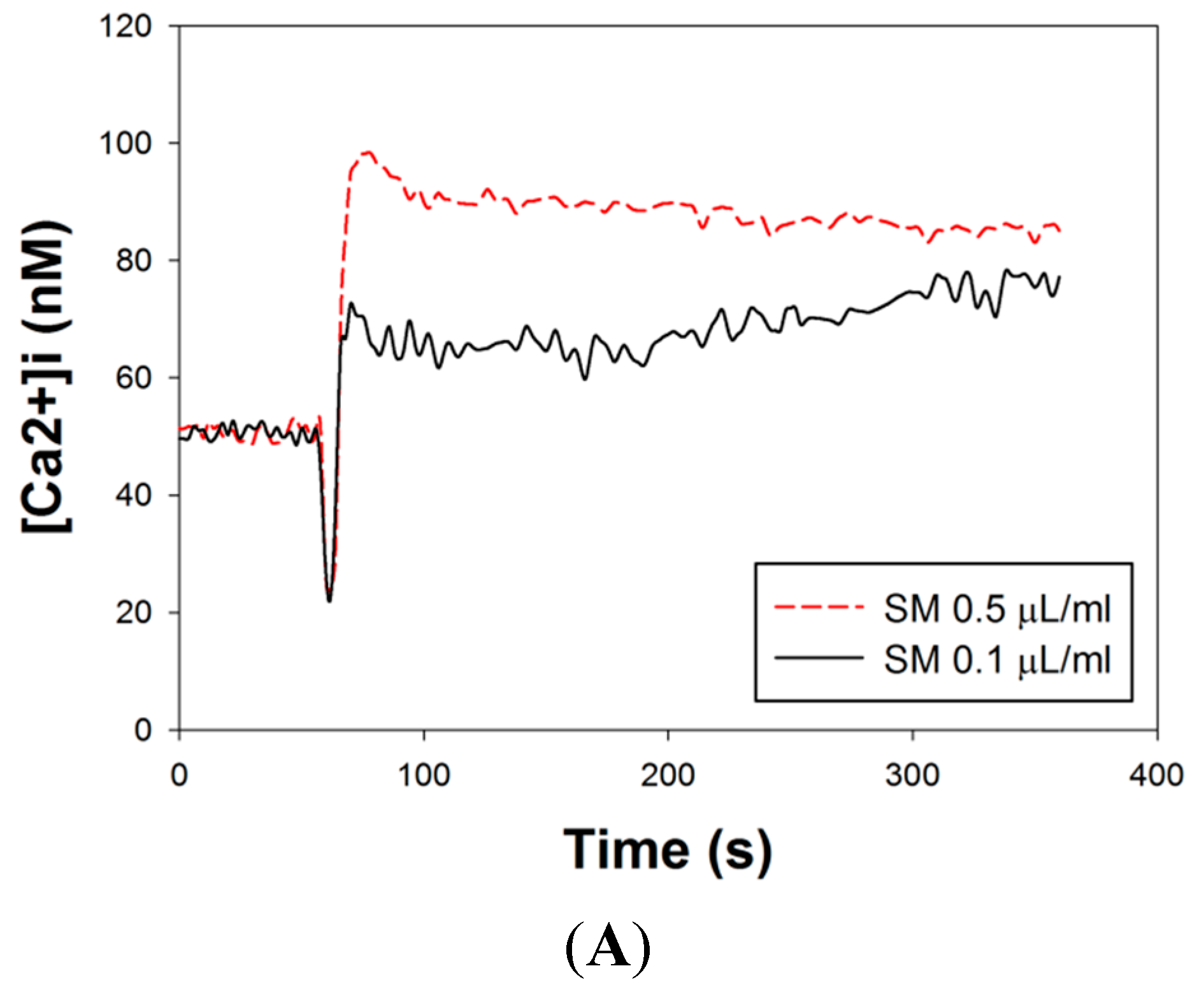
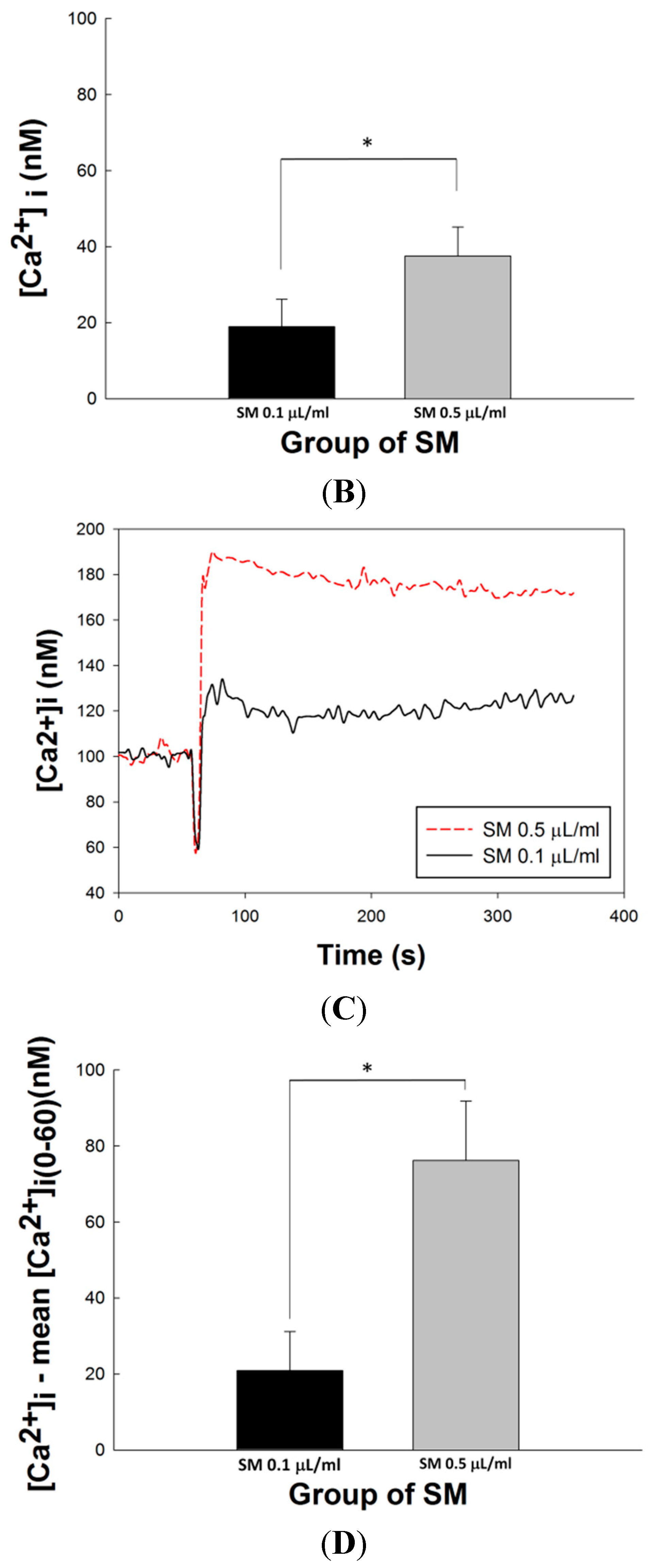
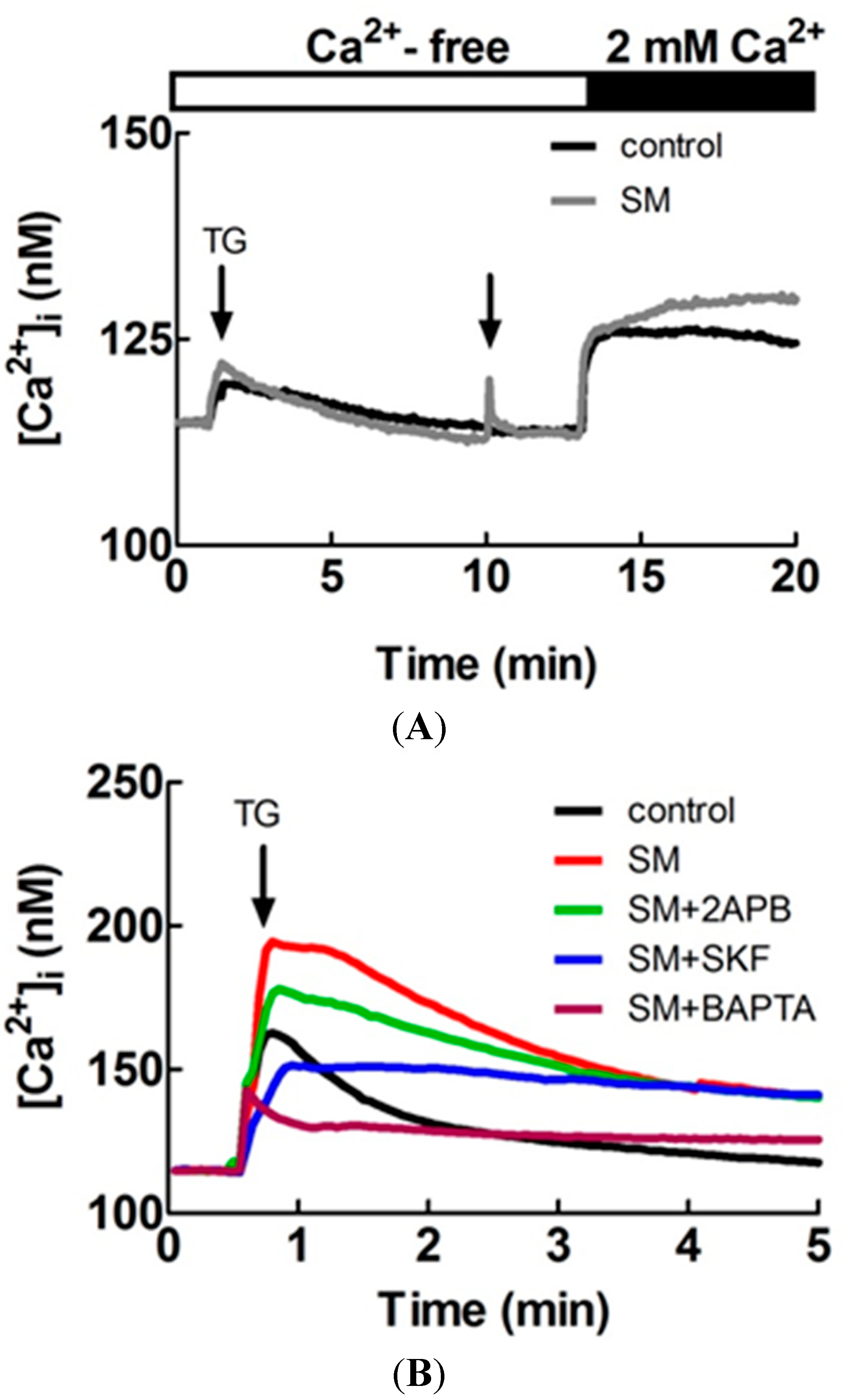
2.7. Effects of SM Extracts on Cytosolic Concentration of Free Ca2+ ([Ca2+]i) in Primary LES Smooth Muscle Cells


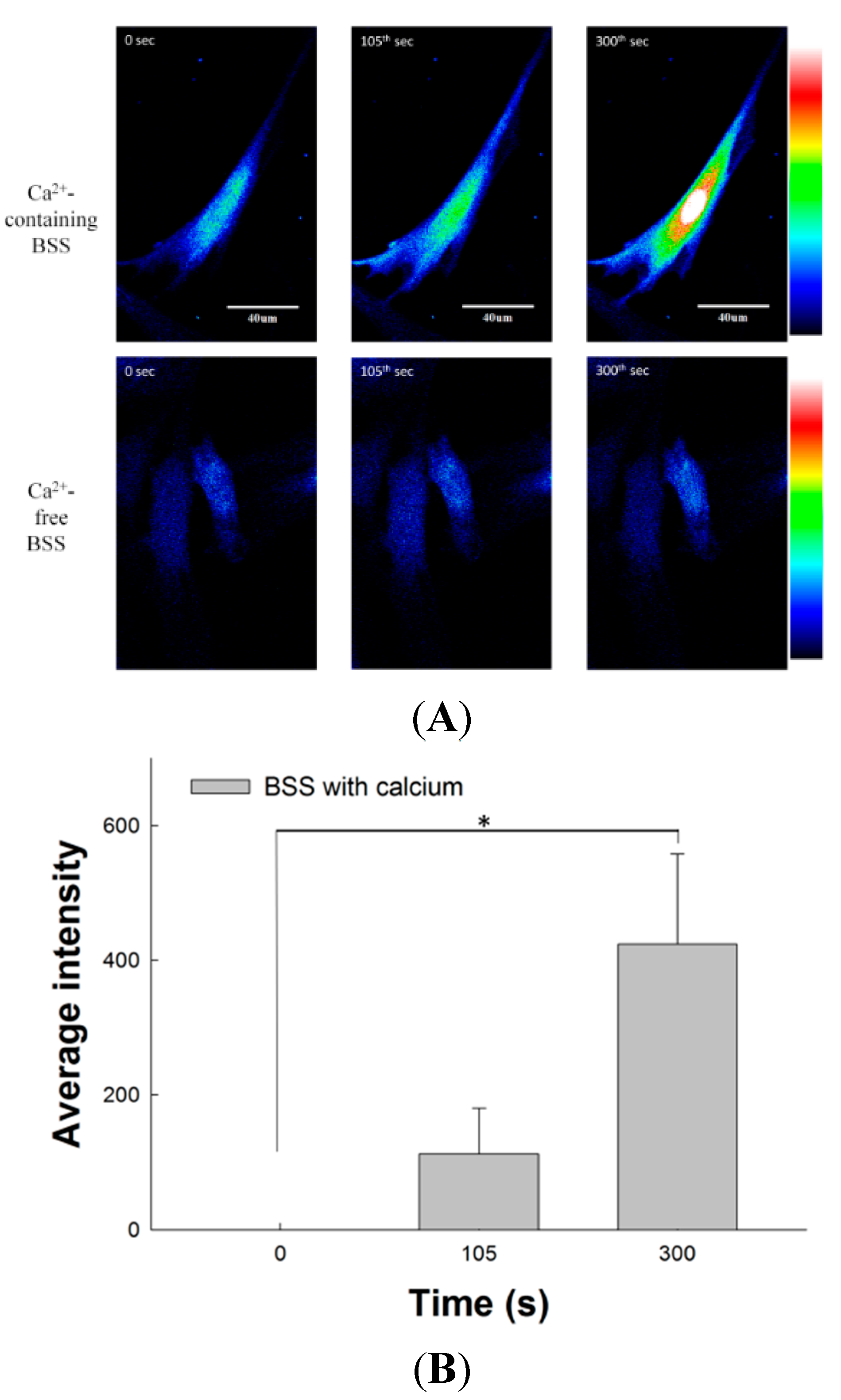
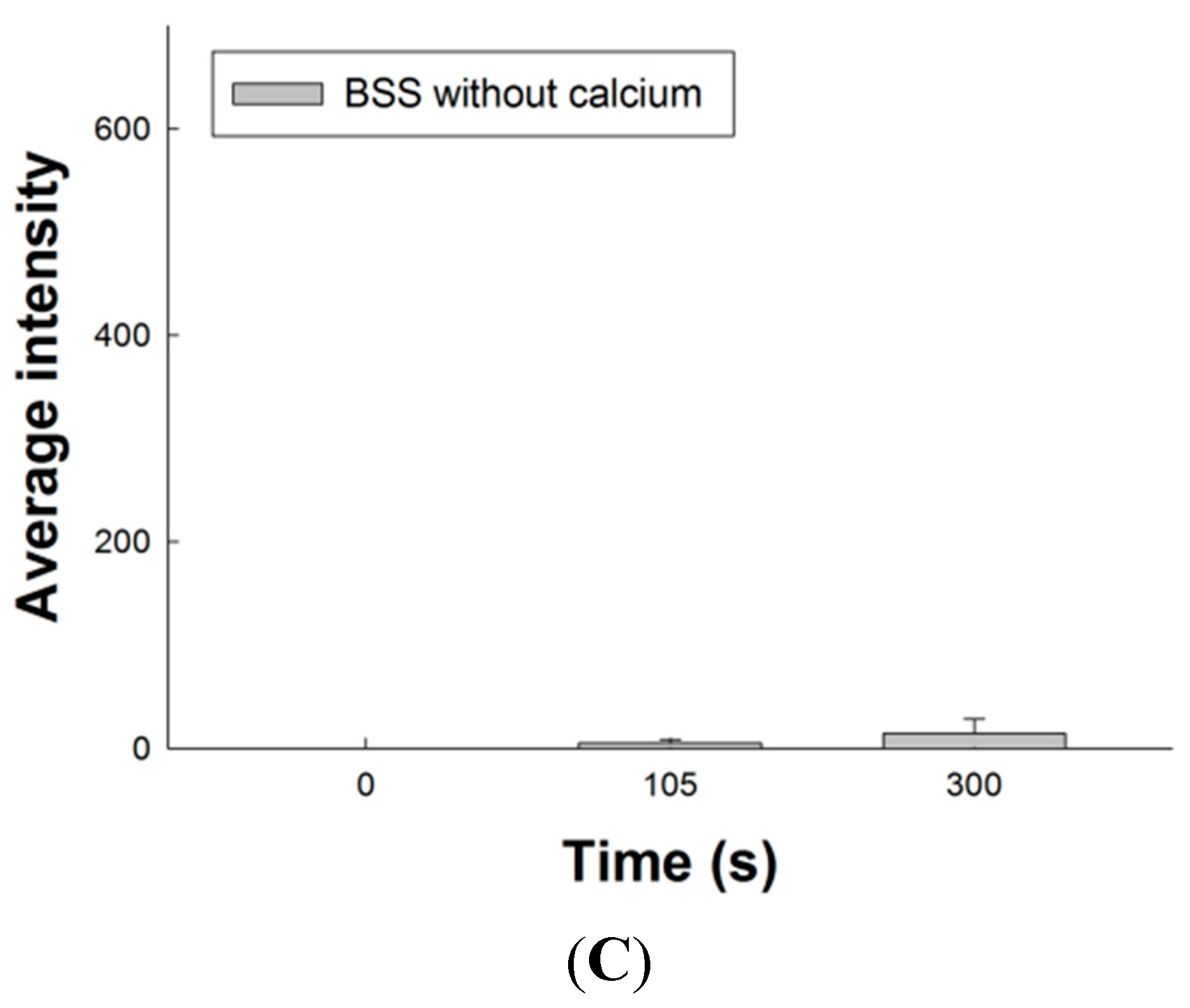
2.8. Effect of SM Extracts on Confocal Images of Changes in Intracellular [Ca2+] in Primary LES Smooth Muscle Cells



3. Experimental Section
3.1. Materials
3.2. Preparation of SM Extracts
3.3. HPLC Analysis of TA in SM Extracts
3.4. Measurement of Contraction of LES in Rat
3.5. Effect of Tetrodotoxin, Atropine, Nifedipine and Ca2+-Free Krebs-Henseleit Solution with EGTA on SM Extract-Induced Contraction of LES in Rat
3.6. Enzymatic Isolation of Primary LES Smooth Muscle Cells
3.7. [Ca2+]i Measurements
3.8. Confocal Microscope Fluorescent Images
3.9. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boeckxstaens, G.E. The lower oesophageal sphincter. Neurogastroenterol. Motil. 2005, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Sifrim, D.; Zerbib, F. Diagnosis and management of patients with reflux symptoms refractory to proton pump inhibitors. Gut 2012, 61, 1340–1354. [Google Scholar] [CrossRef] [PubMed]
- Bredenoord, A.J.; Pandolfino, J.E.; Smout, A.J. Gastro-oesophageal reflux disease. Lancet 2013, 381, 1933–1942. [Google Scholar] [CrossRef]
- Yasawy, M.I.; Randhawa, M.A. GERD is becoming a challenge for the medical profession: Is there any remedy? Hepatogastroenterology 2014, 61, 1623–1626. [Google Scholar] [PubMed]
- Pharmacopoeia Commission of People’s Republic of China. Pharmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2010; p. 71. [Google Scholar]
- Wang, X.; Morris-Natschke, S.L.; Lee, K.H. New developments in the chemistry and biology of the bioactive constituents of Tanshen. Med. Res. Rev. 2007, 27, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Chen, J.M.; Peng, Y.; Wu, Q.; Xiao, P.G. Investigation of Danshen and related medicinal plants in China. J. Ethnopharmacol. 2008, 120, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.M.; Xia, Q.; Zhang, X.; Xu, W.H.; Jiang, H.D.; Chen, J.Z. Salvia miltiorrhiza attenuates the changes in contraction and intracellular calcium induced by anoxia and reoxygenation in rat cardiomyocytes. Life Sci. 2003, 72, 2451–2463. [Google Scholar] [CrossRef]
- Lam, F.F.; Deng, S.Y.; Ng, E.S.; Yeung, J.H.; Kwan, Y.W.; Lau, C.B.; Koon, J.C.; Zhou, L.; Zuo, Z.; Leung, P.C.; et al. Mechanisms of the relaxant effect of a Danshen and Gegen formulation on rat isolated cerebral basilar artery. J. Ethnopharmacol. 2010, 132, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.; Chui, S.H.; Wong, D.Y.; Ha, W.Y.; Chan, C.L.; Wong, R.N. Protective effects of Danshensu from the aqueous extract of Salvia miltiorrhiza (Danshen) against homocysteine-induced endothelial dysfunction. Life Sci. 2004, 75, 3157–3171. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, Y.; Xue, L.; Severino, R.P.; Gao, S.; Niu, J.; Qin, L.P.; Zhang, D.; Brömme, D. Salvia miltiorrhiza: An ancient Chinese herbal medicine as a source for anti-osteoporotic drugs. J. Ethnopharmacol. 2014, 155, 1401–1416. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, J.; Bao, J.; Lu, J.; Wang, Y. The anticancer properties of Salvia miltiorrhiza Bunge (Danshen): A systematic review. Med. Res. Rev. 2014, 34, 768–794. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Chen, X.P.; Lu, Q.P. Effect of Salvia miltiorrhiza pretreatment on the CCK and VIP expression in hepatic ischemia-reperfusion-induced digestive tract congestion. Front. Med. China 2010, 4, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.D.; Wang, C.Z.; Yu, C.; Zhang, Z.; Calway, T.; Wang, Y.; Li, P.; Yuan, C.S. Salvia miltiorrhiza (Danshen) significantly ameliorates colon inflammation in dextran sulfate sodium induced colitis. Am. J. Chin. Med. 2013, 41, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Xiping, Z.; Yan, P.; Xinmei, H.; Guanghua, F.; Meili, M.; Jie, N.; Fangjie, Z. Effects of dexamethasone and Salvia miltiorrhizae on the small intestine and immune organs of rats with severe acute pancreatitis. Inflammation 2010, 33, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Huang, S.C.; Liu, J.K.; Wang, H.C.; Tsai, T.R.; Tsai, P.J.; Liu, C.W.; Chang, L.C. Salvia miltiorrhiza causes tonic contraction in rat ileum through Ca2+-calmodulin pathway. J. Ethnopharmacol. 2012, 142, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Farré, R.; Sifrim, D. Regulation of basal tone, relaxation and contraction of the lower oesophageal sphincter. Relevance to drug discovery for oesophageal disorders. Br. J. Pharmacol. 2008, 153, 858–869. [Google Scholar] [CrossRef] [PubMed]
- Muinuddin, A.; Neshatian, L.; Gaisano, H.Y.; Diamant, N.E. Calcium source diversity in feline lower esophageal sphincter circular and sling muscle. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Biancani, P.; Sohn, U.D.; Rich, H.G.; Harnett, K.M.; Behar, J. Signal transduction pathways in esophageal and lower esophageal sphincter circular muscle. Am. J. Med. 1997, 103, 23S–28S. [Google Scholar] [CrossRef]
- Hornby, P.J.; Abrahams, T.P. Central control of lower esophageal sphincter relaxation. Am. J. Med. 2000, 108, 90–98. [Google Scholar] [CrossRef]
- Wang, J.; Laurier, L.G.; Sims, S.M.; Preiksaitis, H.G. Enhanced capacitative calcium entry and TRPC channel gene expression in human LES smooth muscle. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284. [Google Scholar] [CrossRef] [PubMed]
- Sims, S.M.; Jiao, Y.; Preiksaitis, H.G. Regulation of intracellular calcium in human esophageal smooth muscle. Am. J. Physiol. 1997, 273, 1679–1689. [Google Scholar]
- Sanders, K.M. Signal transduction in smooth muscle: Mechanisms of calcium handling in smooth muscles. J. Appl. Physiol. 2001, 91, 1438–1449. [Google Scholar] [PubMed]
- Zhonghua Bencao Committee of State Traditional Chinese Medicine Administration. Zhonghua Bencao; Shanghai Scientific and Technical Press: Shanghai, China, 1999; pp. 169–186. [Google Scholar]
- Pan, X.; Niu, G.; Liu, H. Microwave-assisted extraction of tanshinones from Salvia miltiorrhiza bunge with analysis by high-performance liquid chromatography. J. Chromatogr. A 2001, 922, 371–375. [Google Scholar] [CrossRef]
- Chang, L.C.; Wu, C.L.; Liu, C.W.; Chuo, W.H.; Li, P.C.; Tsai, T.R. Preparation, characterization and cytotoxicity evaluation of tanshinone IIA nanoemulsions. J. Biomed. Nanotechnol. 2011, 7, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.C. Dendroaspis natriuretic peptide is the most potent natriuretic peptide to cause relaxation of lower esophageal sphincter. Regul. Pept. 2011, 167, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Nagao, M.; Linden, D.R.; Duenes, J.A.; Sarr, M.G. Mechanisms of action of the gasotransmitter hydrogen sulfide in modulating contractile activity of longitudinal muscle of rat ileum. J. Gastrointest. Surg. 2011, 15, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.S.; Chang, J.C.; Wang, Y.S.; Huang, S.C. Cysteinyl leucotriene receptor type 1 mediates contraction in human and guinea-pig oesophagus. Neurogastroenterol. Motil. 2008, 20, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Je, H.D.; Shim, J.H.; Sohn, U.D. Characteristics of spontaneous contraction in the circular smooth muscles of cat ileum. Arch. Pharm. Res. 2010, 33, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Cruz, F.; Paterson, W.G. Evidence for altered circular smooth muscle cell function in lower esophageal sphincter of W/Wv mutant mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [PubMed]
- Ohta, T.; Ito, S.; Nakazato, Y. Chloride currents activated by caffeine in rat intestinal smooth muscle cells. J. Physiol. 1993, 465, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Zohar, O.; Ikeda, M.; Shinagawa, H.; Inoue, H.; Nakamura, H.; Elbaum, D.; Alkon, D.L.; Yoshioka, T. Thermal imaging of receptor-activated heat production in single cells. Biophys. J. 1998, 74, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Florea, A.M.; Splettstoesser, F.; Büsselberg, D. Arsenic trioxide (As2O3) induced calcium signals and cytotoxicity in two human cell lines: SY-5Y neuroblastoma and 293 embryonic kidney (HEK). Toxicol. Appl. Pharmacol. 2007, 220, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples in the present paper are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, C.-C.; Chang, L.-C.; Huang, S.-C.; Tey, S.-L.; Hsu, W.-L.; Su, Y.-T.; Liu, C.-W.; Tsai, T.-R. Salvia miltiorrhiza Induces Tonic Contraction of the Lower Esophageal Sphincter in Rats via Activation of Extracellular Ca2+ Influx. Molecules 2015, 20, 14504-14521. https://doi.org/10.3390/molecules200814504
Tsai C-C, Chang L-C, Huang S-C, Tey S-L, Hsu W-L, Su Y-T, Liu C-W, Tsai T-R. Salvia miltiorrhiza Induces Tonic Contraction of the Lower Esophageal Sphincter in Rats via Activation of Extracellular Ca2+ Influx. Molecules. 2015; 20(8):14504-14521. https://doi.org/10.3390/molecules200814504
Chicago/Turabian StyleTsai, Ching-Chung, Li-Ching Chang, Shih-Che Huang, Shu-Leei Tey, Wen-Li Hsu, Yu-Tsun Su, Ching-Wen Liu, and Tong-Rong Tsai. 2015. "Salvia miltiorrhiza Induces Tonic Contraction of the Lower Esophageal Sphincter in Rats via Activation of Extracellular Ca2+ Influx" Molecules 20, no. 8: 14504-14521. https://doi.org/10.3390/molecules200814504