
Functional Analysis of the Fusarielin Biosynthetic Gene Cluster

 ,
,
Abstract
:1. Introduction
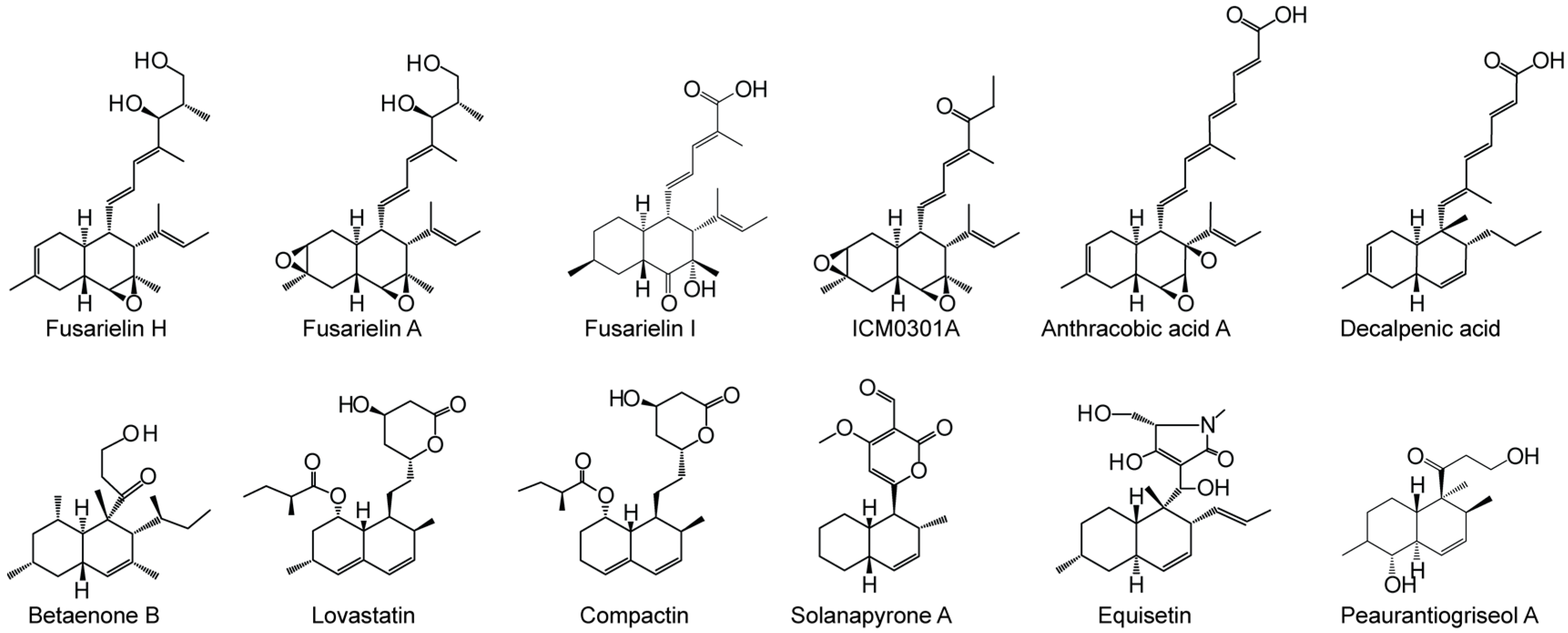
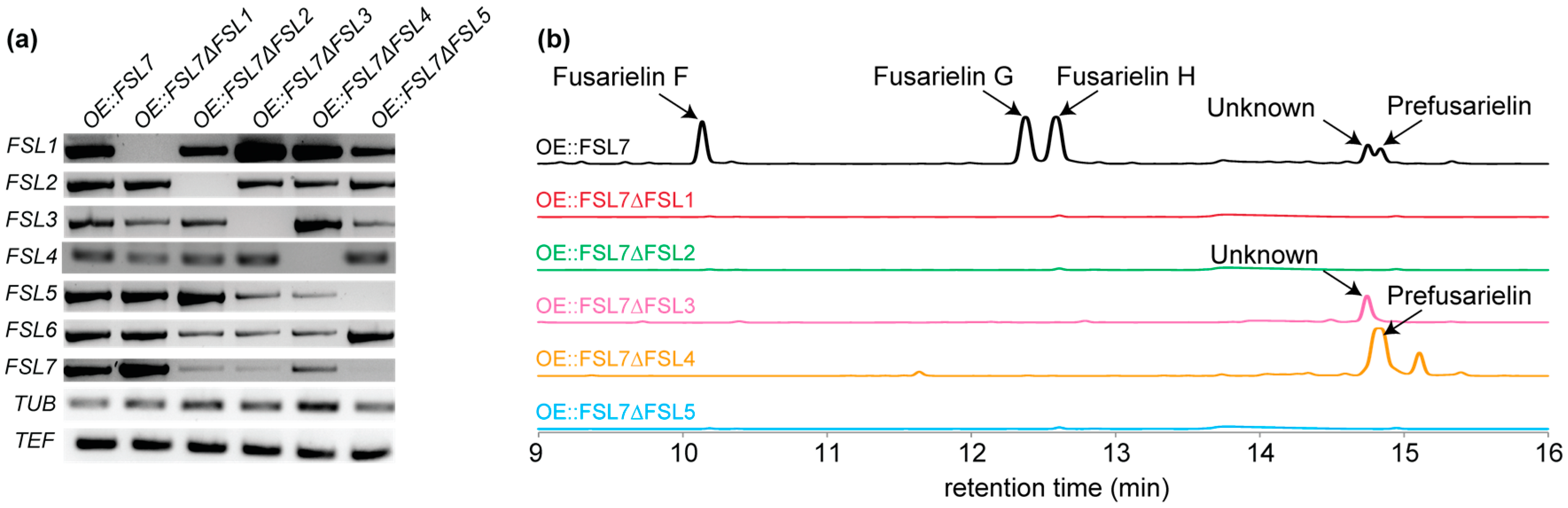
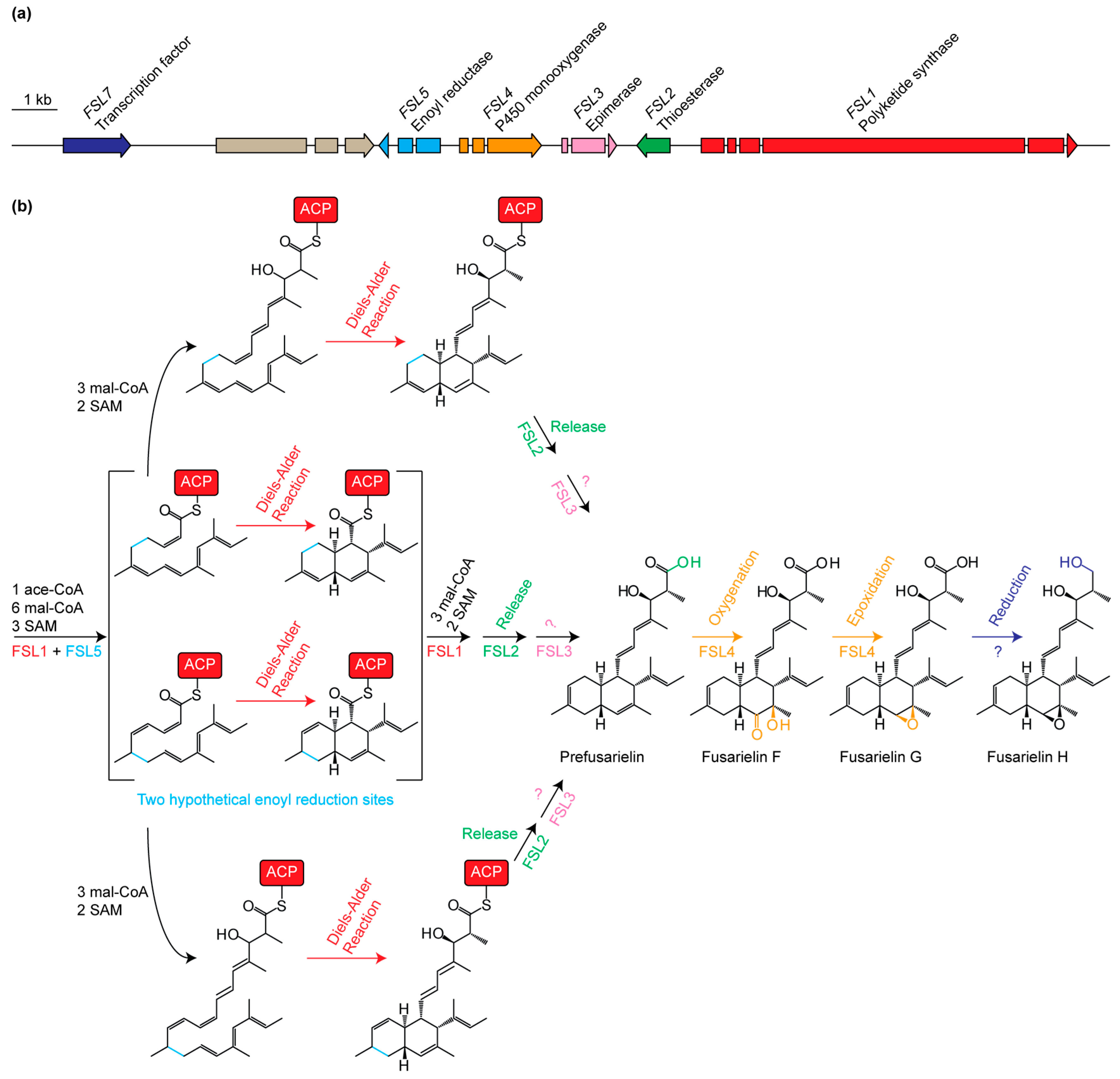
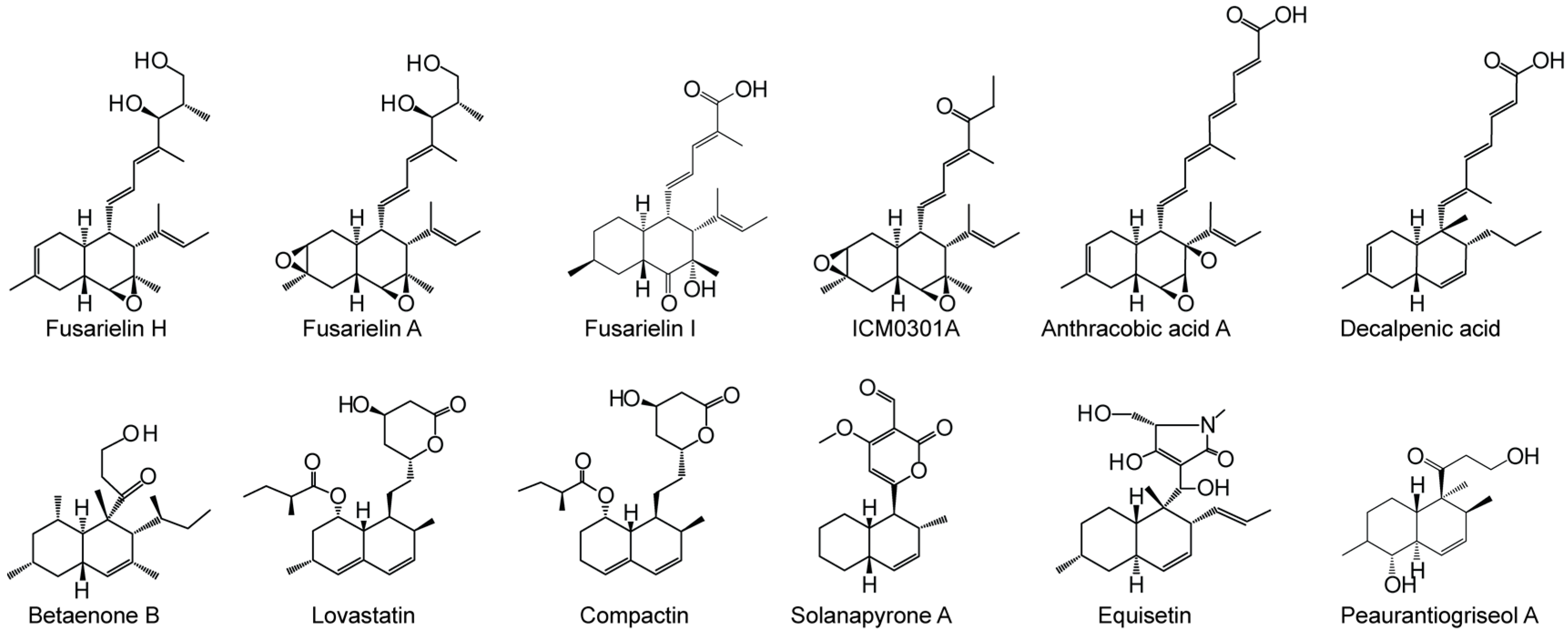
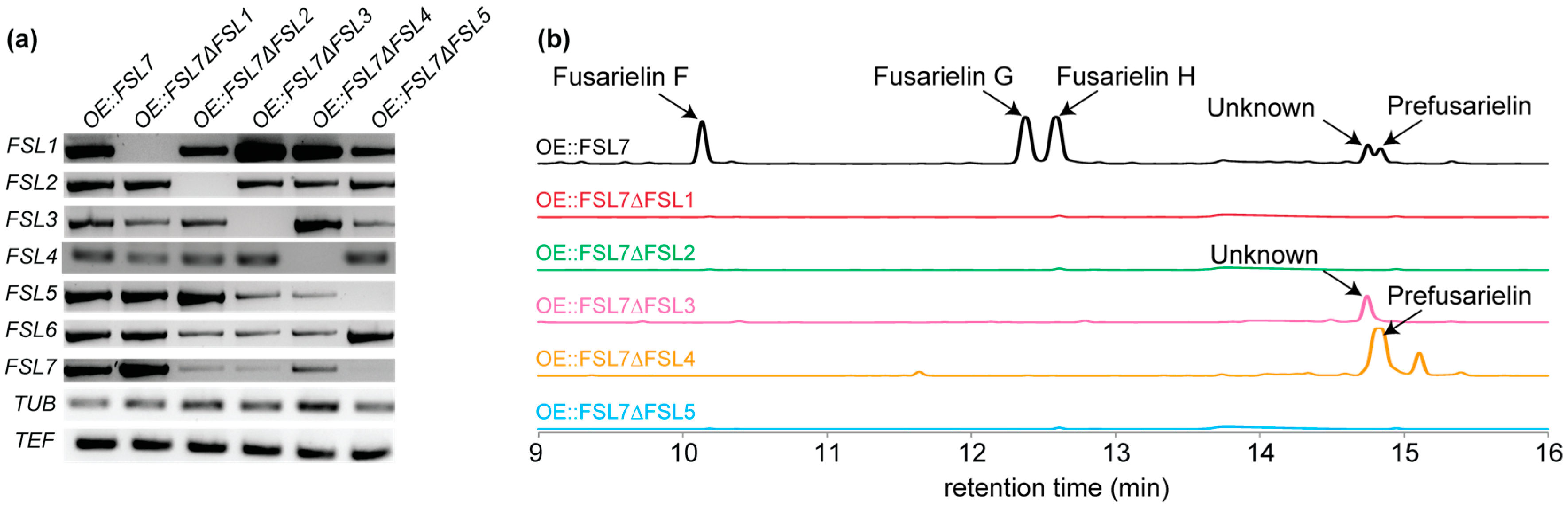
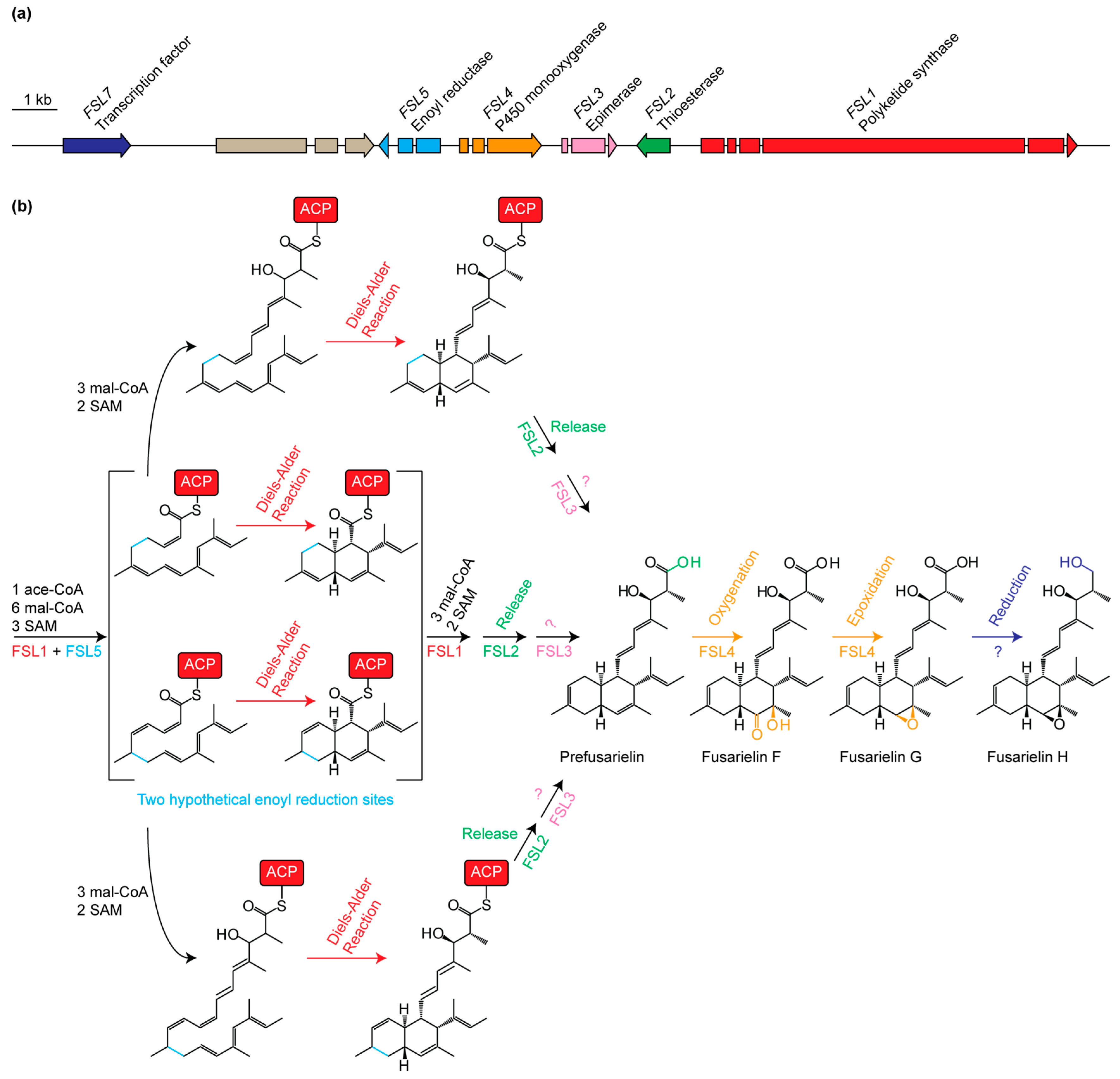
2. Results and Discussion
3. Materials and Methods
3.1. Generation of FSL1-5 Knock Out Mutants
3.2. Transcription Analyses
3.3. Metabolite Profiling
3.4. Isolation and Elucidation of Prefusarielin
3.5. Docking Analyses of FSL4
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, G.; Kusari, S.; Spiteller, M. Natural products containing ‘decalin’ motif in microorganisms. Nat. Prod. Rep. 2014, 31, 1175–1201. [Google Scholar] [CrossRef] [PubMed]
- Kelly, W.L. Intramolecular cyclizations of polyketide biosynthesis: Mining for a “Diels-Alderase”? Org. Biomol. Chem. 2008, 6, 4483–4493. [Google Scholar] [CrossRef] [PubMed]
- Auclair, K.; Sutherland, A.; Kennedy, J.; Witter, D.J.; Van den Heever, J.P.; Hutchinson, C.R.; Vederas, J.C. Lovastatin nonaketide synthase catalyzes an intramolecular Diels-Alder reaction of a substrate analogue. J. Am. Chem. Soc. 2000, 122, 11519–11520. [Google Scholar] [CrossRef]
- Kasahara, K.; Miyamoto, T.; Fujimoto, T.; Oguri, H.; Tokiwano, T.; Oikawa, H.; Ebizuka, Y.; Fujii, I. Solanapyrone synthase, a possible Diels-Alderase and iterative type I polyketide synthase encoded in a biosynthetic gene cluster from Alternaria solani. ChemBioChem 2010, 11, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Nogawa, T.; Hirota, H.; Jang, J.-H.; Takahashi, S.; Ahn, J.S.; Osada, H. A new enzyme involved in the control of the stereochemistry in the decalin formation during equisetin biosynthesis. Biochem. Biophys. Res. Commun. 2015, 460, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Auclair, K.; Kendrew, S.G.; Park, C.; Vederas, J.C.; Hutchinson, C.R. Modulation of polyketide synthase activity by accessory proteins during lovastatin biosynthesis. Science 1999, 284, 1368–1372. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, J.L.; Auclair, K.; Kennedy, J.; Hutchinson, C.R.; Vederas, J.C. Transformations of cyclic nonaketides by Aspergillus terreus mutants blocked for lovastatin biosynthesis at the lovA and lovC genes. Org. Biomol. Chem. 2003, 1, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Ugai, T.; Minami, A.; Fujii, R.; Tanaka, M.; Oguri, H.; Gomi, K.; Oikawa, H. Heterologous expression of highly reducing polyketide synthase involved in betaenone biosynthesis. Chem. Commun. 2015, 51, 1878–1881. [Google Scholar] [CrossRef] [PubMed]
- Kakule, T.B.; Sardar, D.; Lin, Z.J.; Schmidt, E.W. Two related pyrrolidinedione synthetase loci in Fusarium heterosporum ATCC 74349 produce divergent metabolites. ACS Chem. Biol. 2013, 8, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Chooi, Y.H.; Choi, J.W.; Li, S.; Vederas, J.C.; Da Silva, N.A.; Tang, Y. LovG: The thioesterase required for dihydromonacolin L release and lovastatin nonaketide synthase turnover in lovastatin biosynthesis. Angew. Chem. Int. Ed. Engl. 2013, 52, 6472–6475. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Sunaga, R.; Furihata, K.; Morisaki, N.; Iwasaki, S. Isolation and structures of an antifungal antibiotic, fusarielin A, and related compounds produced by a Fusarium sp. J. Antibiot. 1995, 48, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.L.; Hansen, F.T.; Sondergaard, T.E.; Staerk, D.; Lee, T.V.; Wimmer, R.; Klitgaard, L.G.; Purup, S.; Giese, H.; Frandsen, R.J. Production of novel fusarielins by ectopic activation of the polyketide synthase 9 cluster in Fusarium graminearum. Environ. Microbiol. 2012, 14, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.P.; Zhang, D.H.; Lee, U.; Kang, J.S.; Choi, H.D.; Son, B.W. Dehydroxychlorofusarielin B, an antibacterial polyoxygenated decalin derivative from the marine-derived fungus Aspergillus sp. J. Nat. Prod. 2007, 70, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.; Walter, M.; Lee, W.; Mannhaupt, G.; Munsterkotter, M.; Mewes, H.W.; Adam, G.; Guldener, U. FGDB: Revisiting the genome annotation of the plant pathogen Fusarium graminearum. Nucleic Acids Res. 2011, 39, D637–D639. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.D.; Nguyen, C.; Bruegger, J.; Smith, P.; Xu, W.; Ma, S.; Wong, E.; Wong, S.; Xie, X.; Li, J.W.-H.; et al. Crystal structure and biochemical studies of the trans-acting polyketide enoyl reductase LovC from lovastatin biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 11144–11149. [Google Scholar] [CrossRef] [PubMed]
- Gay, D.C.; Spear, P.J.; Keatinge-Clay, A.T. A Double-hotdog with a New Trick: Structure and Mechanism of the trans-Acyltransferase Polyketide Synthase Enoyl-isomerase. ACS Chem. Biol. 2014, 9, 2374–2381. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Ward, S.; Chung, L.; Carney, J.R.; Li, Y.; Reid, R.; Katz, L. Elucidating the Mechanism of cis Double Bond Formation in Epothilone Biosynthesis. J. Am. Chem. Soc. 2004, 126, 46–47. [Google Scholar] [CrossRef] [PubMed]
- He, H.-Y.; Tang, M.-C.; Zhang, F.; Tang, G.-L. Cis-Double Bond Formation by Thioesterase and Transfer by Ketosynthase in FR901464 Biosynthesis. J. Am. Chem. Soc. 2014, 136, 4488–4491. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, N.; Alhamadsheh, M.M.; Reynolds, K.A. Cis-Δ2,3-Double Bond of Phoslactomycins Is Generated by a Post-PKS Tailoring Enzyme. J. Am. Chem. Soc. 2008, 130, 12236–12237. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Q.; Aktas, N.; Konuklugil, B.; Mandi, A.; Daletos, G.; Lin, W.H.; Dai, H.F.; Kurtan, T.; Proksch, P. A new fusarielin analogue from Penicillium sp. isolated from the Mediterranean sponge Ircinia oros. Tetrahedron Lett. 2015, 56, 5317–5320. [Google Scholar] [CrossRef]
- Josefsen, L.; Droce, A.; Sondergaard, T.E.; Sørensen, J.L.; Bormann, J.; Schaefer, W.; Giese, H.; Olsson, S. Autophagy provides nutrients for nonassimilating fungal structures and is necessary for plant colonization but not for infection in the necrotrophic plant pathogen Fusarium graminearum. Autophagy 2012, 8, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, R.J.N.; Andersson, J.A.; Kristensen, M.B.; Giese, H. Efficient four fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Mol. Biol. 2008, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, F.T.; Droce, A.; Sørensen, J.L.; Fojan, P.; Giese, H.; Sondergaard, T.E. Overexpression of NRPS4 leads to increased surface hydrophobicity in Fusarium graminearum. Fungal Biol. 2012, 116, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Yoder, W.T.; Christianson, L.M. Species-specific primers resolve members of Fusarium section Fusarium: Taxonomic status of the edible “Quorn” fungus reevaluated. Fungal Genet. Biol. 1998, 23, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS “KNAW” Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010. [Google Scholar]
- Sørensen, J.L.; Nielsen, K.F.; Sondergaard, T.E. Redirection of pigment biosynthesis to isocoumarins in Fusarium. Fungal Genet. Biol. 2012, 49, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Vriend, G. YASARA View-molecular graphics for all devices-from smartphones to workstations. Bioinformatics 2014, 30, 2981–2982. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Darden, T.; Nabuurs, S.B.; Finkelstein, A.; Vriend, G. Making optimal use of empirical energy functions: Force-field parameterization in crystal space. Proteins Struct. Funct. Bioinform. 2004, 57, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins Struct. Funct. Bioinform. 2009, 77, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Hooft, R.W.W.; Vriend, G.; Sander, C.; Abola, E.E. Errors in protein structures. Nature 1996, 381, 272. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. Software news and update Autodock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Gene Name (Accession Number) [14] | Protein Family | Proposed Function |
|---|---|---|
| FSL1 (FGSG_10464) | Polyketide synthase | Reducing PKS; condensation of ten acetate units |
| FSL2 (FGSG_10463) | Thioesterase | Release of product from FSL1 |
| FSL3 (FGSG_17368) | Aldose 1-epimerase | Epimerization of unknown entry product |
| FSL4 (FGSG_10461) | Cytochrome P450 | Oxygenation of C-15 and C-16 |
| FSL5 (FGSG_17367) | Enoyl reductase | Enoyl reduction at C10=C11 or C12=C13 |
| FSL6 (FGSG_10459) | AMP-binding | Unknown role or not involved in biosynthesis |
| FSL7 (FGSG_10458) | Transcription factor | Transcriptional regulation of the fusarielin gene cluster |
| Pos. | 1H Shift (ppm) 1 | 13C Shift (ppm) | Pos. | 1H Shift (ppm) 1 | 13C Shift (ppm) |  |
| 1 | 176.40 | 13 | 1.69, 1H, dd | 37.46 | ||
| 2 | 2.38, 1H, dq | 43.53 | 1.99, 1H, dd | |||
| 3 | 3.92, 1H, d | 78.80 | 14 | 1.90, 1H, dddd | 38.06 | |
| 4 | 135.53 | 15 | 5.36, 1H, s | 126.62 | ||
| 5 | 5.90, 1H, d | 126.89 | 16 | 134.05 | ||
| 6 | 5.32, 1H, d | 136.34 | 17 | 2.58, 1H, d | 54.10 | |
| 7 | 6.24, 1H, dd | 126.09 | 18 | 134.60 | ||
| 8 | 2.21, 1H, ddd | 47.73 | 19 | 5.16, 1H, q | 122.07 | |
| 9 | 1.41, 1H, dddd | 34.02 | 20 | 1.57, 3H, d | 13.38 | |
| 10 | 1.44, 1H, ddd | 30.91 | 21 | 0.81, 3H, d | 13.99 | |
| 1.82, 1H, ddd | 22 | 1.62, 3H, s | 11.01 | |||
| 11 | 5.34, 1H, dd | 121.31 | 23 | 1.62, 3H, s | 23.12 | |
| 12 | 133.31 | 24 | 1.54, 3H, s | 18.01 | ||
| 25 | 1.51, 3H, s | 21.88 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Droce, A.; Saei, W.; Jørgensen, S.H.; Wimmer, R.; Giese, H.; Wollenberg, R.D.; Sondergaard, T.E.; Sørensen, J.L. Functional Analysis of the Fusarielin Biosynthetic Gene Cluster. Molecules 2016, 21, 1710. https://doi.org/10.3390/molecules21121710
Droce A, Saei W, Jørgensen SH, Wimmer R, Giese H, Wollenberg RD, Sondergaard TE, Sørensen JL. Functional Analysis of the Fusarielin Biosynthetic Gene Cluster. Molecules. 2016; 21(12):1710. https://doi.org/10.3390/molecules21121710
Chicago/Turabian StyleDroce, Aida, Wagma Saei, Simon Hartung Jørgensen, Reinhard Wimmer, Henriette Giese, Rasmus Dam Wollenberg, Teis Esben Sondergaard, and Jens Laurids Sørensen. 2016. "Functional Analysis of the Fusarielin Biosynthetic Gene Cluster" Molecules 21, no. 12: 1710. https://doi.org/10.3390/molecules21121710