
Bioassay-Guided Fractionation of a Leaf Extract from Combretum mucronatum with Anthelmintic Activity: Oligomeric Procyanidins as the Active Principle

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Characterization of a Hydroethanolic Leaf Extract from C. mucronatum





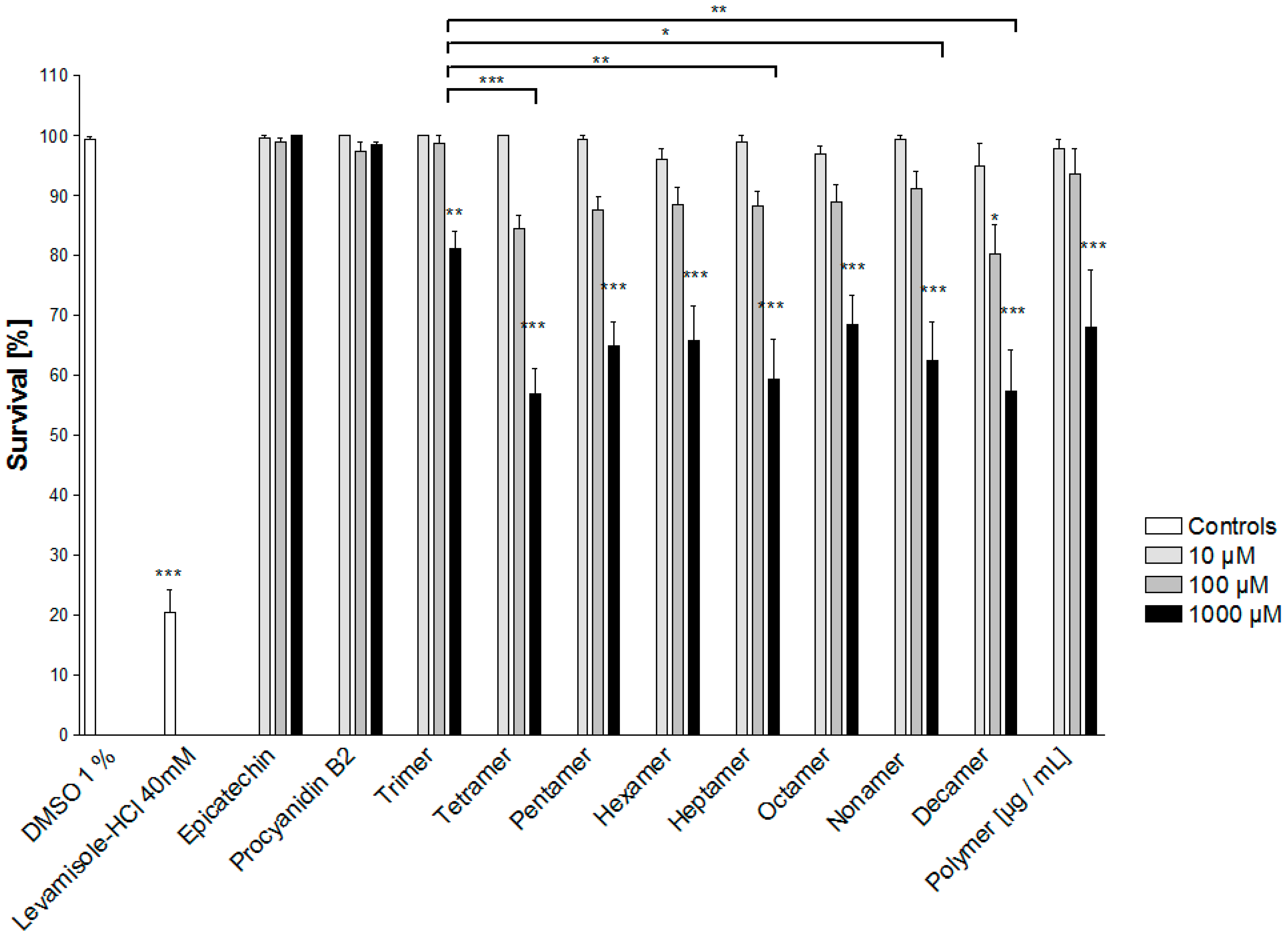
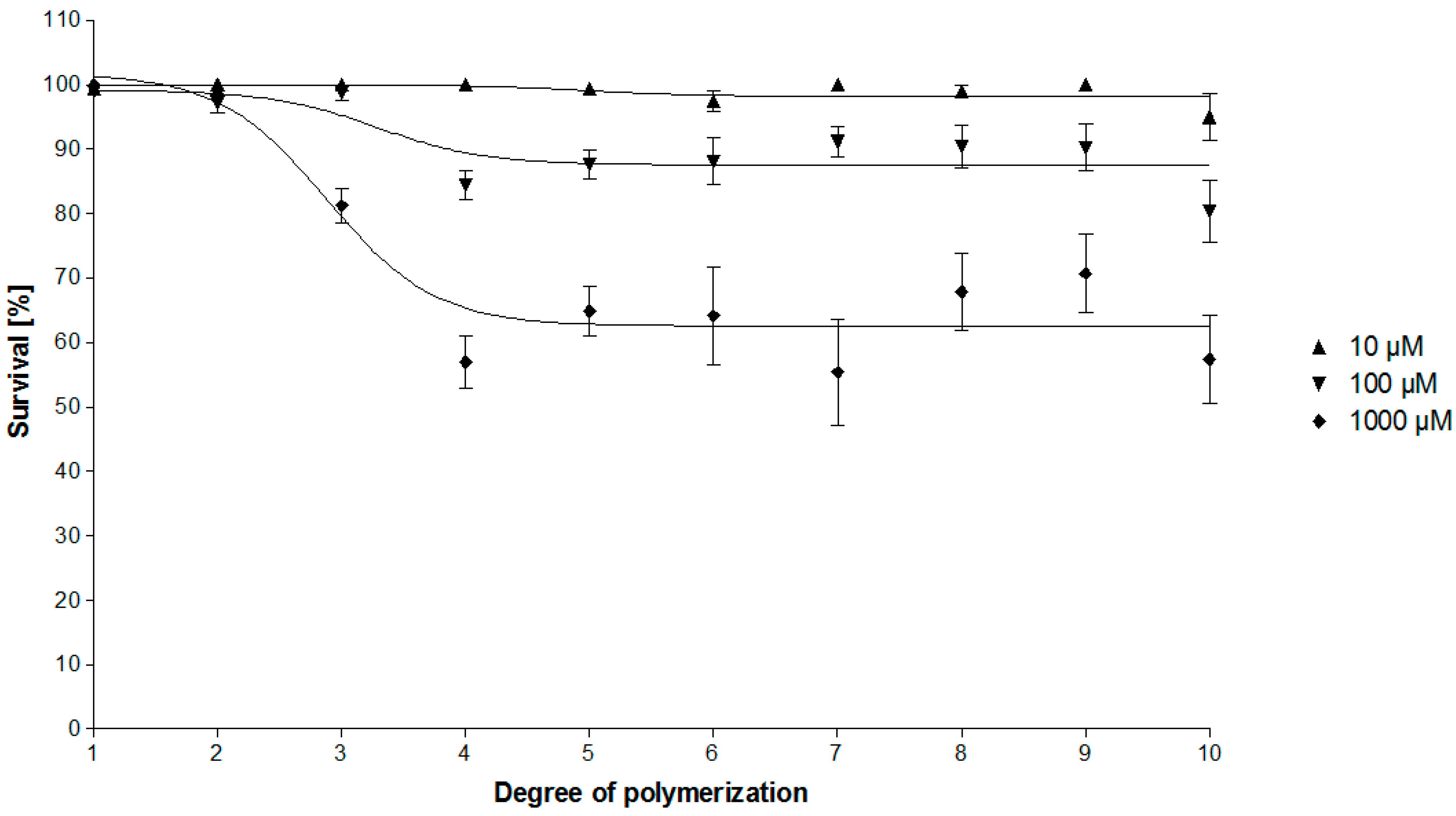
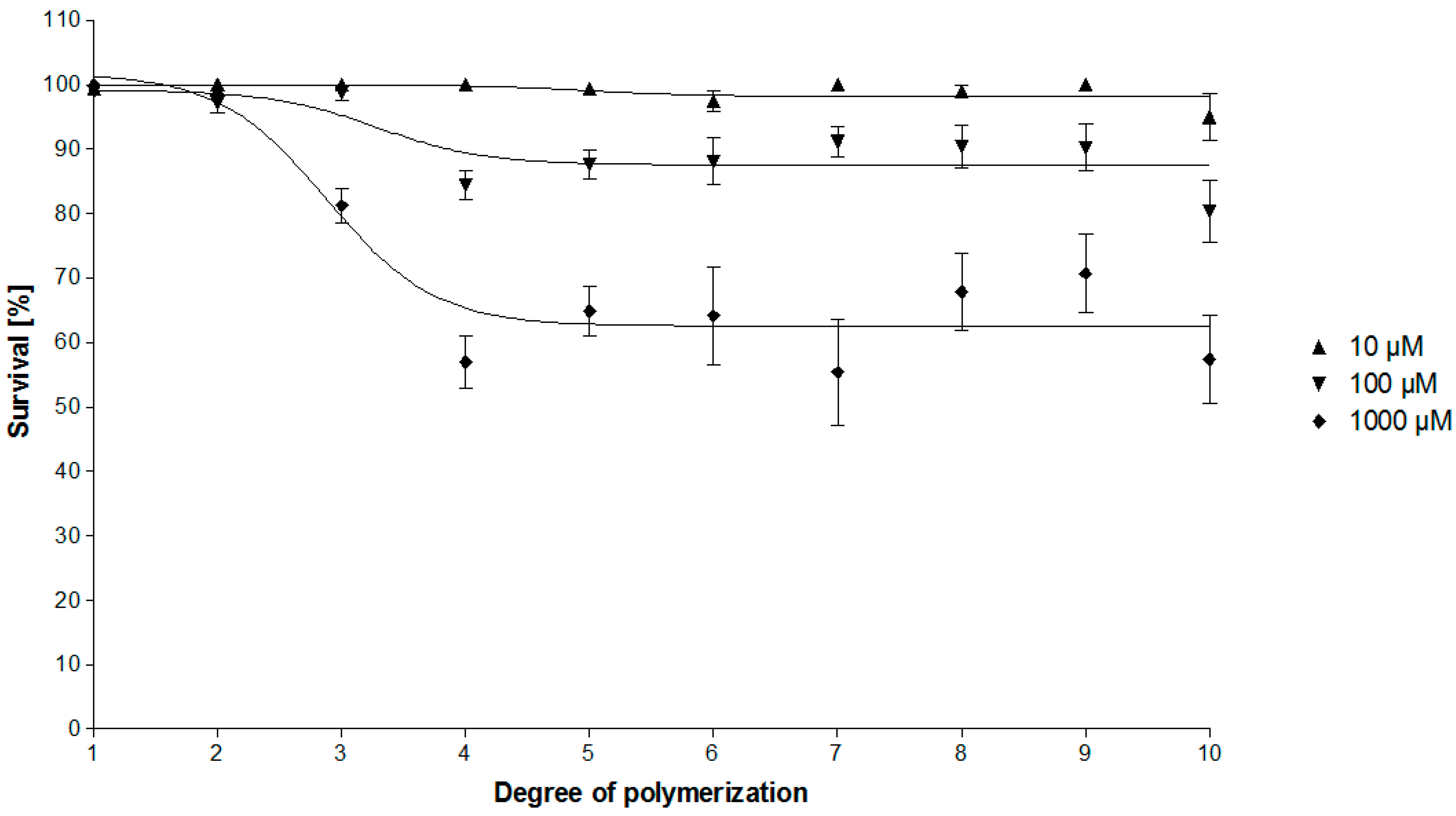
2.2. Bioassay-Guided Fractionation

3. Experimental Section
3.1. Plant Material and Chemicals
3.2. General Analytical Techniques
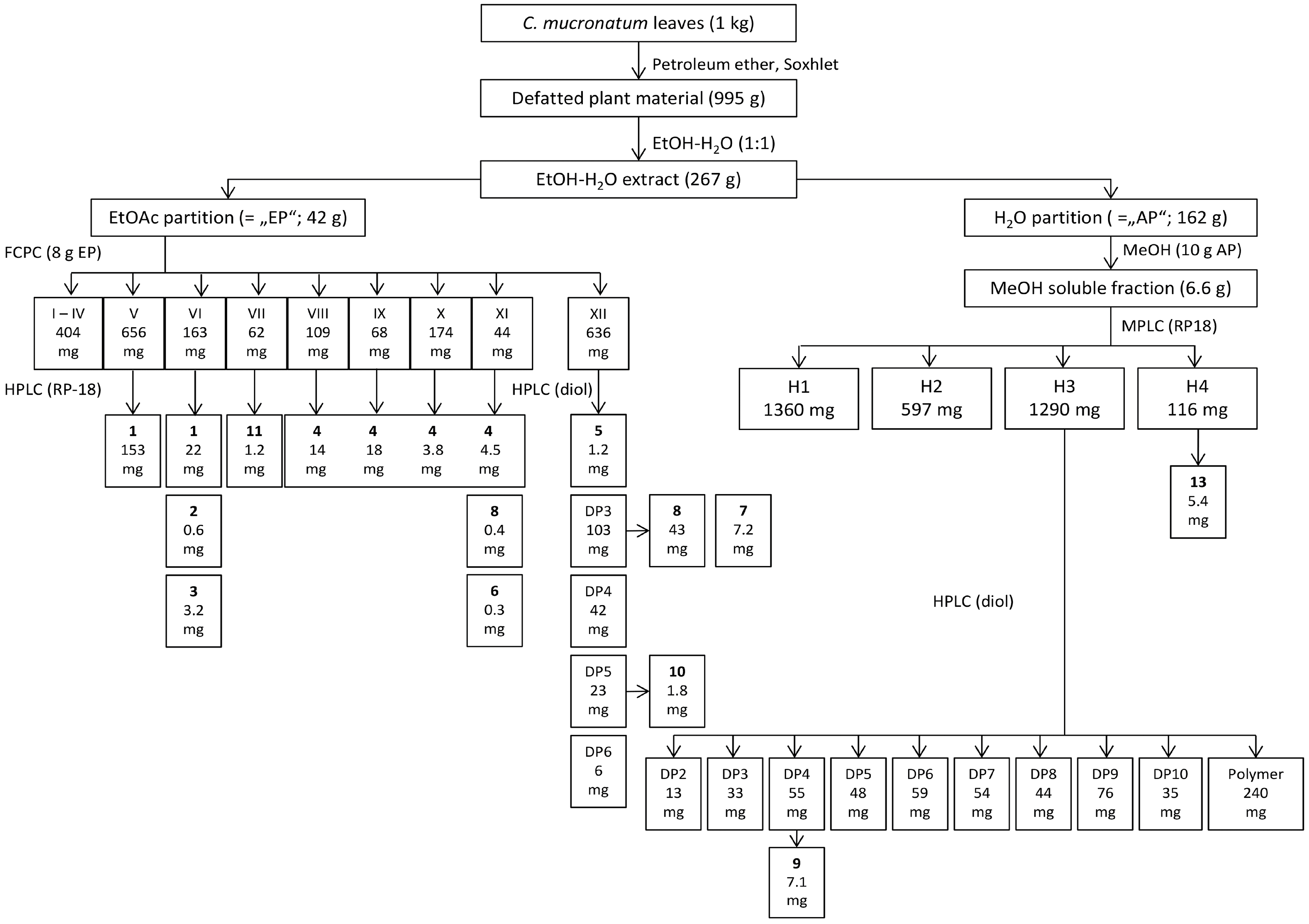
3.3. Preparation of Plant Extract and Partitions (see Figure 1)
3.4. Isolation of Oligomeric Procyanidins with DP 1 to 3
3.5. Isolation of OPC with Higher DP (DP > 3)
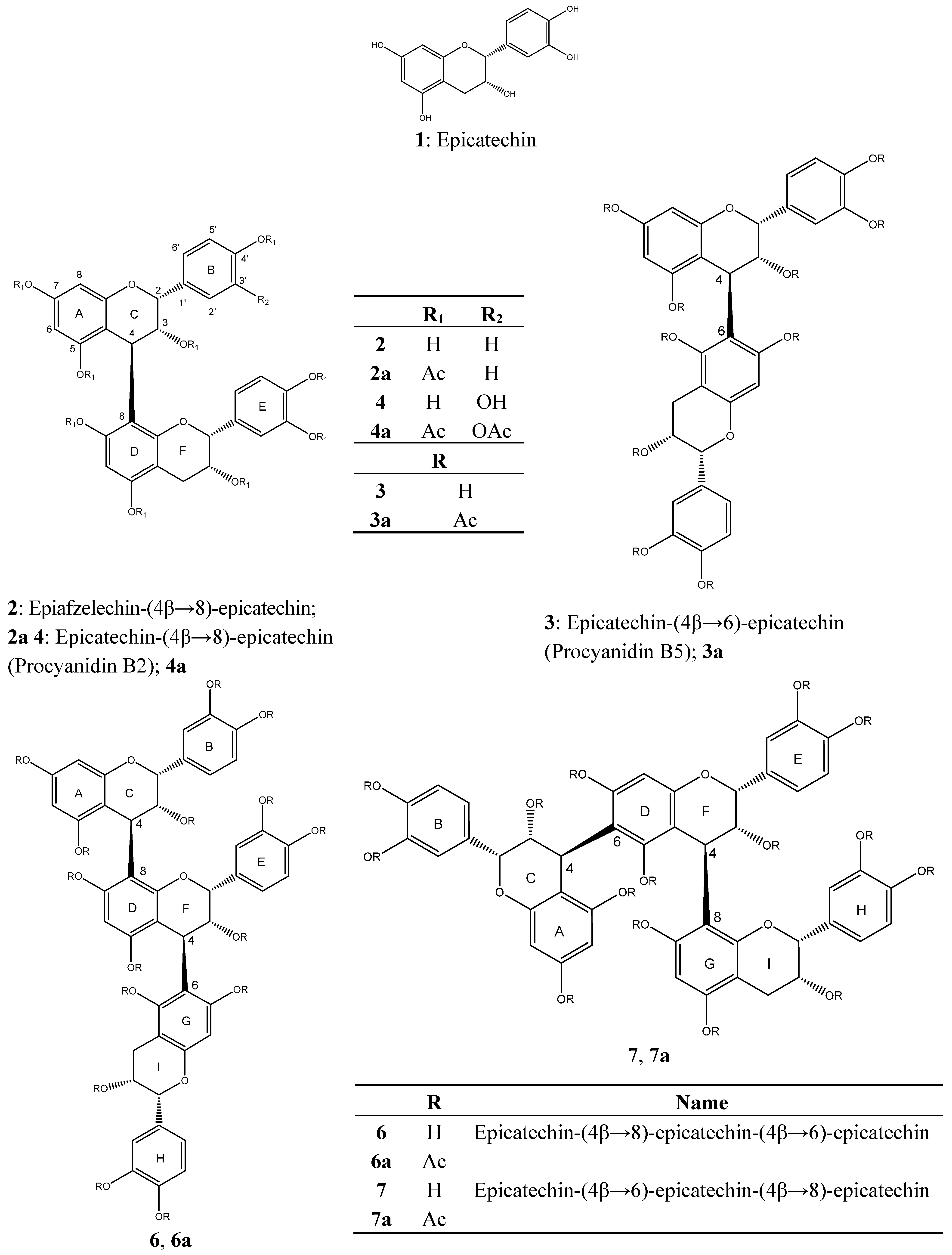
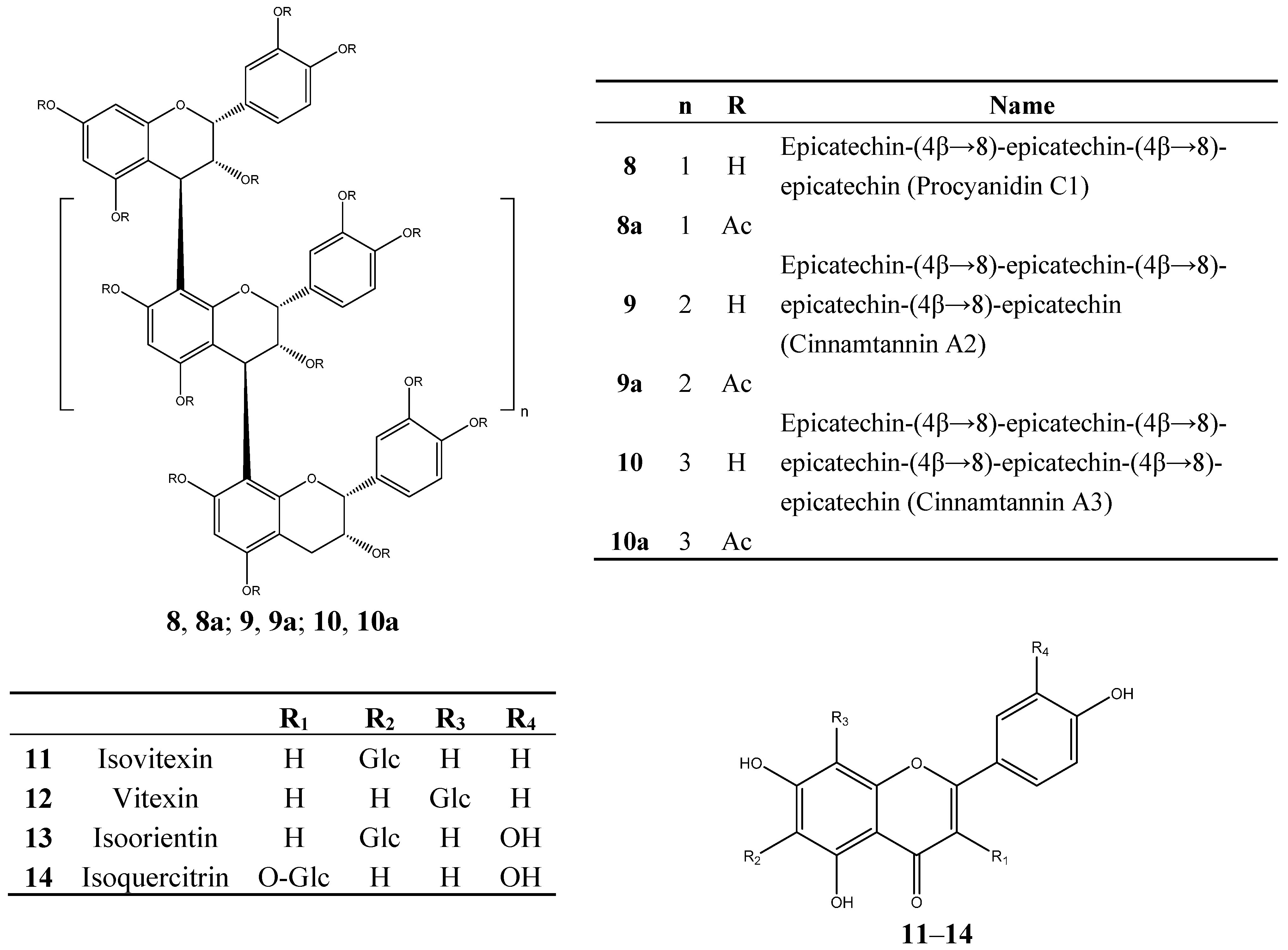
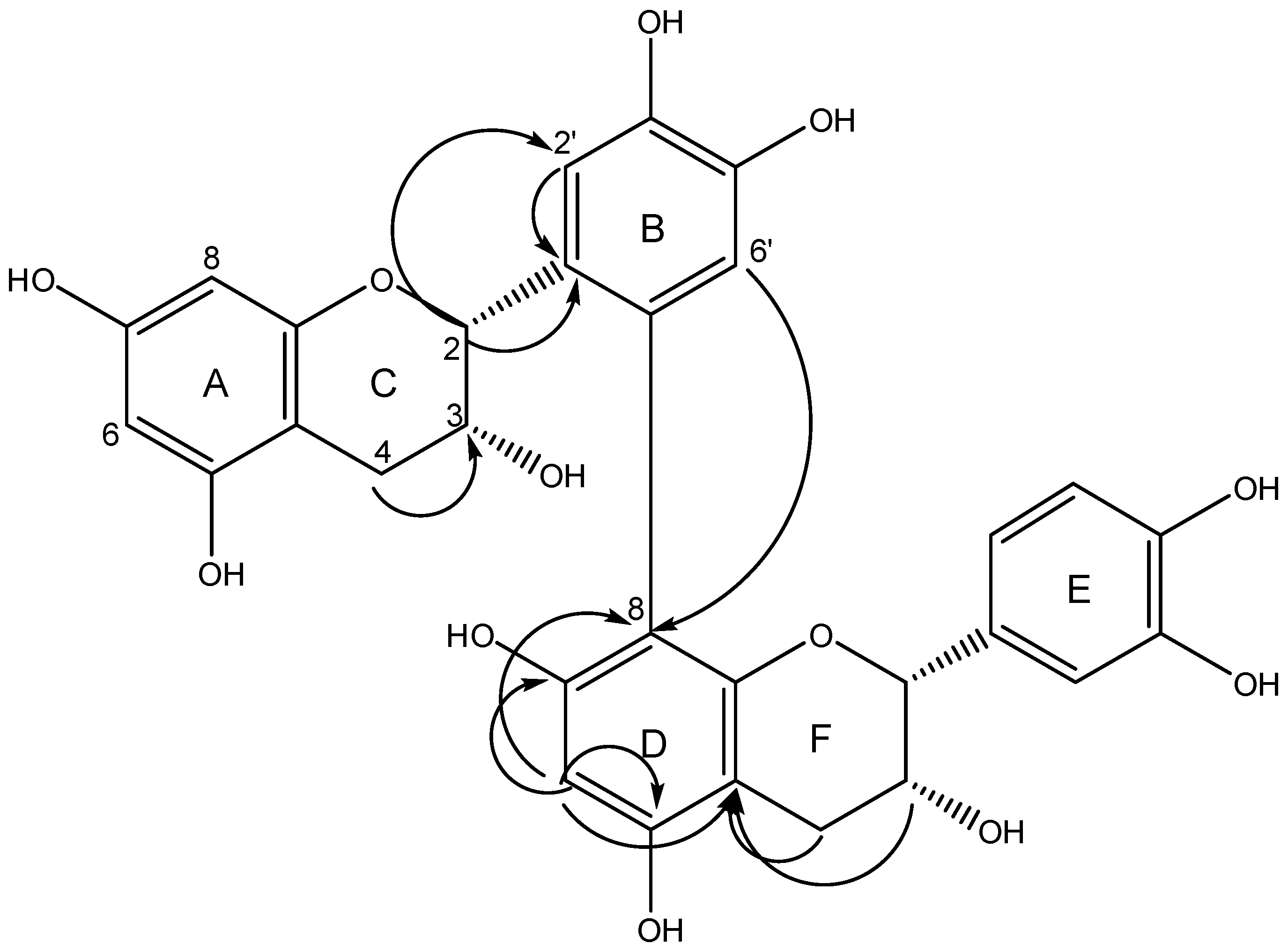
3.6. Identification of Isolated Compounds
3.7. Tannin-Depleted Extract: Precipitation of Proanthocyanidins with PVPP
3.8. Identification of Flavonoids by HPLC
3.9. Anthelmintic Assay
3.9.1. Monoxenic and Axenic Maintenance of Caenorhabditis Elegans
3.9.2. In Vitro Screening
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- World Health Organization. Soil-Transmitted Helminth Infections. Available online: http://www.who.int/mediacentre/factsheets/fs366/en/ (accessed on 26 January 2015).
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Lustigman, S.; Prichard, R.K.; Gazzinelli, A.; Grant, W.N.; Boatin, B.A.; McCarthy, J.S.; Basanez, M.G. A research agenda for helminth diseases of humans: The problem of helminthiases. PLoS Negl. Trop. Dis. 2012, 6, e1582. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Brindley, P.J.; Bethony, J.M.; King, C.H.; Pearce, E.J.; Jacobson, J. Helminth infections: The great neglected tropical diseases. J. Clin. Investig. 2008, 118, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Humphries, D.; Nguyen, S.; Boakye, D.; Wilson, M.; Cappello, M. The promise and pitfalls of mass drug administration to control intestinal helminth infections. Curr. Opin. Infect. Dis. 2012, 25, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Keiser, J.; Utzinger, J. Efficacy of current drugs against soil-transmitted helminth infections: Systematic review and meta-analysis. J. Am. Med. Assoc. 2008, 299, 1937–1948. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, J.; Albonico, M.; Behnke, J.M.; Kotze, A.C.; Prichard, R.K.; McCarthy, J.S.; Montresor, A.; Levecke, B. Is anthelmintic resistance a concern for the control of human soil-transmitted helminths? Int. J. Parasitol. Drugs Drug Resist. 2011, 1, 14–27. [Google Scholar] [CrossRef] [PubMed]
- WHO. 18th WHO Model List of Essential Medicines; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Vercruysse, J.; Levecke, B.; Prichard, R. Human soil-transmitted helminths: Implications of mass drug administration. Curr. Opin. Infect. Dis. 2012, 25, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Fabricant, D.S.; Farnsworth, N.R. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect. 2001, 109 (Suppl. 1), 69–75. [Google Scholar] [CrossRef] [PubMed]
- Ndjonka, D.; Rapado, L.N.; Silber, A.M.; Liebau, E.; Wrenger, C. Natural products as a source for treating neglected parasitic diseases. Int. J. Mol. Sci. 2013, 14, 3395–3439. [Google Scholar] [CrossRef] [PubMed]
- Agyare, C.; Spiegler, V.; Sarkodie, H.; Asase, A.; Liebau, E.; Hensel, A. An ethnopharmacological survey and in vitro confirmation of the ethnopharmacological use of medicinal plants as anthelmintic remedies in the Ashanti region, in the central part of Ghana. J. Ethnopharmacol. 2014, 158PA, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Kone, W.M.; Vargas, M.; Keiser, J. Anthelmintic activity of medicinal plants used in Côte d'Ivoire for treating parasitic diseases. Parasitol. Res. 2012, 110, 2351–2362. [Google Scholar] [CrossRef] [PubMed]
- Kisseih, E.; Lechtenberg, M.; Petereit, F.; Sendker, J.; Brandt, S.; Agyare, C.; Hensel, A. Phytochemical characterization and in vitro wound healing activity of leaf extracts from Combretum mucronatum Schum. & Thonn.: Oligomeric procyanidins as strong inductors of cellular differentiation. J. Ethnopharmacol. 2015. [Google Scholar] [CrossRef]
- McGaw, L.J.; Rabe, T.; Sparg, S.G.; Jager, A.K.; Eloff, J.N.; van Staden, J. An investigation on the biological activity of Combretum species. J. Ethnopharmacol. 2001, 75, 45–50. [Google Scholar] [CrossRef]
- Weller, M.G. A unifying review of bioassay-guided fractionation, effect-directed analysis and related techniques. Sensors (Basel) 2012, 12, 9181–9209. [Google Scholar] [CrossRef] [PubMed]
- Bicker, J.; Petereit, F.; Hensel, A. Proanthocyanidins and a phloroglucinol derivative from Rumex acetosa L. Fitoterapia 2009, 80, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Bicker, J. Isolierung und Charakterisierung von Proanthocyanidinen aus Rumex acetosa L. und Myrothamnus Flabellifolia Welw, sowie Testung auf Adhäsions-Beeinflussende Eigenschaften Gegenüber Uropathogenen E. coli. Ph.D. Thesis, University of Münster, Münster, Germany, 2008. [Google Scholar]
- Hör, M.; Heinrich, M.; Rimpler, H. Proanthocyanidin polymers with antisecretory activity and proanthocyanidin oligomers from Guazuma ulmifolia bark. Phytochemistry 1996, 42, 109–119. [Google Scholar] [CrossRef]
- Zumdick, S.; Petereit, F.; Luftmann, H.; Hensel, A. Preparative isolation of oligomeric procyanidins from hawthorn (Crataegus spp.). Pharmazie 2009, 64, 286–288. [Google Scholar] [PubMed]
- Hellenbrand, N.; Sendker, J.; Lechtenberg, M.; Petereit, F.; Hensel, A. Isolation and quantification of oligomeric and polymeric procyanidins in leaves and flowers of hawthorn (Crataegus spp.). Fitoterapia 2015, 104, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Hellenbrand, N.; Lechtenberg, M.; Petereit, F.; Sendker, J.; Hensel, A. Isolation and quantification of oligomeric and polymeric procyanidins in the aerial parts of St. John’s wort (Hypericum perforatum). Planta Med. 2015. [Google Scholar] [CrossRef]
- Sendker, J.; Petereit, F.; Lautenschlager, M.; Hellenbrand, N.; Hensel, A. Phenylpropanoid-substituted procyanidins and tentatively identified procyanidin glycosides from hawthorn (Crataegus spp.). Planta Med. 2013, 79, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Guyot, S.; Vercauteren, J.; Cheynier, V. Structural determination of colourless and yellow dimers resulting from (+)-catechin coupling catalysed by grape polyphenoloxidase. Phytochemistry 1996, 42, 1279–1288. [Google Scholar] [CrossRef]
- Rohr, G.E. Analytical Investigation on and Isolation of Procyanidins from Crataegus Leaves and Flowers. Ph.D. Thesis, ETH Zürich, Zürich, Switzerland, 1999. [Google Scholar]
- Weinges, K.; Ebert, W.; Huthwelk, D.; Mattauch, H.; Perner, J. Oxidative coupling of phenols, II Structure and mechanism of formation of dehydrodicatechin A. Liebigs Ann. Chem. 1969, 726, 114–124. [Google Scholar] [CrossRef]
- Weinges, K.; Huthwelk, D. Oxydative coupling of phenols, III Isolation and constitution of a dehydro-dicatechin (b4) linked in 8,6′-position. Liebigs Ann. Chem. 1970, 731, 161–170. [Google Scholar] [CrossRef]
- Young, D.A.; Young, E.; Roux, D.G.; Brandt, E.V.; Ferreira, D. Synthesis of condensed tannins. Part 19. Phenol oxidative coupling of (+)-catechin and (+)-mesquitol. Conformation of bis-(+)-catechins. J. Chem. Soc. Perkin Trans. 1 1987, 2345–2351. [Google Scholar] [CrossRef]
- He, F.; Pan, Q.H.; Shi, Y.; Zhang, X.T.; Duan, C.Q. Identification of autoxidation oligomers of flavan-3-ols in model solutions by HPLC-MS/MS. J. Mass Spectrom. 2009, 44, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Foo, L.Y.; Lu, Y.R.; Wong, H. Biphenyl-linked biflavanoids from grape pomace. Phytochemistry 1998, 47, 1137–1140. [Google Scholar] [CrossRef]
- Ahn, B.Z.; Gstirner, F. Über Catechin-Dimere der Eichenrinde. Arch. Pharmaz. 1971, 304, 666–673. [Google Scholar] [CrossRef]
- Busia, K. Ghana Herbal Pharmacopoeia, 1st ed.; The Advent Press: Accra, Ghana, 1992. [Google Scholar]
- Rogers, C.B. Acidic dammarane arabinofuranosides from Combretum rotundifolium. Phytochemistry 1995, 40, 833–836. [Google Scholar] [CrossRef]
- Jossang, A.; Seuleiman, M.; Maidou, E.; Bodo, B. Pentacyclic triterpenes from Combretum nigricans. Phytochemistry 1996, 41, 591–594. [Google Scholar] [CrossRef]
- Pegel, K.H.; Rogers, C.B. Mollic acid 3-beta-d-glucoside, a novel 1-alpha-hydroxycycloartane saponin from Combretum molle (Combretaceae). Tetrahedron Lett. 1976, 17, 4299–4302. [Google Scholar] [CrossRef]
- Burglin, T.R.; Lobos, E.; Blaxter, M.L. Caenorhabditis elegans as a model for parasitic nematodes. Int. J. Parasitol. 1998, 28, 395–411. [Google Scholar] [CrossRef]
- Katiki, L.M.; Ferreira, J.F.; Zajac, A.M.; Masler, C.; Lindsay, D.S.; Chagas, A.C.; Amarante, A.F. Caenorhabditis elegans as a model to screen plant extracts and compounds as natural anthelmintics for veterinary use. Vet. Parasitol. 2011, 182, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ellis, B.L.; Yiu, Y.Y.; Miller, M.M.; Urban, J.F.; Shi, L.Z.; Aroian, R.V. An extensive comparison of the effect of anthelmintic classes on diverse nematodes. PLoS ONE 2013, 8, e70702. [Google Scholar] [CrossRef] [PubMed]
- Barrau, E.; Fabre, N.; Fouraste, I.; Hoste, H. Effect of bioactive compounds from sainfoin (Onobrychis viciifolia scop.) on the in vitro larval migration of Haemonchus contortus: Role of tannins and flavonol glycosides. Parasitology 2005, 131, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E. Chemistry of Tannin-Protein Complexation. In Chemistry and Significance of Condensed Tannins; Springer US: New York, NY, USA, 1989; pp. 323–333. [Google Scholar]
- Mohamed, A.S.A.; Mori, T.; Islam, S.Q.; Sato, M.; Yamasaki, T. Lethal activity of gallo- and condensed tannins against the free-living soil-inhabiting nematode, Caenorhabditis elegans. J. Pestic. Sci. 2000, 25, 410–415. [Google Scholar] [CrossRef]
- Williams, A.R.; Fryganas, C.; Ramsay, A.; Mueller-Harvey, I.; Thamsborg, S.M. Direct anthelmintic effects of condensed tannins from diverse plant sources against Ascaris suum. PLoS ONE 2014, 9, e97053. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, F.; Tsuda, Y.; Kondo, K.; Yoshimura, H.; Nishioka, I.; Nonaka, G. Studies on crude drugs effective on visceral Larva migrans. 3. The bursting activity of tannins on dog roundworm larva. Chem. Pharm. Bull. 1988, 36, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Quijada, J.; Fryganas, C.; Ropiak, H.M.; Ramsay, A.; Mueller-Harvey, I.; Hoste, H. Anthelmintic activities against Haemonchus contortus or Trichostrongylus colubriformis from small ruminants are influenced by structural features of condensed tannins. J. Agric. Food Chem. 2015, 63, 6346–6354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.R.; Ropiak, H.M.; Fryganas, C.; Desrues, O.; Mueller-Harvey, I.; Thamsborg, S.M. Assessment of the anthelmintic activity of medicinal plant extracts and purified condensed tannins against free-living and parasitic stages of Oesophagostomum dentatum. Parasites Vectors 2014, 7, 518. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.M.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Magnolato, D.; Lilley, T.H.; Haslam, E. Polyphenol complexation—Some thoughts and observations. Phytochemistry 1988, 27, 2397–2409. [Google Scholar] [CrossRef]
- Porter, L.J.; Woodruffe, J. Hemanalysis: The relative astringency of proanthocyanidin polymers. Phytochemistry 1984, 23, 1255–1256. [Google Scholar] [CrossRef]
- Katiki, L.M.; Ferreira, J.F.; Gonzalez, J.M.; Zajac, A.M.; Lindsay, D.S.; Chagas, A.C.; Amarante, A.F. Anthelmintic effect of plant extracts containing condensed and hydrolyzable tannins on Caenorhabditis elegans, and their antioxidant capacity. Vet. Parasitol. 2013, 192, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Mohamed, A.S.A.; Sato, M.; Yamasaki, T. Ellagitannin toxicity in the free-living soil-inhabiting nematode, Caenorhabditis elegans. J. Pestic. Sci. 2000, 25, 405–409. [Google Scholar] [CrossRef]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 2006, 22, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lancheros, E.; Viau, C.; Walter, T.N.; Francis, A.; Geary, T.G. Activity of novel nicotinic anthelmintics in cut preparations of Caenorhabditis elegans. Int. J. Parasitol. 2011, 41, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ohshita, Y.; Tsushida, T. Antioxidant compounds from buckwheat (Fagopyrum esculentum Moench) hulls. J. Agric. Food Chem. 1997, 45, 1039–1044. [Google Scholar] [CrossRef]
- Rayyan, S.; Fossen, T.; Nateland, H.S.; Andersen, O.M. Isolation and identification of flavonoids, including flavone rotamers, from the herbal drug “crataegi folium cum flore” (hawthorn). Phytochem. Anal. 2005, 16, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Yang, C.R. Chemical studies on Gentianella azurea, a tibetan medicinal plant. Acta Bot. Yunnan 1993, 16, 401–406. [Google Scholar]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ndjonka, D.; Agyare, C.; Luersen, K.; Djafsia, B.; Achukwi, D.; Nukenine, E.N.; Hensel, A.; Liebau, E. In vitro activity of Cameroonian and Ghanaian medicinal plants on parasitic (Onchocerca ochengi) and free-living (Caenorhabditis elegans) nematodes. J. Helminthol. 2011, 85, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Lenaerts, I.; Walker, G.A.; Van Hoorebeke, L.; Gems, D.; Vanfleteren, J.R. Dietary restriction of Caenorhabditis elegans by axenic culture reflects nutritional requirement for constituents provided by metabolically active microbes. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of all compounds, except OPC decamer are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spiegler, V.; Sendker, J.; Petereit, F.; Liebau, E.; Hensel, A. Bioassay-Guided Fractionation of a Leaf Extract from Combretum mucronatum with Anthelmintic Activity: Oligomeric Procyanidins as the Active Principle. Molecules 2015, 20, 14810-14832. https://doi.org/10.3390/molecules200814810
Spiegler V, Sendker J, Petereit F, Liebau E, Hensel A. Bioassay-Guided Fractionation of a Leaf Extract from Combretum mucronatum with Anthelmintic Activity: Oligomeric Procyanidins as the Active Principle. Molecules. 2015; 20(8):14810-14832. https://doi.org/10.3390/molecules200814810
Chicago/Turabian StyleSpiegler, Verena, Jandirk Sendker, Frank Petereit, Eva Liebau, and Andreas Hensel. 2015. "Bioassay-Guided Fractionation of a Leaf Extract from Combretum mucronatum with Anthelmintic Activity: Oligomeric Procyanidins as the Active Principle" Molecules 20, no. 8: 14810-14832. https://doi.org/10.3390/molecules200814810