
Escherichia coli ASKA Clone Library Harboring tRNA-Specific Adenosine Deaminase (tadA) Reveals Resistance towards Xanthorrhizol

Abstract
:1. Introduction
2. Results and Discussion
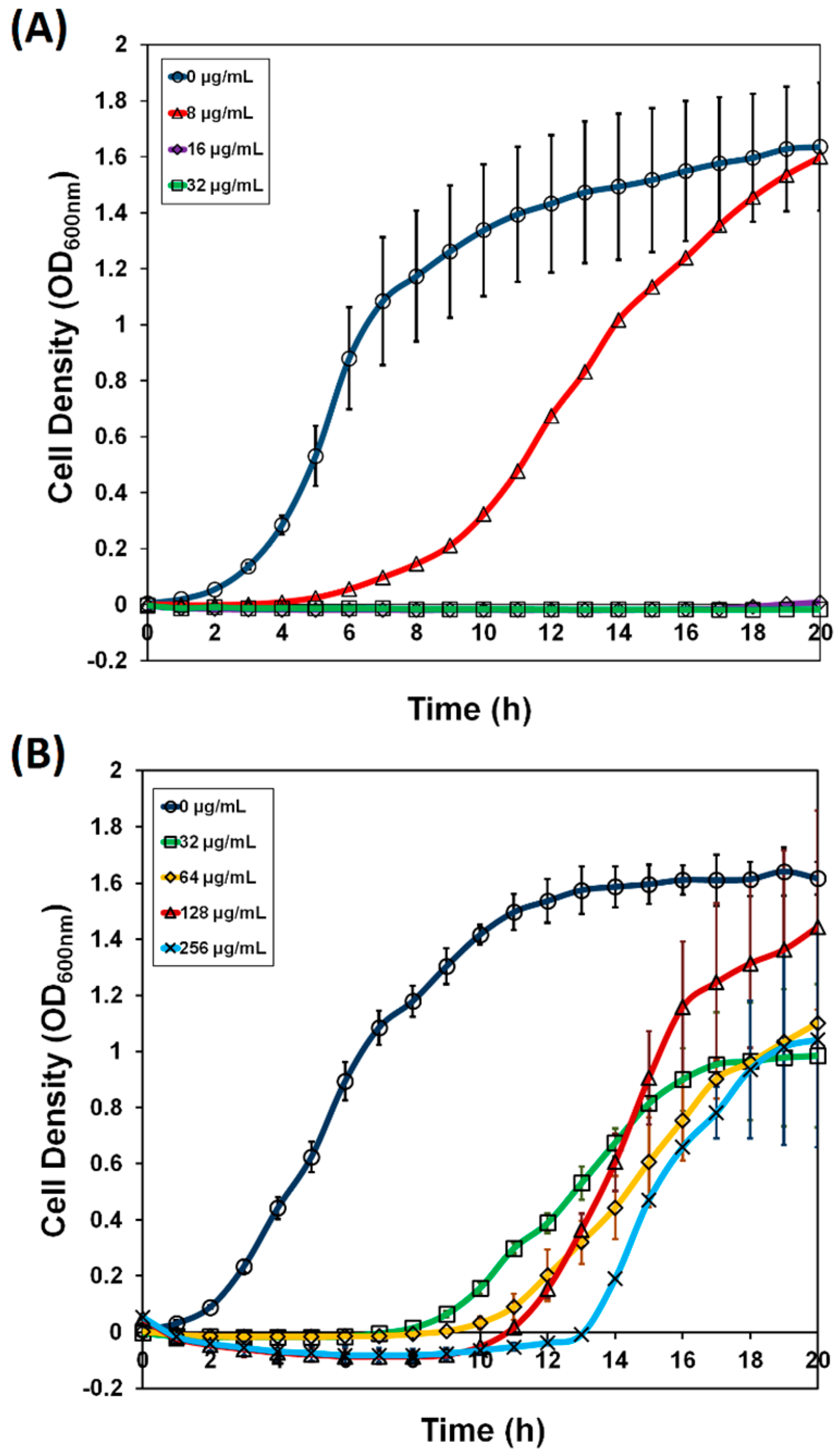
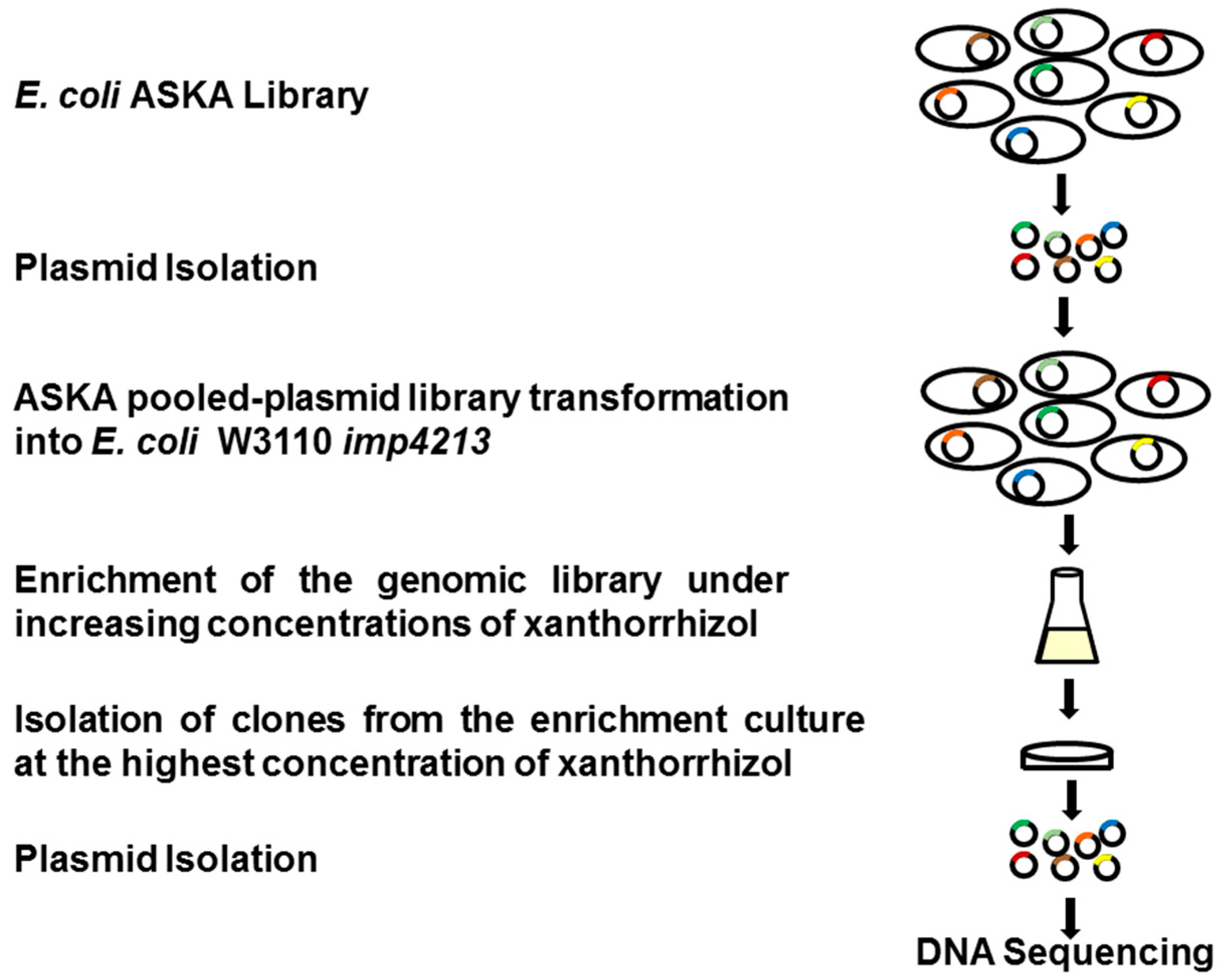
2.1. ASKA Library Enrichment and Screening for Xanthorrhizol Resistance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Function | COG Classification a | Essentiality b |
|---|---|---|---|
| tadA | tRNA-Specific Adenine Deaminase | Translation | Essential |
| galU | Glucose-1-Phosphate Uridylyltransferase | Cell Envelope Biogenesis, Outer Membrane | Non-essential |
| ydaC | Protein Involved in Chromosome Maintenance/Rac Prophage Gene | DNA Replication, Recombination, and Repair | Non-essential |
| fucU | l-Fucose Mutarotase | Carbohydrate Transport and Metabolism | Non-essential |
| mltF (yfhD) | Membrane-Bound Lytic Murein Transglycosylase F | Cell Envelope Biogenesis, Outer Membrane | Non-essential |
| ydeA | Sugar Efflux Transporter | Carbohydrate Transport and Metabolism | Non-essential |
| soxS | DNA-Binding Transcriptional Regulator | Transcription | Non-essential |
| yiiD | Predicted Acetyltransferase | General Function Prediction Only | Non-essential |
| nrdH (ygaN) | Glutaredoxin-Like Protein | Post-Translational Modification, Protein Turnover, Chaperones | Non-essential c |
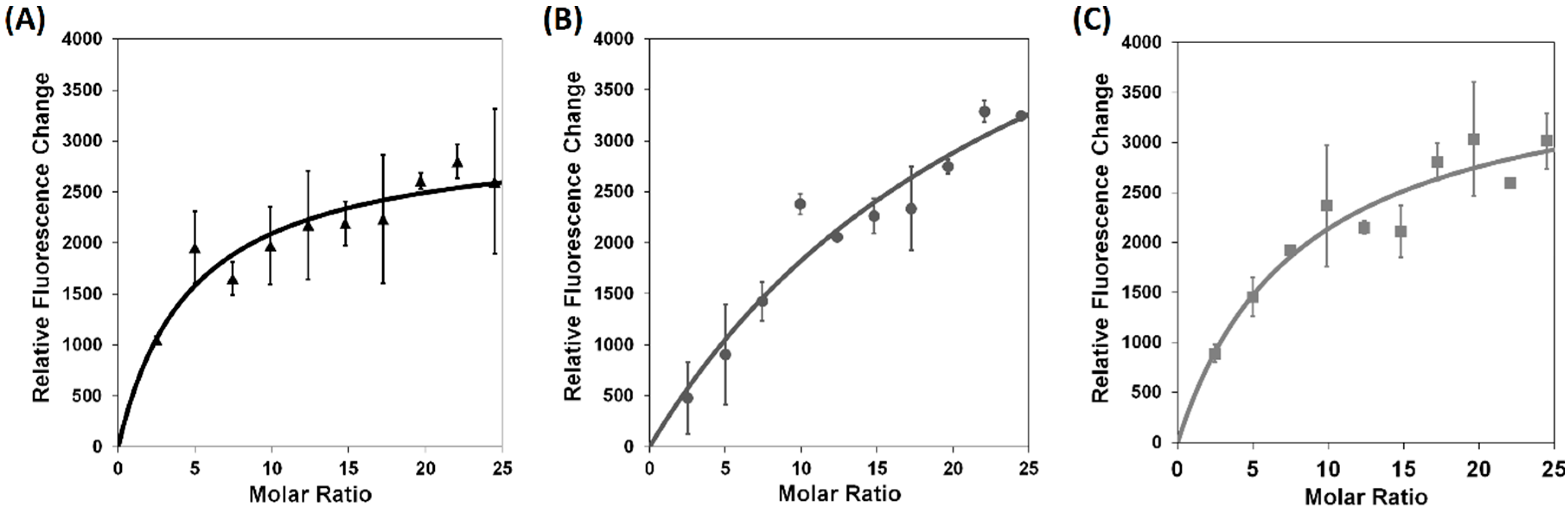
2.2. Verification of tadA Overexpression-Induced Resistance to Xanthorrhizol in E. coli W3110 imp4213 and Bacillus subtilis 168, Site-Directed Mutagenesis, and Binding Study
| Bacterial Strain | MIC (μg/mL) |
|---|---|
| E. coli W3110 imp4213 | 16 |
| E. coli W3110 imp4213 (pCA24N) | 16 |
| E. coli W3110 imp4213 (pCA24N-tadA) | 128 |
| E. coli W3110 imp4213 (pCA24N-tadA (A143V)) | 256 |
| E. coli W3110 imp4213 (pCA24N-tadA (F149G)) | 256 |
| Bacillus subtilis 168 | 8 |
| B. subtilis 168 (pHT255) | 8 |
| B. subtilis 168 (pHT255-tadA) | 32 |

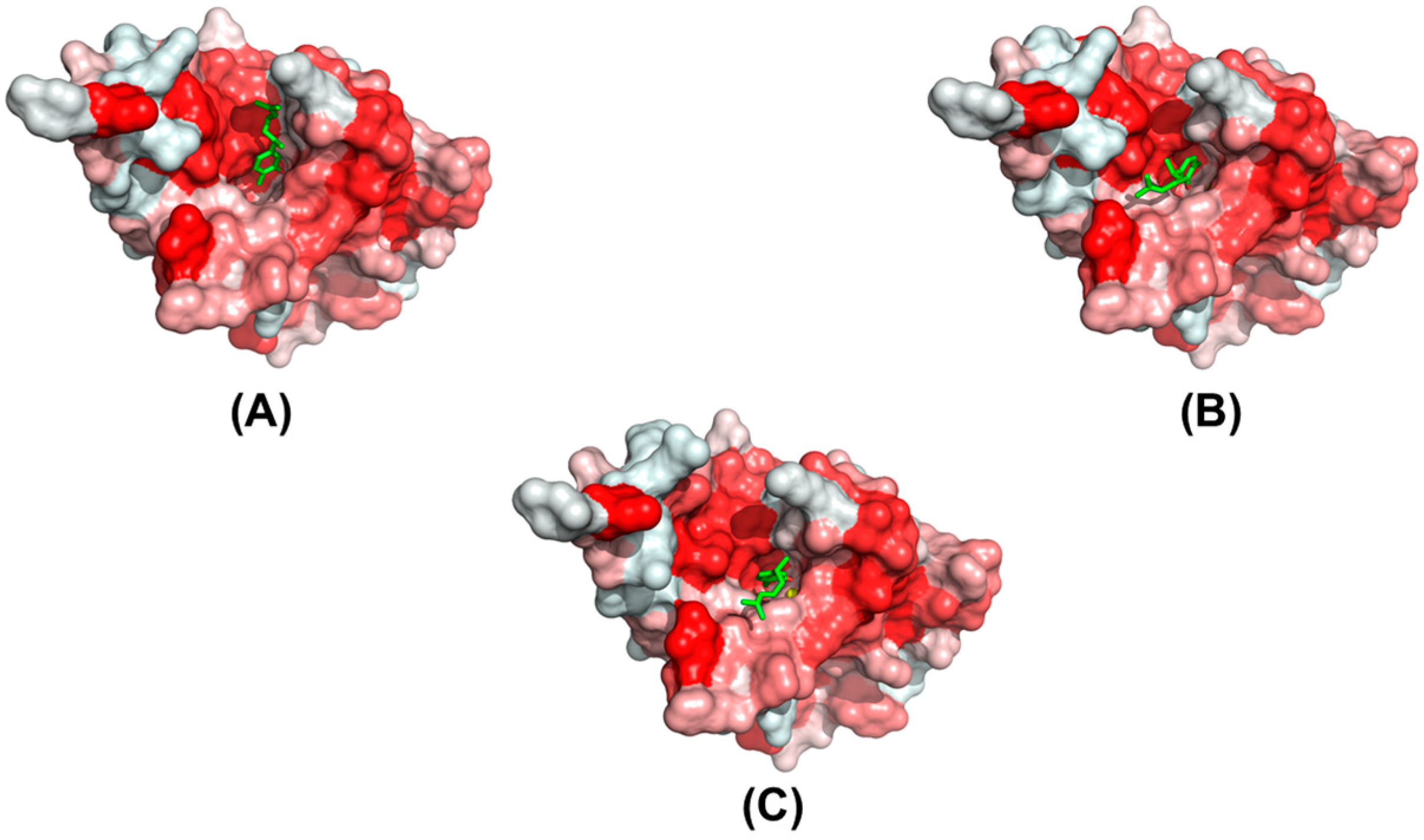
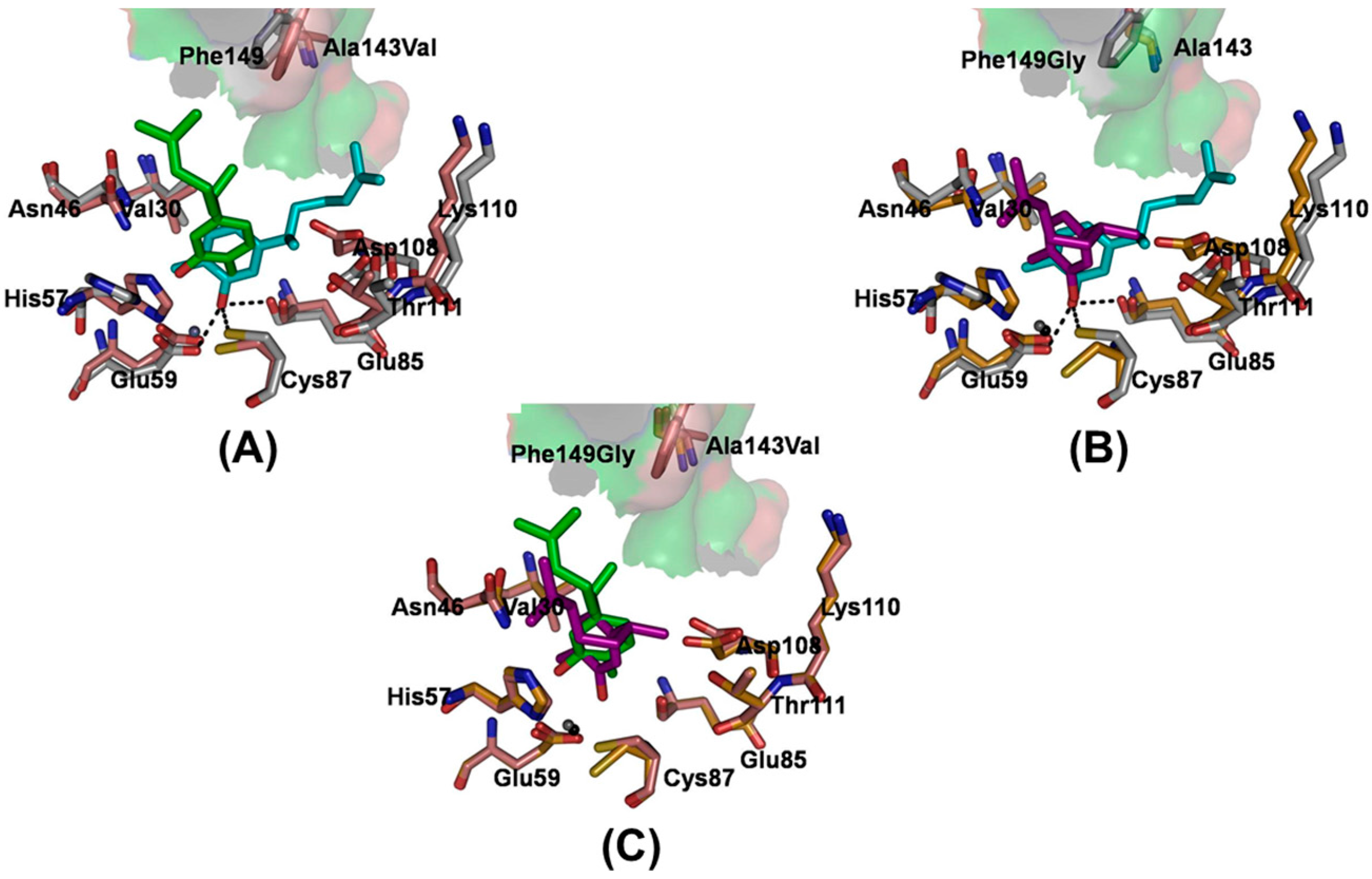
2.3. Modeling of the Binding Mechanism of Xanthorrhizol to Wild-Type and Mutant EcTadA


3. Experimental Section
3.1. Chemicals, Bacterial Strains, and Plasmids
| Name | Description | Source |
|---|---|---|
| E. coli Strain | ||
| DH5α | F− endA1 glnV44 thi-1 recA1 relA1 gyrA96 deoR nupG Φ80dlacZΔM15 Δ(lacZYA-argF)U169, hsdR17(rK− mK+), λ– | Laboratory Stock |
| W3110 imp4213 | F− λ− rph-1 INV(rrnD, rrnE) Δimp::TcR | Laboratory Stock |
| W3110 imp4213 (pCA24N) | W3110 Δimp + pCA24N | Laboratory Stock |
| W3110 imp4213 + pCA24N-tadA | tadA overexpression | This Study |
| W3110 imp4213 + pCA24N-tadA (A143V) | tadA (A143V) overexpression | This Study |
| W3110 imp4213 + pCA24N-tadA (F149G) | tadA (F149G) overexpression | This Study |
| Bacillus strain | ||
| B. subtilis 168 | trpC2 (Trp−) | Laboratory Stock |
| B. subtilis 168 (pHT255) | B. subtilis 168 + pHT255 | This Study |
| B. subtilis 168 (pHT255-tadA) | tadA overexpression | This Study |
| Primer | ||
| A143V-F | GTGCGTGGCGTTGCTCAGTGAC | This Study |
| A143-R | TCATCCGCCAGTATTCCTTCC | This Study |
| F149G-F | AGTGACGGCTTTCGCATGCGCCGCCAG | This Study |
| F149-R | GAGCAACGCCGCGCACTCATCCGC | This Study |
| bstadA-F | CCGGCTCTAGAATGACACAAGATG AACTTTATATGAAAGAAGC | This Study |
| bstadA-R | CCGGCGACGTCCTATTCAGACAAG TTTTTCCTGGC | This Study |
3.2. ASKA Pooled-Plasmid Library Construction
3.3. Xanthorrhizol-Resistance Library Enrichment and Screening

3.4. Verification of tadA Overexpression-Induced Resistance to Xanthorrhizol and Site-Directed Mutagenesis
3.5. tadA Overexpression in B. subtilis 168
3.6. Xanthorrhizol-tadA Binding Study
3.7. Computational Methods
3.7.1. Molecular Docking
3.7.2. Molecular Dynamics Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Hwang, J.K.; Shim, J.S.; Baek, N.I.; Pyun, Y.R. Xanthorrhizol: A potential antibacterial agent from Curcuma xanthorrhiza against Streptococcus mutans. Planta Med. 2000, 66, 196–197. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.K.; Shim, J.S.; Pyun, Y.R. Antibacterial activity of xanthorrhizol from Curcuma xanthorrhiza against oral pathogens. Fitoterapia 2000, 71, 321–323. [Google Scholar] [CrossRef]
- Lee, Y.L.; Shim, J.S.; Rukayadi, Y.; Hwang, J.K. Antibacterial activity of xanthorrhizol isolated from Curcuma xanthorrhiza Roxb. against foodborne pathogens. J. Food Prot. 2008, 71, 1926–1930. [Google Scholar] [PubMed]
- Yogiara; Kim, D.; Hwang, J.K.; Pan, J.G. The food-grade antimicrobial xanthorrhizol targets the enoyl-ACP reductase or FabI in bacteria. Bioorg. Med. Chem. Lett. 2015. under review. [Google Scholar]
- Park, H.S.; Yoon, Y.M.; Jung, S.J.; Kim, C.M.; Kim, J.M.; Kwak, J.-H. Antistaphylococcal activities of CG400549, a new bacterial enoyl-acyl carrier protein reductase (FabI) inhibitor. J. Antimicrob. Chemother. 2007, 60, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Yoon, Y.M.; Jung, S.J.; Yun, I.N.R.; Kim, C.M.; Kim, J.M.; Kwak, J.-H. CG400462, a new bacterial enoyl-acyl carrier protein reductase (FabI) inhibitor. Int. J. Antimicrob. Agents 2007, 30, 446–451. [Google Scholar] [CrossRef] [PubMed]
- NBRP E.coli Strain. Available online: http://www.shigen.nig.ac.jp/ecoli/strain (accessed on 2 October 2014).
- Kitagawa, M.; Ara, T.; Arifuzzaman, M.; Ioka-Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Mori, H. Complete set of ORF clones of Escherichia coli ASKA library (a complete set of E. coli K-12 ORF archive): Unique resources for biological research. DNA Res. 2006, 12, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Couce, A.; Briales, A.; Rodríguez-Rojas, A.; Costas, C.; Pascual, Á.; Blázquez, J. Genomewide overexpression screen for fosfomycin resistance in Escherichia coli: murA confers clinical resistance at low fitness cost. Antimicrob. Agents Chemother. 2012, 56, 2767–2769. [Google Scholar] [CrossRef] [PubMed]
- Soo, V.W.C.; Hanson-Manful, P.; Patrick, W.M. Artificial gene amplification reveals an abundance of promiscuous resistance determinants in Escherichia coli. Proc. Natl. Acad. Sci. USA 2011, 108, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Kim, J.; Ju, H.; Choi, S.-K.; Hwang, S.; Park, S.; Kim, E.; Pan, J.-G. Genome-wide enrichment screening reveals multiple targets and resistance genes for triclosan in Escherichia coli. J. Microbiol. 2012, 50, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Silhavy, T.J. Imp/OstA is required for cell envelope biogenesis in Escherichia coli. Mol. Microbiol. 2002, 45, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, N.; Falcone, B.; Kahne, D.; Silhavy, T.J. Chemical conditionality: A genetic strategy to probe organelle assembly. Cell 2005, 121, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Sampson, B.A.; Misra, R.; Benson, S.A. Identification and characterization of a new gene of Escherichia coli K-12 involved in outer membrane permeability. Genetics 1989, 122, 491–501. [Google Scholar] [PubMed]
- Database of Essential Gene. Available online: http://www.essentialgene.org (accessed on 25 September 2013).
- Wolf, J.; Gerber, A.P.; Keller, W. tadA, an essential tRNA-specific adenosine deaminase from Escherichia coli. EMBO J. 2002, 21, 3841–3851. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, S.Y.; Scholle, M.D.; Campbell, J.W.; Balázsi, G.; Ravasz, E.; Daugherty, M.D.; Somera, A.L.; Kyrpides, N.C.; Anderson, I.; Gelfand, M.S.; et al. Experimental determination and system level analysis of essential genes in Escherichia coli MG1655. J. Bacteriol. 2003, 185, 5673–5684. [Google Scholar] [CrossRef] [PubMed]
- Deutschbauer, A.; Price, M.N.; Wetmore, K.M.; Shao, W.; Baumohl, J.K.; Xu, Z.; Nguyen, M.; Tamse, R.; Davis, R.W.; Arkin, A.P. Evidence-based annotation of gene function in Shewanella oneidensis MR-1 using genome-wide fitness profiling across 121 conditions. PLoS Genet. 2011, 7, e1002385. [Google Scholar] [CrossRef] [PubMed]
- OGEEdb. Available online: http://ogeedb.embl.de (accessed on 25 September 2013).
- Schaub, M.; Keller, W. RNA editing by adenosine deaminases generates RNA and protein diversity. Biochimie 2002, 84, 791–803. [Google Scholar] [CrossRef]
- Gerber, A.P.; Keller, W. RNA editing by base deamination: More enzymes, more targets, new mysteries. Trends Biochem. Sci. 2001, 26, 376–384. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.I. A genomic perspective on protein families. Science 1997, 278, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Lin, Y.; Gao, F.; Zhang, C.-T.; Zhang, R. DEG 10, an update of the database of essential genes that includes both protein-coding genes and noncoding genomic elements. Nucleic Acids Res. 2014, 42, D574–D580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lin, Y. DEG 5.0, a database of essential genes in both prokaryotes and eukaryotes. Nucleic Acids Res. 2009, 37 (Suppl. 1), D455–D458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ou, H.Y.; Zhang, C.T. DEG: A database of essential genes. Nucleic Acids Res. 2004, 32 (Suppl. 1), D271–D272. [Google Scholar] [CrossRef] [PubMed]
- Jayakar, H.; Parvathareddy, J.; Fitzpatrick, E.; Bina, X.; Bina, J.; Re, F.; Emery, F.; Miller, M. A galU mutant of Francisella tularensis is attenuated for virulence in a murine pulmonary model of tularemia. BMC Microbiol. 2011, 11, 179. [Google Scholar] [CrossRef] [PubMed]
- Felczak, M.M.; Kaguni, J.M. The rcbA gene product reduces spontaneous and induced chromosome breaks in Escherichia coli. J. Bacteriol. 2012, 194, 2152–2164. [Google Scholar] [CrossRef] [PubMed]
- Scheurwater, E.M.; Clarke, A.J. The C-terminal domain of Escherichia coli YfhD functions as a lytic transglycosylase. J. Biol. Chem. 2008, 283, 8363–8373. [Google Scholar] [CrossRef] [PubMed]
- Bennik, M.H.J.; Pomposiello, P.J.; Thorne, D.F.; Demple, B. Defining a rob regulon in Escherichia coli by using transposon mutagenesis. J. Bacteriol. 2000, 182, 3794–3801. [Google Scholar] [CrossRef] [PubMed]
- Zafar, M.A.; Sanchez-Alberola, N.; Wolf, R.E., Jr. Genetic evidence for a novel interaction between transcriptional activator SoxS and region 4 of the σ70 subunit of RNA polymerase at class II SoxS-dependent promoters in Escherichia coli. J. Mol. Biol. 2011, 407, 333–353. [Google Scholar] [CrossRef] [PubMed]
- McMurry, L.M.; Oethinger, M.; Levy, S.B. Overexpression of marA, soxS, or acrAB produces resistance to triclosan in laboratory and clinical strains of Escherichia coli. FEMS Microbiol. Lett. 1998, 166, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.N.; Levy, S.B. Regulation of chromosomally mediated multiple antibiotic resistance: The mar regulon. Antimicrob. Agents Chemother. 1997, 41, 2067–2075. [Google Scholar] [PubMed]
- Lee, J.-H.; Lee, K.-L.; Yeo, W.-S.; Park, S.-J.; Roe, J.-H. SoxRS-Mediated lipopolysaccharide modification enhances resistance against multiple drugs in Escherichia coli. J. Bacteriol. 2009, 191, 4441–4450. [Google Scholar] [CrossRef] [PubMed]
- Butland, G.; Peregrin-Alvarez, J.M.; Li, J.; Yang, W.; Yang, X.; Canadien, V.; Starostine, A.; Richards, D.; Beattie, B.; Krogan, N.; et al. Interaction network containing conserved and essential protein complexes in Escherichia coli. Nature 2005, 433, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Reddy, C.C.; Krishnasastry, M.V.; Surolia, N.; Surolia, A. Slow-tight binding inhibition of enoyl acyl carrier protein reductase from Plasmodium falciparum by triclosan. Biochem. J. 2004, 381, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Cummings, J.E.; England, K.; Slayden, R.A.; Tonge, P.J. Mechanism and inhibition of the FabI enoyl-ACP reductase from Burkholderia pseudomallei. J. Antimrob. Chemother. 2011, 66, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Sun, X.; Jarvis, T.; Critchley, I.; Janjic, N. Aminoacyl-tRNA synthetases: Essential and still promising targets for new anti-infective agents. Expert Opin. Investig. Drugs 2007, 16, 573–593. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Meng, Q.; Bai, L.; Zhou, H. In silico discovery of aminoacyl-tRNA synthetase inhibitors. Int. J. Mol. Sci. 2014, 15, 1358–1373. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-Q.; Flavin, M.T.; Flavin, J. Combating multidrug-resistant Gram-negative bacteria infections. Expert Opin. Investig. Drugs 2014, 23, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.M. MRSA patients: Proven methods to treat colonization and infection. J. Hosp. Infect. 2001, 48 (Suppl. A), S9–S14. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Bron, S. Plasmids. In Molecular Biological Methods for Bacillus; Harwood, C.R., Cutting, S.M., Eds.; John Wiley & Sons: Chichester, UK, 1990; pp. 75–174. [Google Scholar]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, M.L.; Cole, J.C.; Hartshorn, M.J.; Murray, C.W.; Taylor, R.D. Improved protein-ligand docking using GOLD. Proteins 2003, 52, 609–623. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System, version 1.5.0.4; Schrödinger, LLC: New York, NY, USA, 2000.
- Maestro, version 9.2; Schrödinger, LLC: New York, NY, USA, 2011.
- MacroModel, version 9.9; Schrödinger, LLC: New York, NY, USA, 2011.
- Marelius, J.; Kolmodin, K.; Feierberg, I.; Åqvist, J. Q: A molecular dynamics program for free energy calculations and empirical valence bond simulations in biomolecular systems. J. Mol. Graph. Model. 1998, 16, 213–225. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- King, G.; Warshel, A. A surface constrained all-atom solvent model for effective simulations of polar solutions. J. Chem. Phys. 1989, 91, 3647–3661. [Google Scholar] [CrossRef]
- Lee, F.S.; Warshel, A. A local reaction field method for fast evaluation of long-range electrostatic interactions in molecular simulations. J. Chem. Phys. 1992, 97, 3100–3107. [Google Scholar] [CrossRef]
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yogiara; Kim, D.; Hwang, J.-K.; Pan, J.-G. Escherichia coli ASKA Clone Library Harboring tRNA-Specific Adenosine Deaminase (tadA) Reveals Resistance towards Xanthorrhizol. Molecules 2015, 20, 16290-16305. https://doi.org/10.3390/molecules200916290
Yogiara, Kim D, Hwang J-K, Pan J-G. Escherichia coli ASKA Clone Library Harboring tRNA-Specific Adenosine Deaminase (tadA) Reveals Resistance towards Xanthorrhizol. Molecules. 2015; 20(9):16290-16305. https://doi.org/10.3390/molecules200916290
Chicago/Turabian StyleYogiara, Dooil Kim, Jae-Kwan Hwang, and Jae-Gu Pan. 2015. "Escherichia coli ASKA Clone Library Harboring tRNA-Specific Adenosine Deaminase (tadA) Reveals Resistance towards Xanthorrhizol" Molecules 20, no. 9: 16290-16305. https://doi.org/10.3390/molecules200916290