
Isolation of Flavonoids from Deguelia duckeana and Their Effect on Cellular Viability, AMPK, eEF2, eIF2 and eIF4E

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
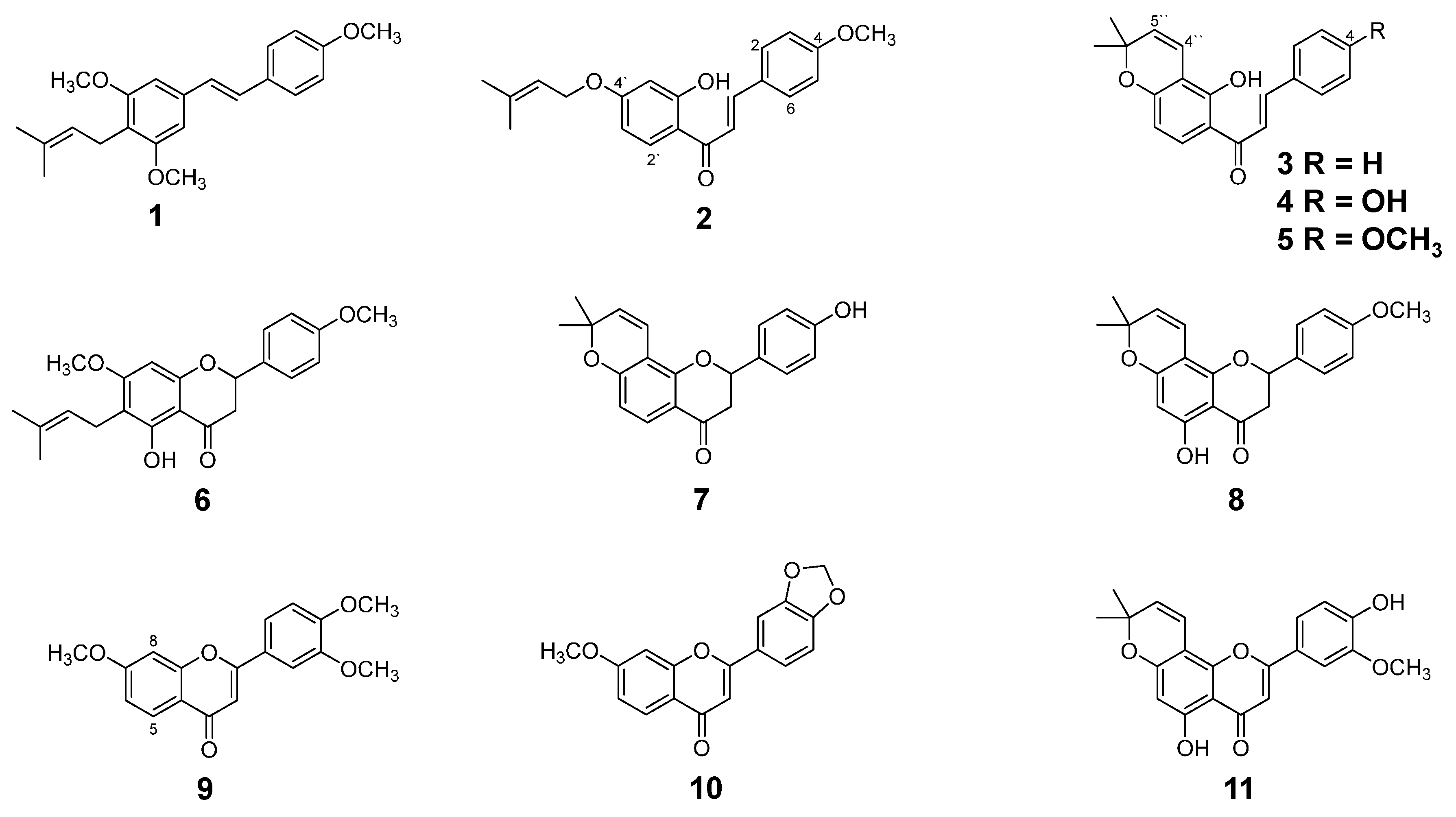
2.1. Isolation and Identification of Flavonoids from D. duckeana Including a New Flavanone


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | δC (ppm) | δH (ppm) |
|---|---|---|
| 2 | 79.1 | 5.34 (dd, 13.0 and 3.0) |
| 3 | 43.3 | 2.76 (dd, 17.2 and 3.0) a and 3.09 (dd, 17.2 and 13.0) b |
| 4 | 196.0 | - |
| 5 | 160.0 | - |
| 6 | 109.9 | - |
| 7 | 165.4 | - |
| 8 | 90.9 | 6.07 (1H, s) |
| 9 | 161.4 | - |
| 10 | 102.9 | - |
| 1’ | 130.3 | - |
| 2’/6’ | 127.7 | 7.38 (d, 8.5) |
| 3’/5’ | 114.2 | 6.95 (d, 8.5) |
| 4’ | 160.2 | - |
| 1” | 21.0 | |
| 2” | 122.2 | 5.19 (1H, m) |
| 3” | 131.6 | |
| 4” | 25.8 | 1.76 (3H, s) |
| 5” | 17.7 | 1.81 (3H, s) |
| 5-OH | 160.2 | 12.06 (1H, s) |
| 7-OCH3, 4’-OCH3 | 55.3 | 3.83 (6H, s) |
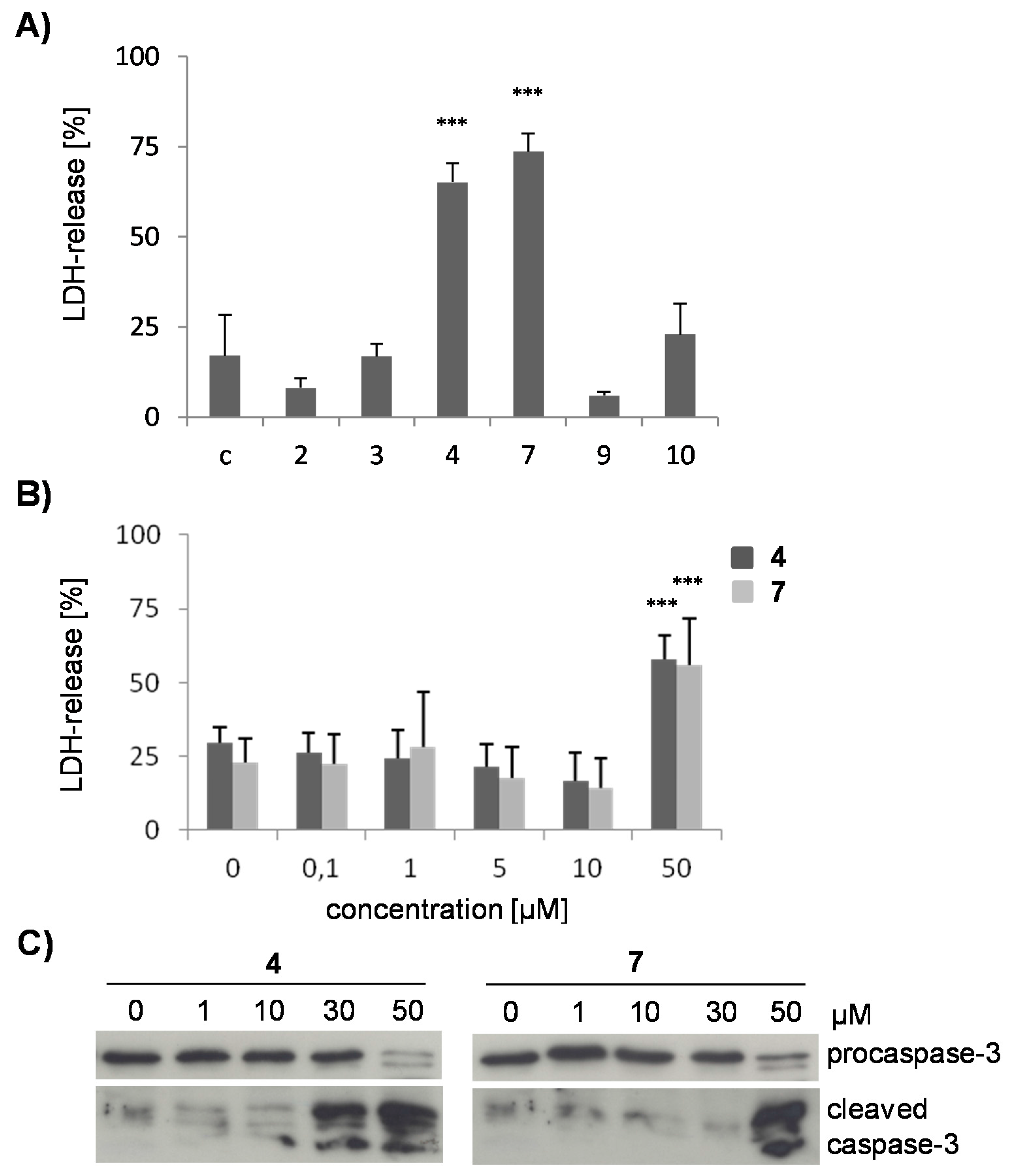
2.2. The Flavonoids from D. duckeana Differently Influence Cell Viability


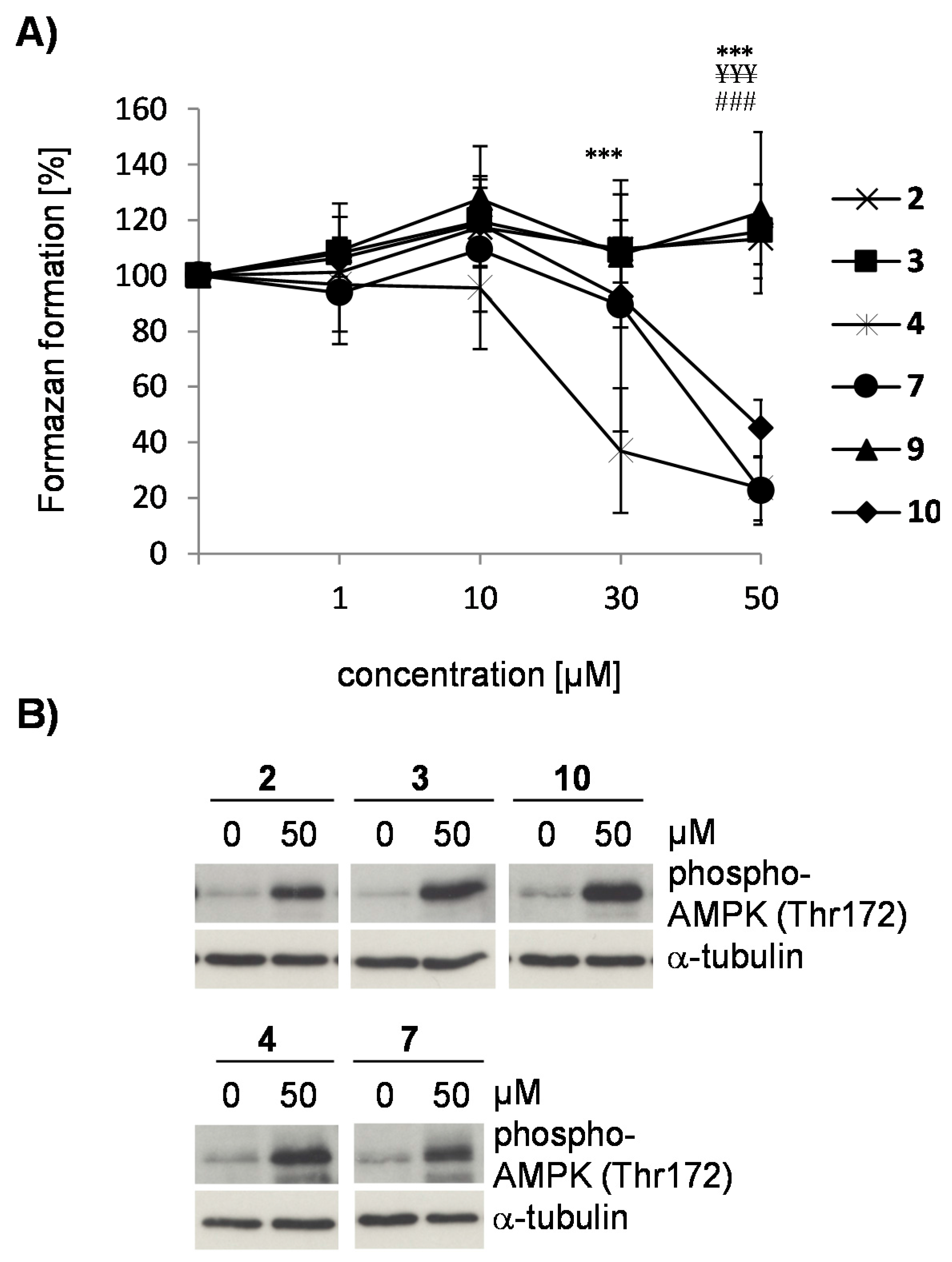
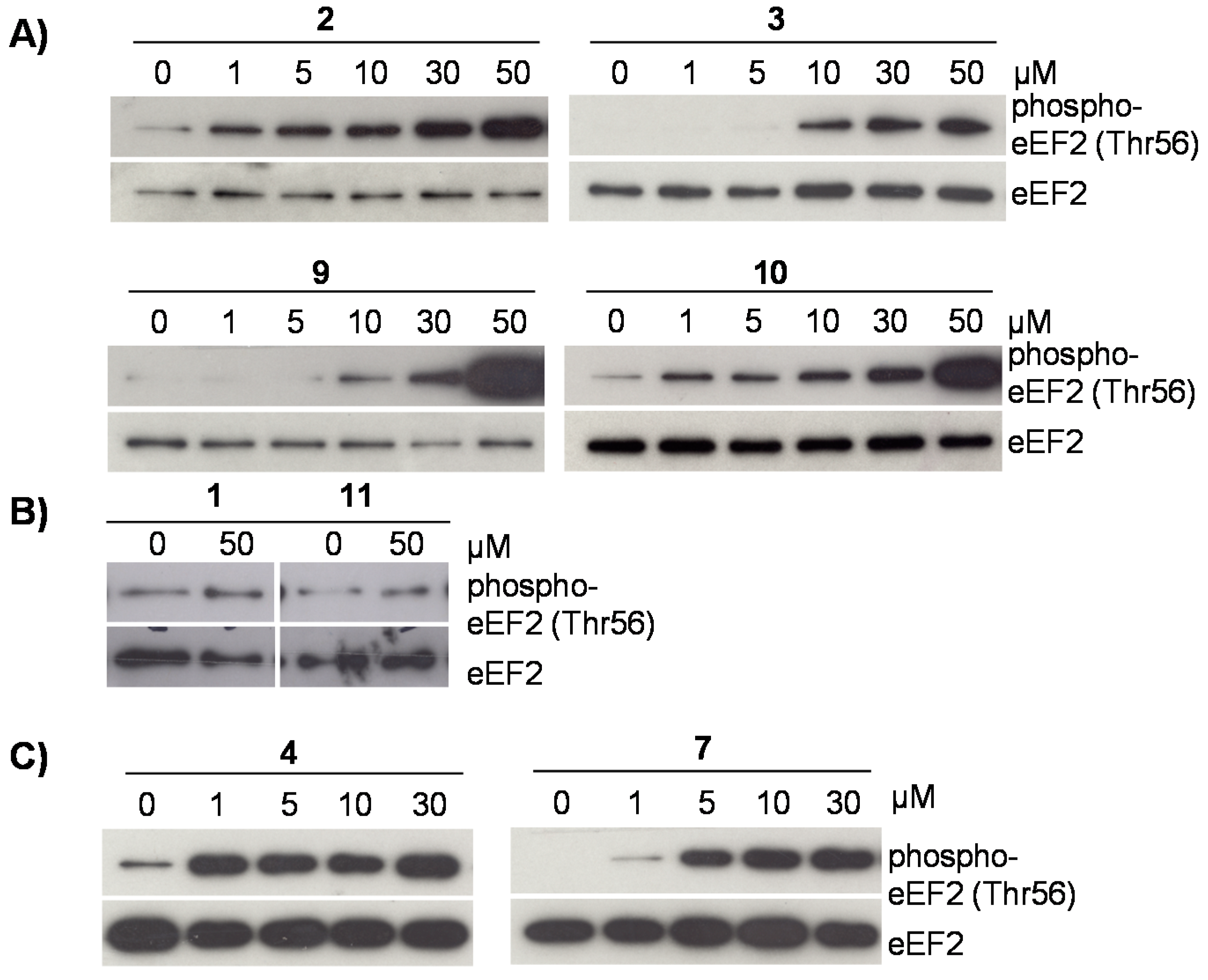
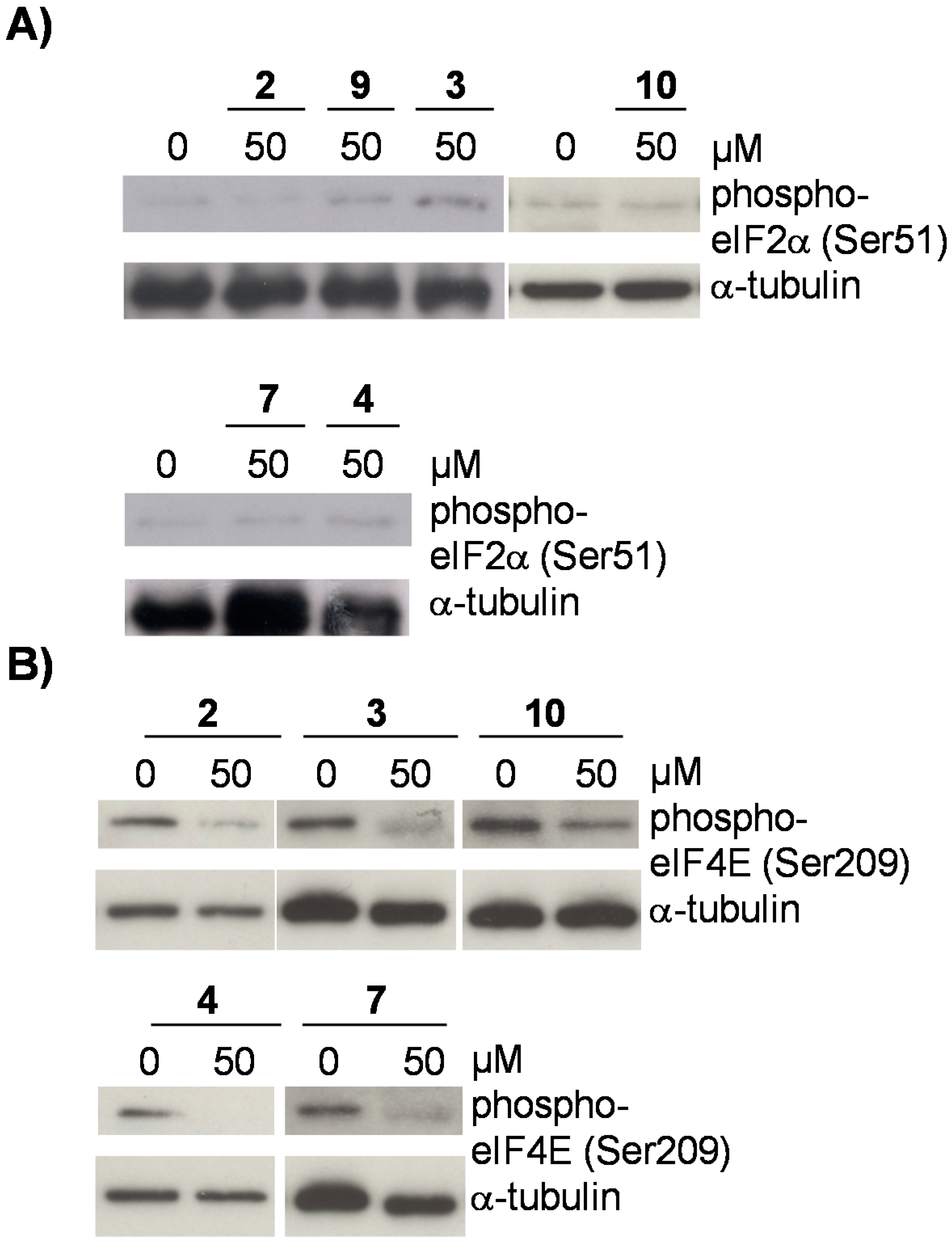
2.3. Flavonoids from D. duckeana Induce the Phosphorylation of AMPK and eEF2, But Not eIF2alpha


2.4. Conclusions
3. Experimental Section
3.1. General Procedures
3.2. Plant Material
3.3. Extraction and Isolation
3.4. Cell Culture and Treatment
3.5. Cytotoxicity Assay
3.6. Metabolic Activity
3.7. Immunoblot Analysis
3.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ribeiro, D.; Freitas, M.; Lima, F.J.F.C.; Fernandes, E. Proinflammatory pathways: The modulation by flavonoids. Rev. Med. Res. 2015, 35, 877–936. [Google Scholar] [CrossRef] [PubMed]
- Nijveldt, R.J.; Nood, E.; Hoorn, D.E.C.; Boelen, P. G.; Norren, K.; Leeuwen, P.A.M. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [PubMed]
- Kuete, V.; Ngameni, B.; Wiench, B.; Krusche, B.; Horwedel, C.; Ngadjui, B.T.; Efferth, T. Cytotoxicity and mode of action of four naturally occurring flavonoids from the genus Dorstenia: Gancaonin Q, 4-hydroxylonchocarpin, 6-prenylapigenin and 6,8-diprenyleriodictyol. Planta Med. 2011, 77, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Nkuete, A.H.L.; Mbaveng, A.T.; Wiench, B.; Wabo, H.K.; Tane, P.; Efferth, T. Cytotoxicity and modes of action of 4-hydroxy-2,6-dimethoxychalcone and other flavonoids toward drug-sensitive and multidrug-resistant cancer cell lines. Phytomedicine 2014, 21, 1651–1657. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.F.; Tozzi, A.M.G.A.; Magalhães, E.V.; Moraes, V.R.S. Prenylated flavonoids from Deguelia hatschbachii and their systematic significance in Deguelia. Phytochemistry 2001, 57, 77–89. [Google Scholar] [CrossRef]
- Braz Filho, R.; Gottlieb, O.R.; Mourão, A.P.; Rocha, A.I.; Oliveira, F.S. Flavonoids from Derris species. Phytochem. Rep. 1975, 14, 1454–1456. [Google Scholar] [CrossRef]
- Magalhães, A.F.; Tozzi, A.M.G.A.; Magalhães, E.G.; Moraes, V.R.S. New spectral data of some flavonoids from Deguelia hatschbachii A.M.G. Azevedo. J. Braz. Chem. Soc. 2003, 14, 133–137. [Google Scholar] [CrossRef]
- Lobo, L.V.; Silva, G.A.; Freitas, M.C.C; Souza Filho, A.P.S.; Silva, M.N.; Arruda, A.C.; Guilhon, G.M.S.P.; Santos, L.S.; Santos, A.S.; Arruda, M.S.P. Stilbenes from Deguelia rufescens var. urucu (Ducke) A.M.G. Azevedo leaves: Effects on seed germination and plant growth. J. Braz. Chem. Soc. 2010, 21, 1838–1844. [Google Scholar] [CrossRef]
- Magalhães, A.F.; Tozzi, A.M.G.A.; Magalhães, E.G.; Souza-Neta, L.C. New prenylated metabolites of Deguelia longeracemosa and evaluation of their antimicrobial potential. Planta Med. 2006, 72, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.G.; Almeida, C.M.C.; Silva, C.Y.Y.; Arruda, M.S.P.; Arruda, A.C.; Lopes, D.C.F.; Yamada, E.S.; Costa, E.T.; Silva, M.N. Flavonoids from the leaves of Deguelia utilis (Leguminosae): Structural elucidation and neuroprotective properties. J. Braz. Chem. Soc. 2012, 23, 1933–1939. [Google Scholar] [CrossRef]
- Lima, N.M.; Andrade, J.I.A.; Lima, K.C.S.; Santos, F.N.; Barison, A.; Salomé, K.S.; Matsuura, T.; Nunez, C.V. Chemical profile and biological activities of Deguelia duckeana A.M.G. Azevedo (Fabaceae). Nat. Prod. Res. 2013, 27, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Dagne, E.; Bekele, A.; Waterman, P.G. The flavonoids of Millettia ferruginosa and subsp. Darassana in Ethiopia. Phytochemistry 1989, 28, 1897–1900. [Google Scholar] [CrossRef]
- Jang, D.S.; Cuendet, M.; Hawthorne, M.E.; Kardono, L.B.S.; Kawanishi, K.; Fong, H.H.S.; Mehta, R.G.; Pezzuto, J.M.; Kinghorn, A.D. Prenylated flavonoids of the leaves of Macaranga conifera with inhibitory activity against cyclooxygenase-2. Phytochemistry 2002, 61, 867–872. [Google Scholar] [CrossRef]
- Mahmoud, E.N.; Waterman, G. Flavonoids from the stem bark of Millettia hemsleyana. Phytochemistry 1985, 24, 369–371. [Google Scholar] [CrossRef]
- Banerji, A.; Luthria, D.L.; Prabhu, B.R. Prenylated compounds from Atalantia racemosa: Isolation and synthesis of two pyranoflavones. Phytochemistry 1988, 27, 3637–3640. [Google Scholar] [CrossRef]
- Kato, M.J.; Yoshida, M.; Gottlieb, O.R. Flavones and lignans in flowers, fruits and seedlings of Virola venosa. Phytochemistry 1992, 31, 283–287. [Google Scholar] [CrossRef]
- Intekhab, J.; Mohammad, A. Isolation of a flavonoid from Feronia limonia. J. Saudi Chem. Soc. 2009, 13, 295–298. [Google Scholar] [CrossRef]
- Jain, A.C.; Khazanchi, R.; Kumar, A. Synthesis of carpachromene and related isopentenylated derivatives of apigenin. Tetrahedron 1978, 34, 3569–3573. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hardie, D.G. The AMP-activated protein kinase pathway-new players upstream and downstream. J. Cell Sci. 2004, 117, 5479–5487. [Google Scholar] [CrossRef] [PubMed]
- Sanz, P. AMP-Activated protein kinase: Structure and regulation. Curr. Protein Pept. Sci. 2008, 9, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Horman, S.; Browne, G.; Krause, U.; Patel, J.; Vertommen, D.; Bertrand, L.; Lavoinne, A.; Hue, L.; Proud, C.; Rider, M. Activation of AMP-activated protein kinase leads to the phosphorylation of elongation factor 2 and an inhibition of protein synthesis. Curr. Biol. 2002, 12, 1419–1423. [Google Scholar] [CrossRef]
- Stein, S.C.; Woods, A.; Jones, N.A.; Davison, M.D.; Carling, D. The regulation of AMP-activated protein kinase by phosphorylation. Biochem. J. 2000, 345, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Mamane, Y.; Petroulakis, E.; Rong, L; Yoshida, K.; Ler, L.W.; Sonnenberg, N. eIF4E—From translation to transformation. Oncogene 2004, 23, 3172–3179. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cursino, L.M.C.; Lima, N.M.; Murillo, R.; Nunez, C.V.; Merfort, I.; Humar, M. Isolation of Flavonoids from Deguelia duckeana and Their Effect on Cellular Viability, AMPK, eEF2, eIF2 and eIF4E. Molecules 2016, 21, 192. https://doi.org/10.3390/molecules21020192
Cursino LMC, Lima NM, Murillo R, Nunez CV, Merfort I, Humar M. Isolation of Flavonoids from Deguelia duckeana and Their Effect on Cellular Viability, AMPK, eEF2, eIF2 and eIF4E. Molecules. 2016; 21(2):192. https://doi.org/10.3390/molecules21020192
Chicago/Turabian StyleCursino, Lorena M. C., Nerilson M. Lima, Renato Murillo, Cecilia V. Nunez, Irmgard Merfort, and Matjaz Humar. 2016. "Isolation of Flavonoids from Deguelia duckeana and Their Effect on Cellular Viability, AMPK, eEF2, eIF2 and eIF4E" Molecules 21, no. 2: 192. https://doi.org/10.3390/molecules21020192