
Antimicrobial Activity and Stability of Short and Long Based Arachnid Synthetic Peptides in the Presence of Commercial Antibiotics


Abstract
:1. Introduction
2. Results
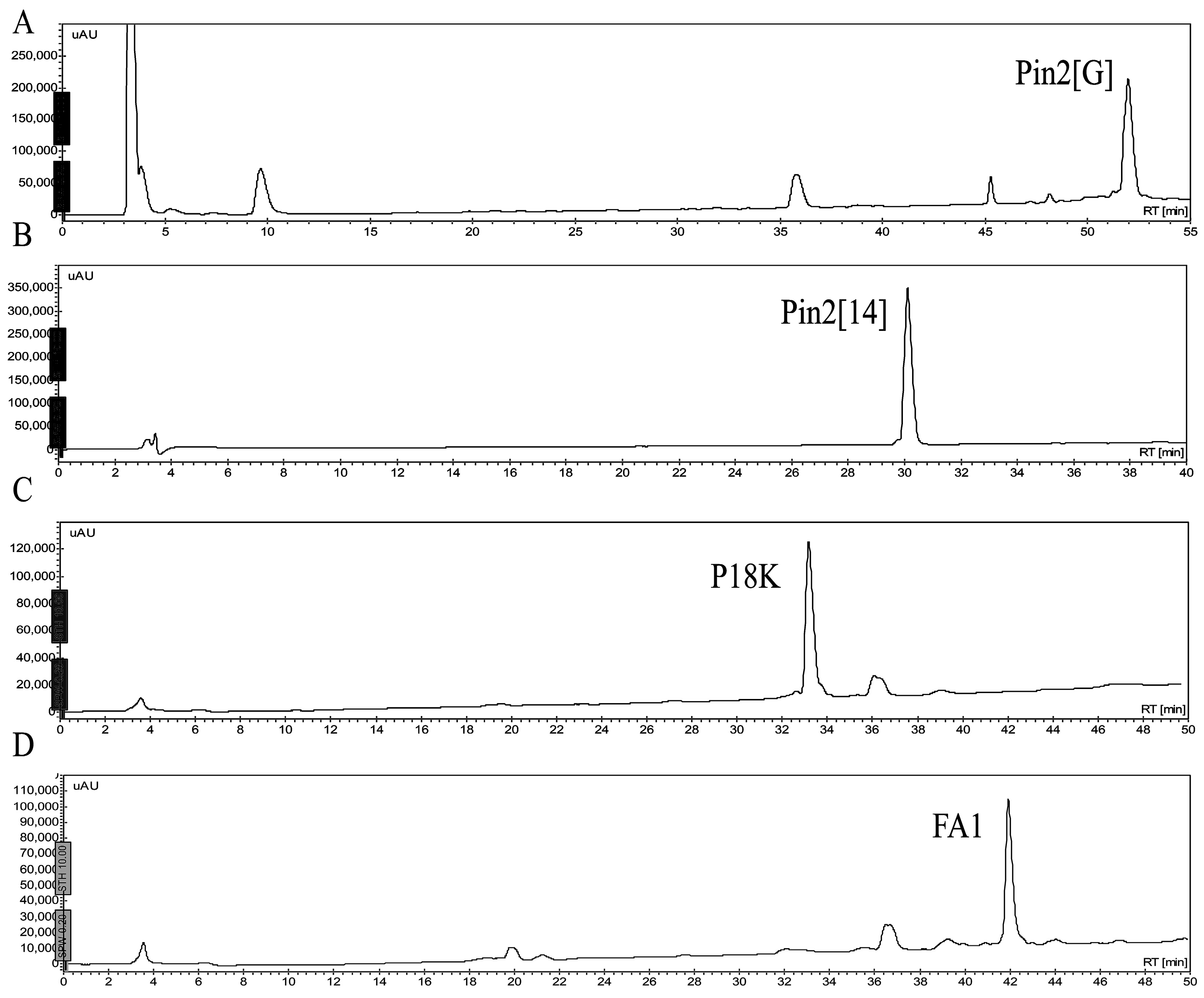
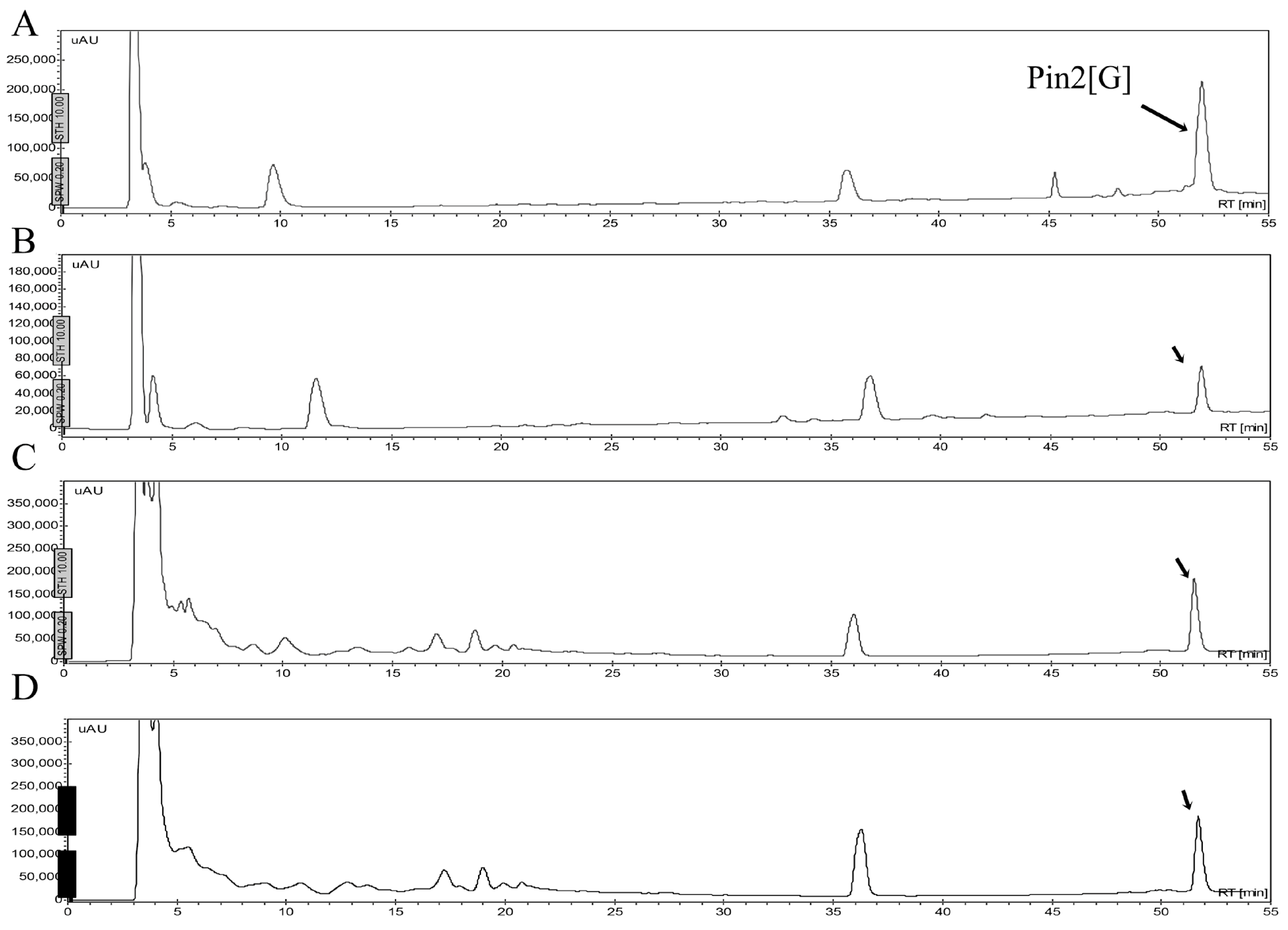
2.1. Isolation and Sequence Analysis
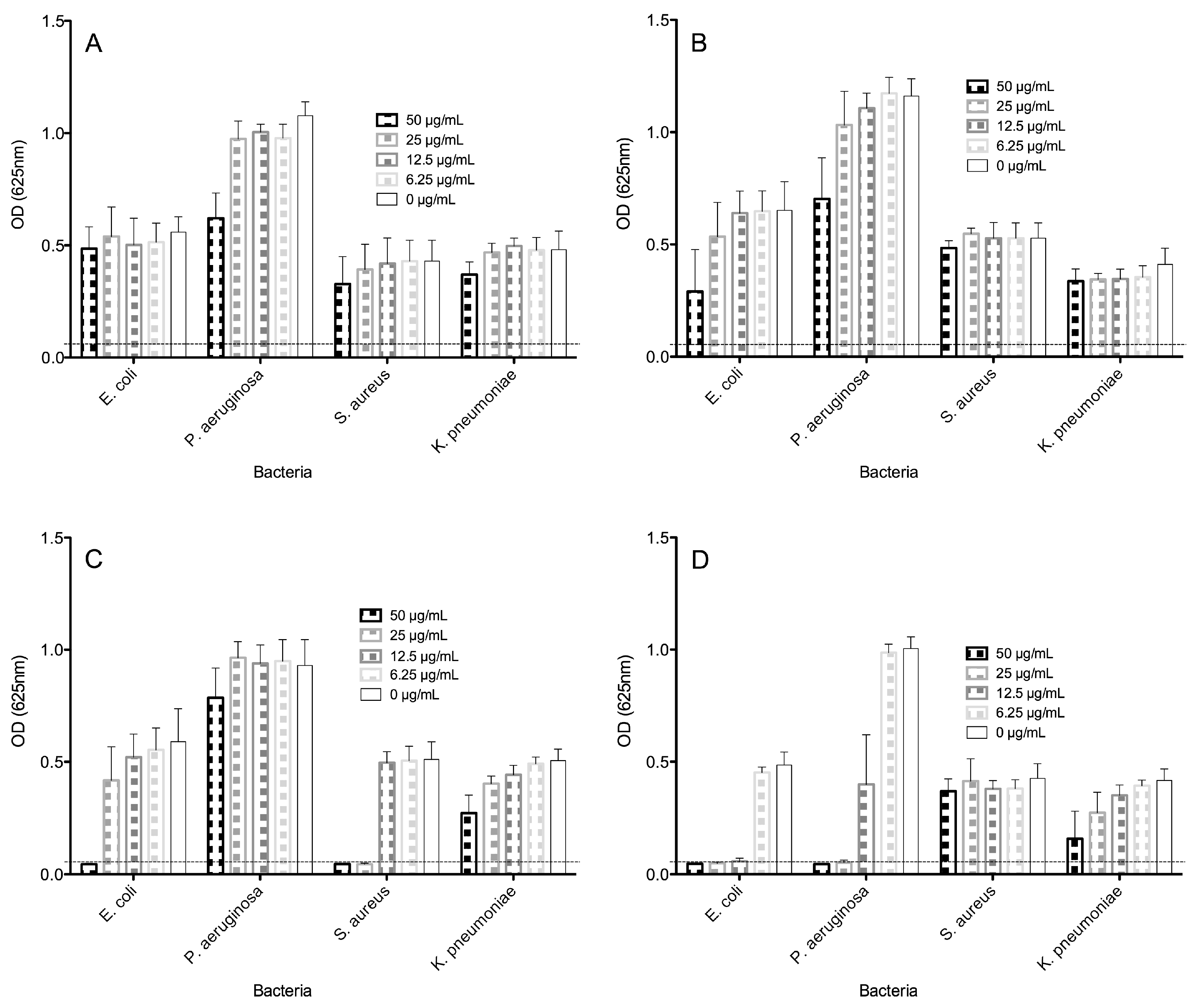
2.2. Antimicrobial Activities of Peptides
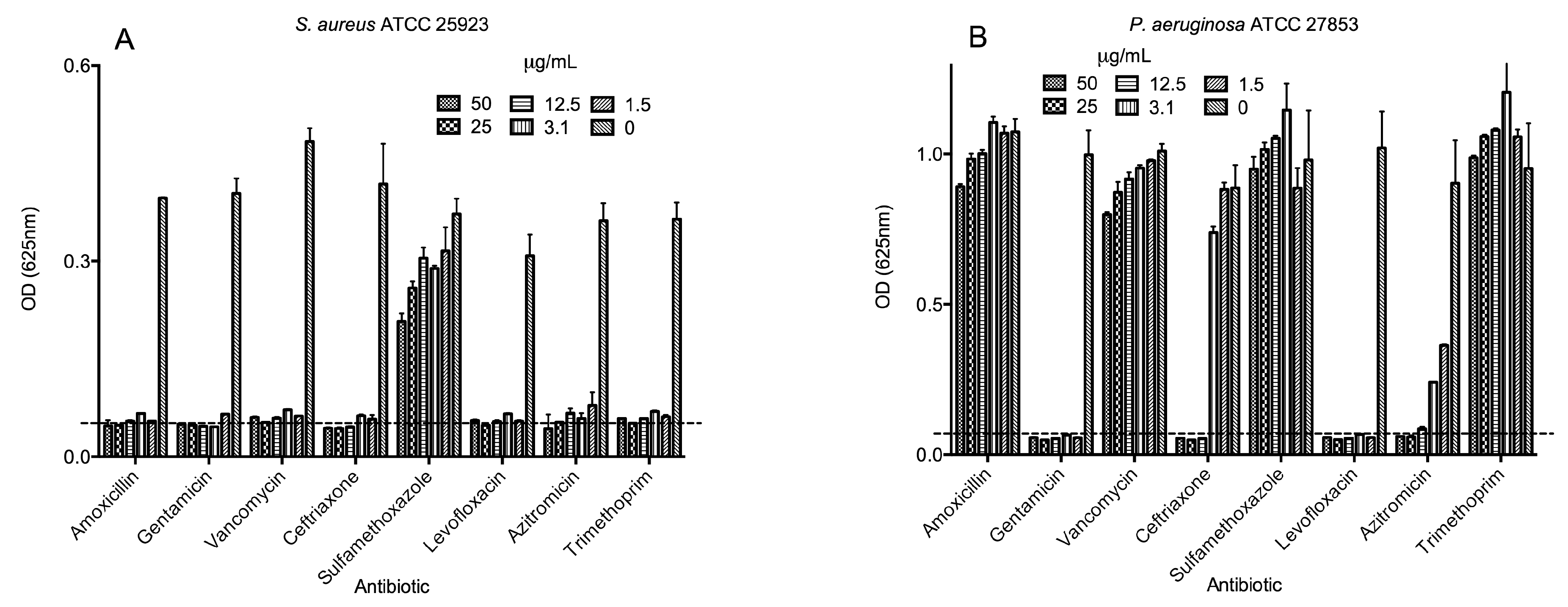
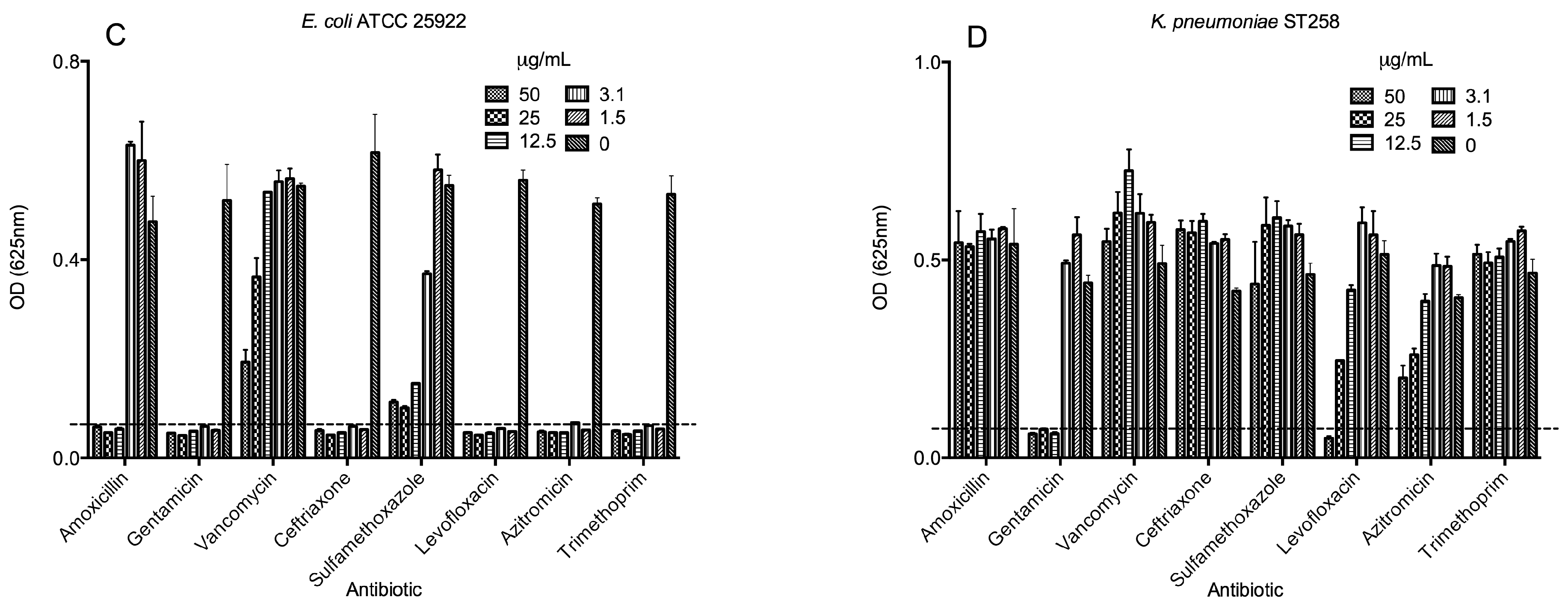
2.3. Antimicrobial Activities of Selected Antibiotics
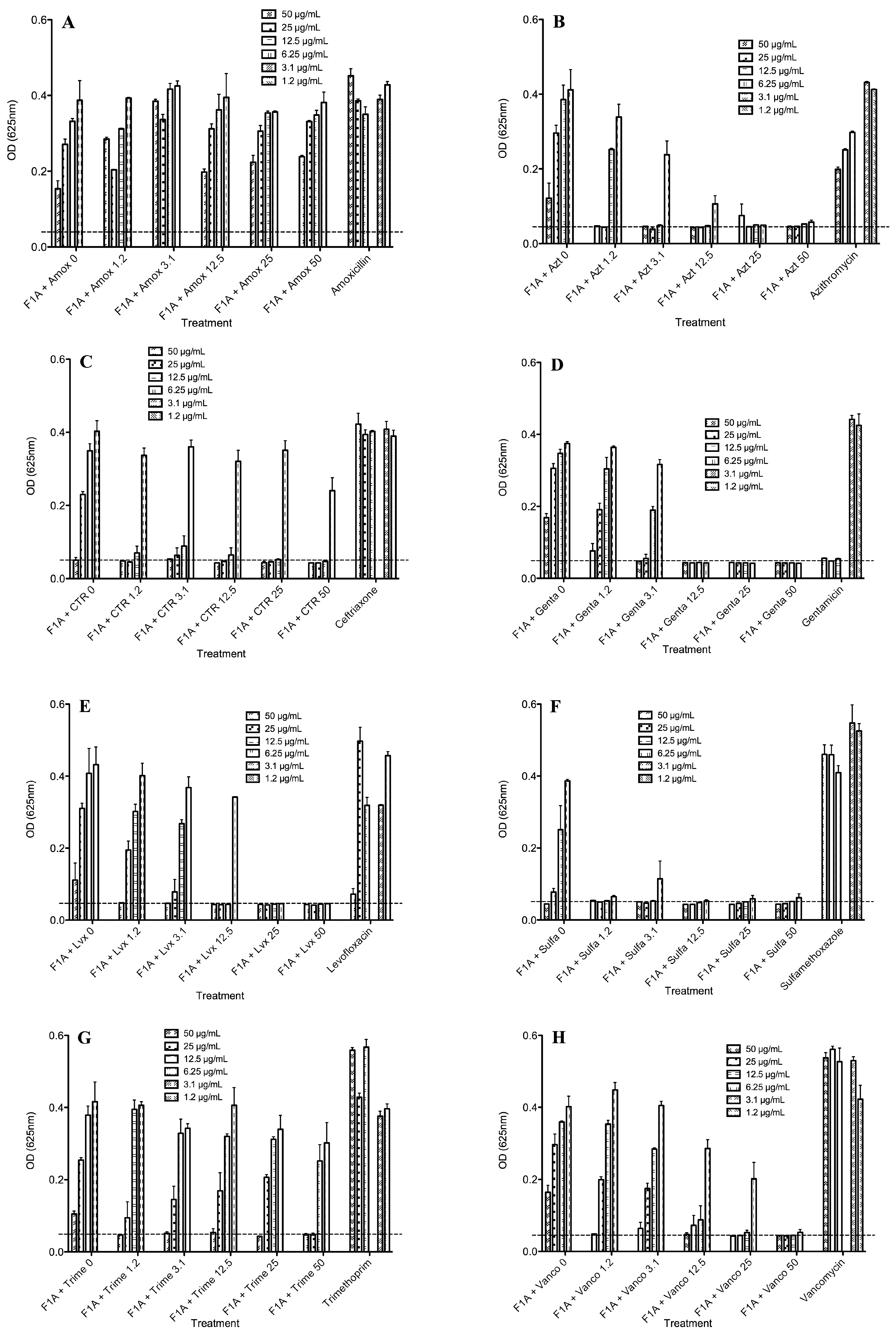
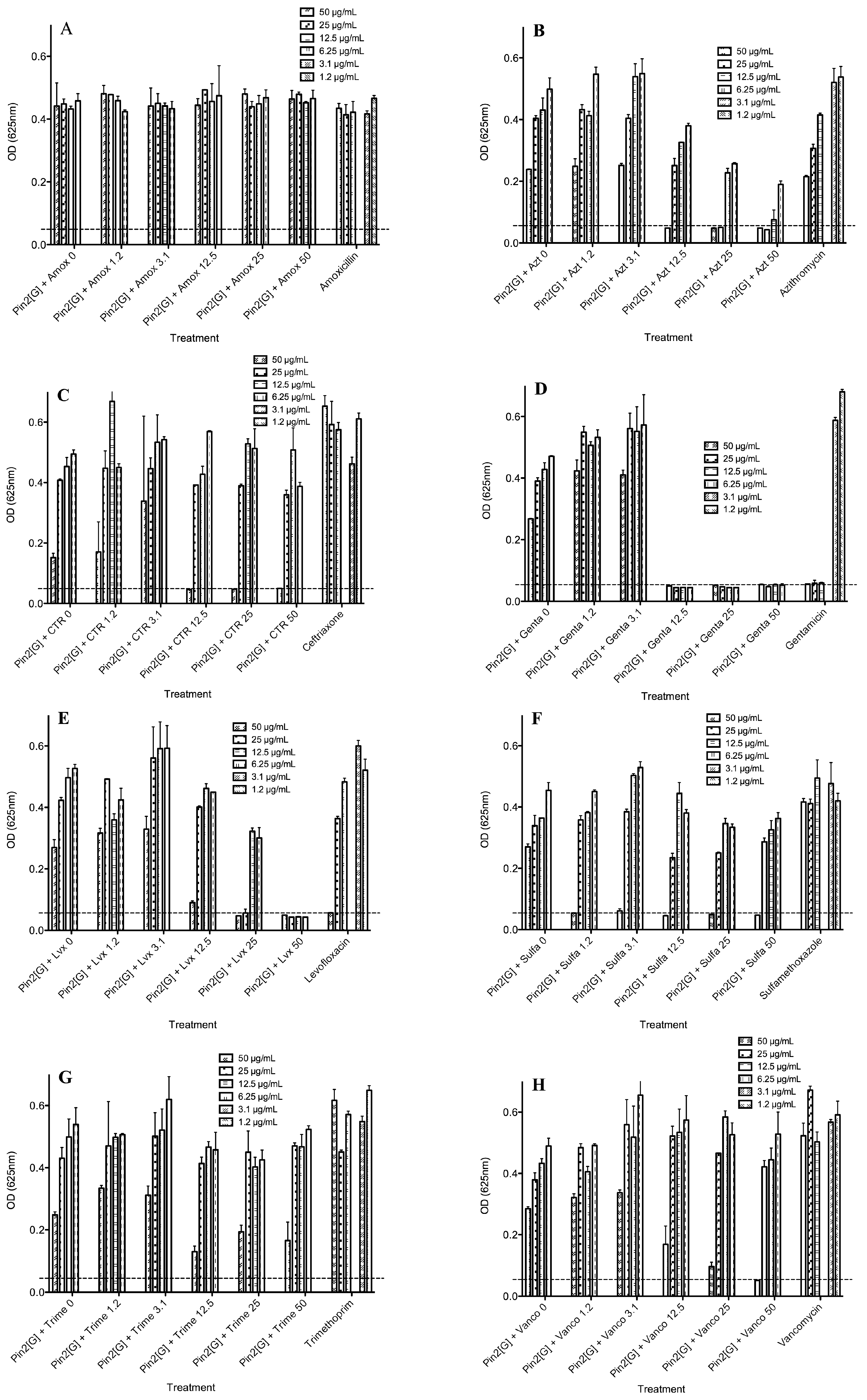
2.4. Biological Activities of Selected Peptides and Antibiotics
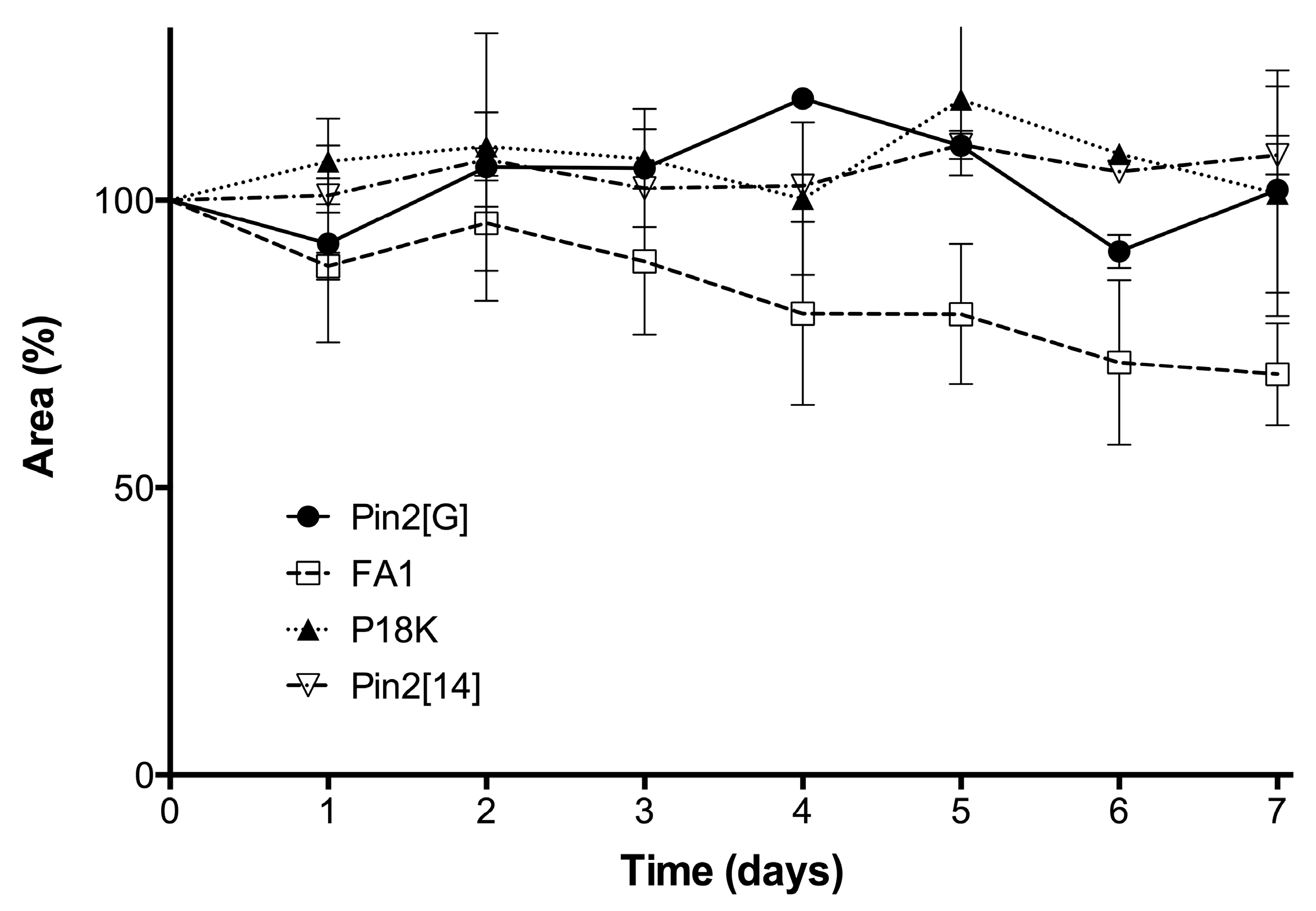
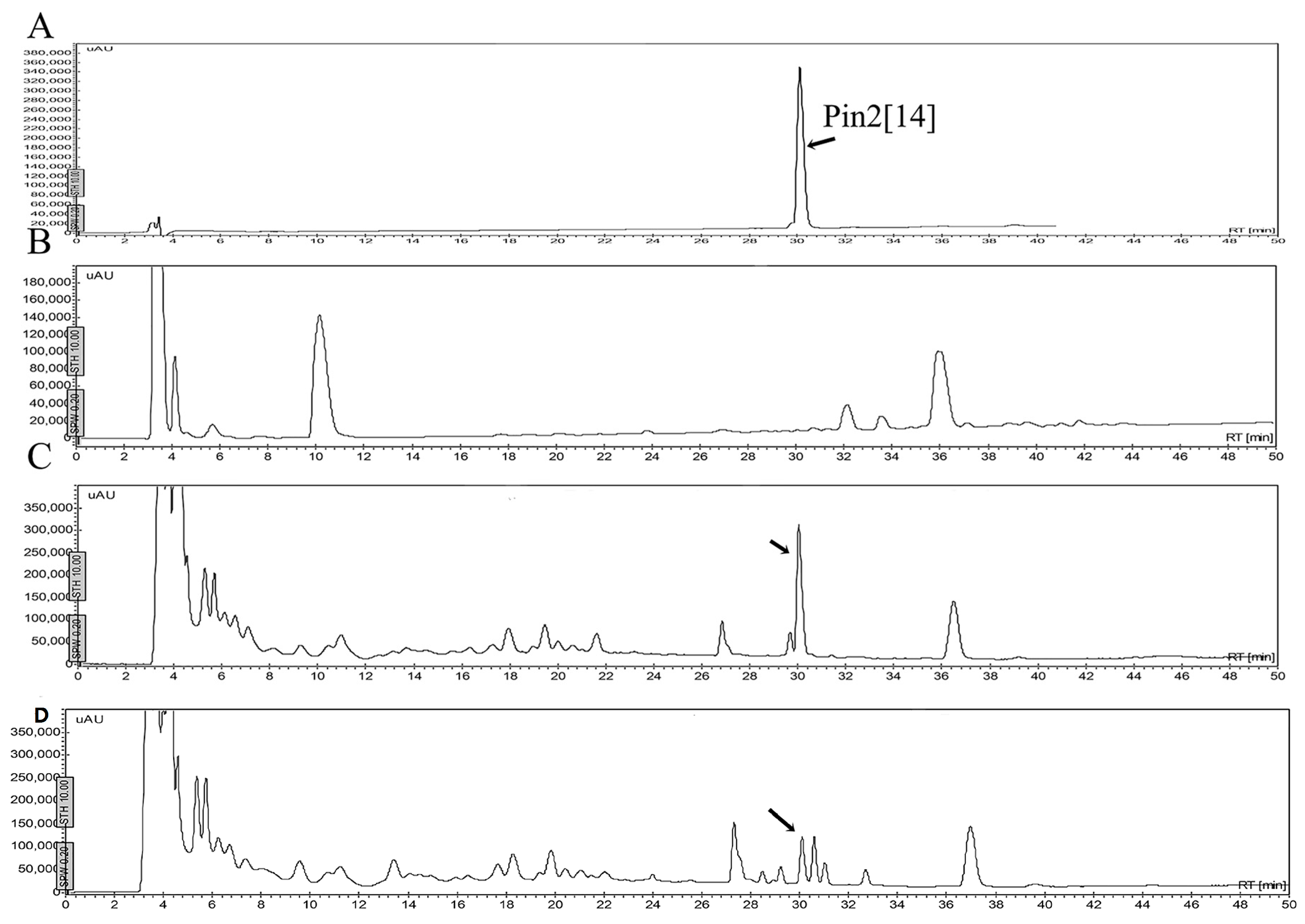
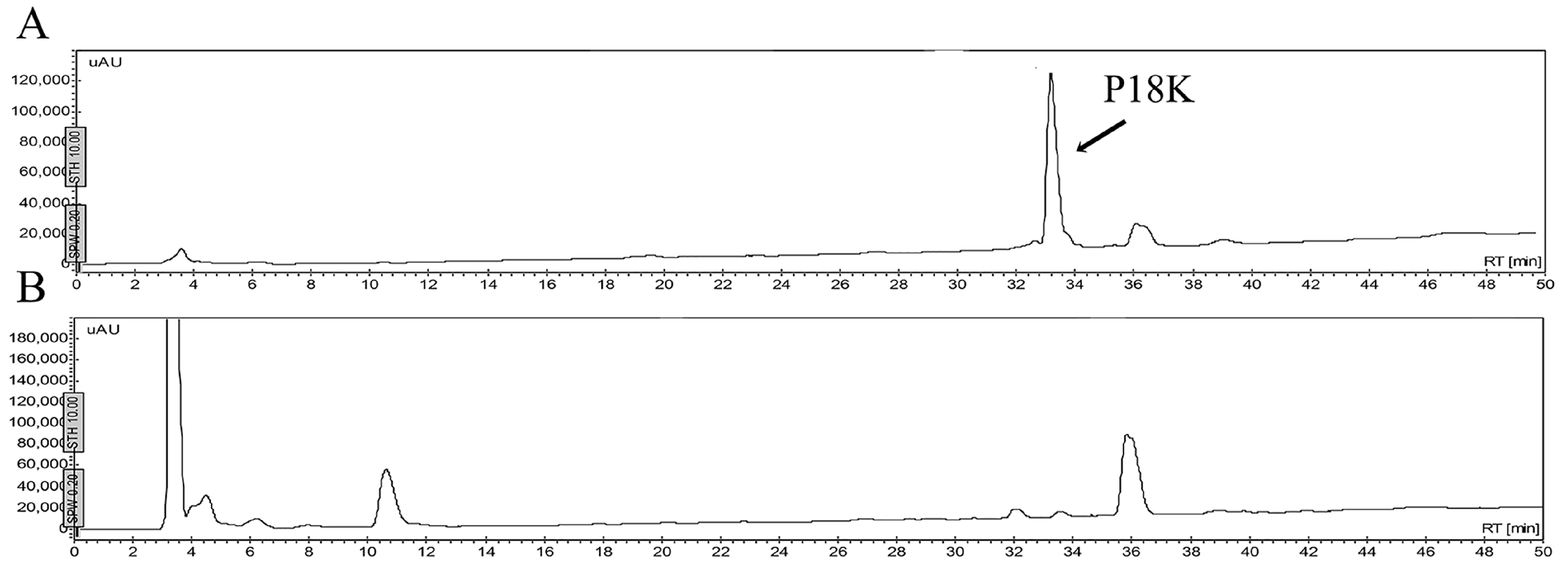
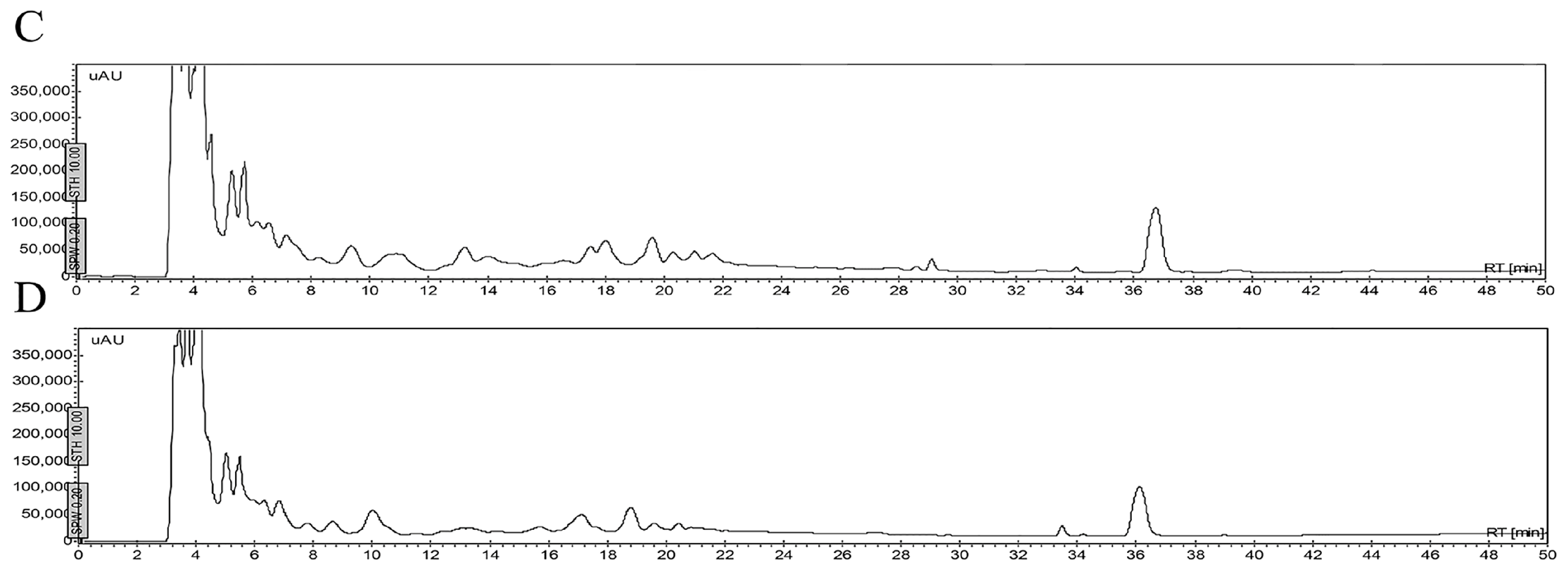
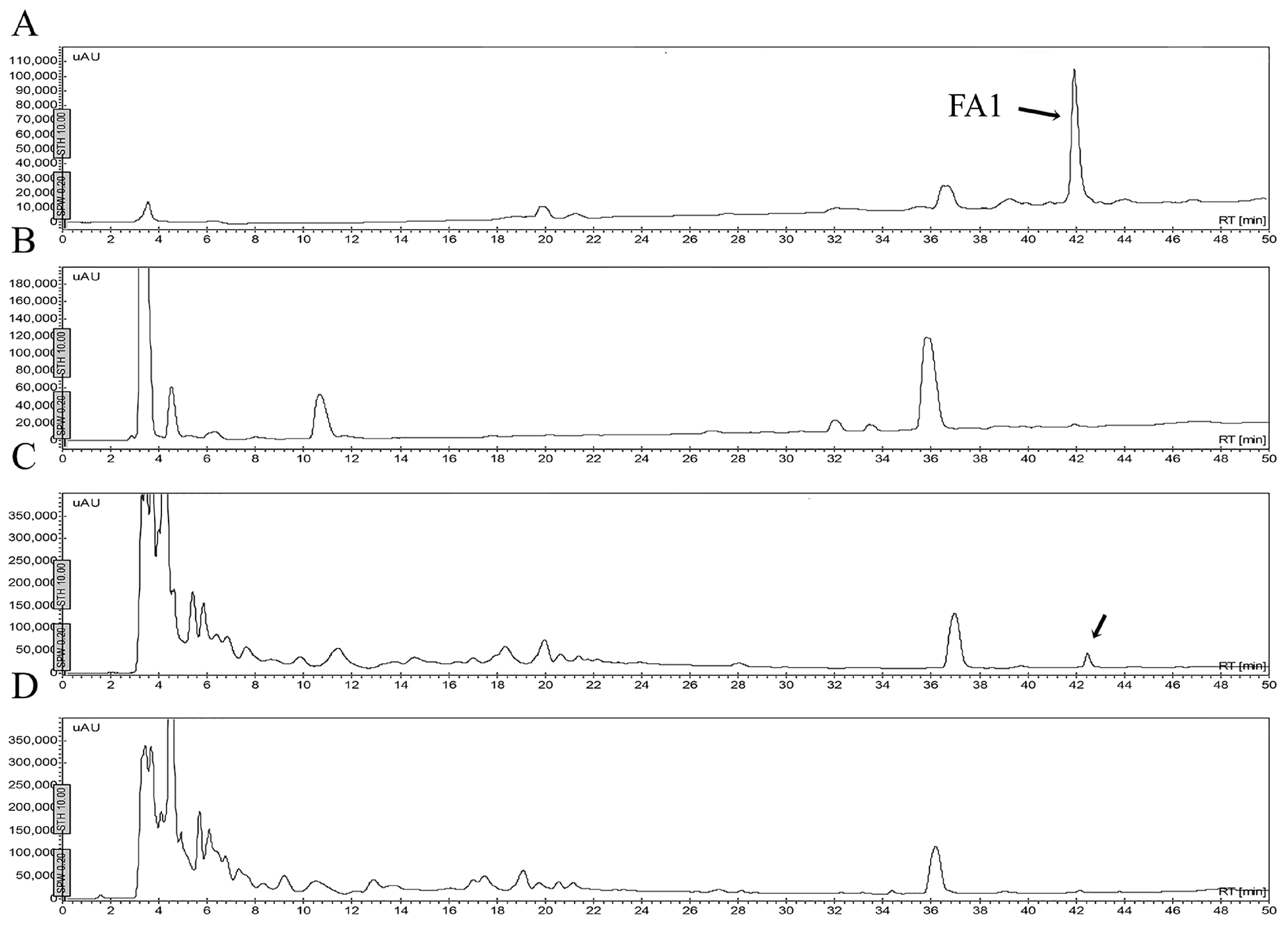
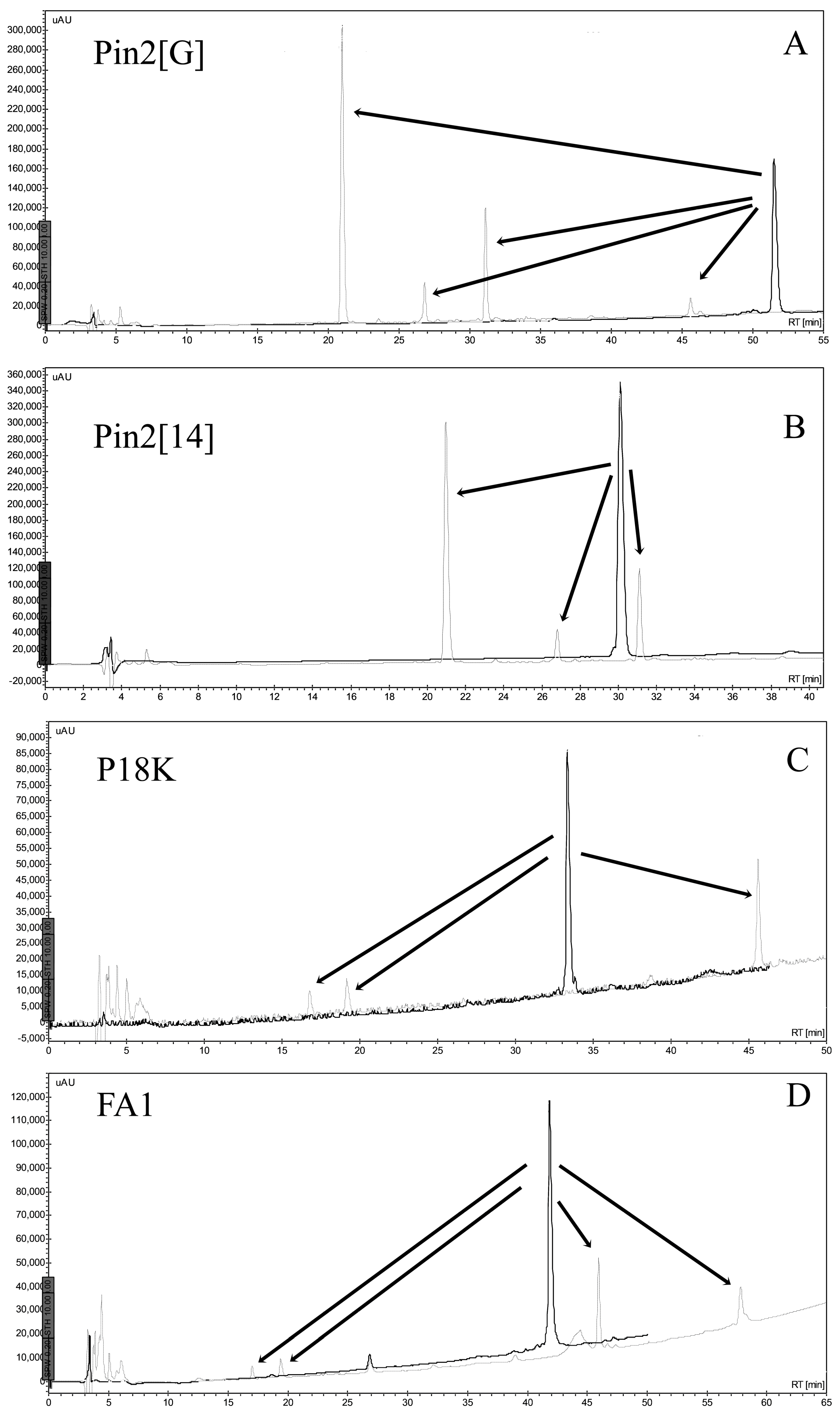
2.5. Stability of Peptides in Saline Buffer, Human Blood Plasma and in the Presence of Enzymes
3. Discussion
4. Materials and Methods
4.1. Biologicals
4.2. Peptide Synthesis and Purification
4.3. Sequence Analysis and Mass Spectrometry
4.4. Antimicrobial Activity
4.5. Antibiotic Analysis
4.6. Stability of Peptides
4.6.1. Stability in Saline Buffer
4.6.2. Stability in Human Blood Plasma and in the Supernatant of Bacterial Growth
4.6.3. Stability in the Presence of Enzymes
4.7. Statistical Analysis
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Verhoef, J. Antibiotic resistance: The pandemic. Adv. Exp. Med. Biol. 2003, 531, 301–313. [Google Scholar] [PubMed]
- Gould, I.M.; Bal, A.M. New antibiotic agents in the pipeline and how they can help overcome microbial resistance. Virulence 2013, 4, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.; Villegas, E.; Espino-Solis, G.P.; Rodriguez, A.; Paniagua-Solis, J.F.; Sandoval-Lopez, G.; Possani, L.D.; Corzo, G. Antimicrobial peptides from arachnid venoms and their microbicidal activity in the presence of commercial antibiotics. J. Antibiot. 2013, 66, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Carreto, S.; Jimenez-Vargas, J.M.; Rivas-Santiago, B.; Corzo, G.; Possani, L.D.; Becerril, B.; Ortiz, E. Peptides from the scorpion vaejovis punctatus with broad antimicrobial activity. Peptides 2015, 73, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.e.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the expasy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Liu, L.; Fang, Y.; Huang, Q.; Pan, Q.; Wu, J. A new structure-activity relationship of linear cationic α-helical antimicrobial peptides. In 7th Asian-Pacific Conference on Medical and Biological Engineering; Peng, Y., Weng, X., Eds.; Springer: Berlin, Germany; Heidelberg, Germany, 2008; Vol. 19, pp. 167–170. [Google Scholar]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. Heliquest: A web server to screen sequences with specific alpha-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef] [PubMed]
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. Comparison of three different in vitro methods of detecting synergy: Time-kill, checkerboard, and e test. Antimicrob. Agents Chemother. 1996, 40, 1914–1918. [Google Scholar] [PubMed]
- Zureik, M.; Robert, L.; Courbon, D.; Touboul, P.J.; Bizbiz, L.; Ducimetiere, P. Serum elastase activity, serum elastase inhibitors, and occurrence of carotid atherosclerotic plaques: The etude sur le vieillissement arteriel (eva) study. Circulation 2002, 105, 2638–2645. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Lehrer, R.I. Cationic peptides: A new source of antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Yeaman, R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Melo, M.N.; Castanho, M.A. Cell-penetrating peptides and antimicrobial peptides: How different are they? Biochem. J. 2006, 399, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.M.; Edwards, M.A.; Li, J.; Yip, C.M.; Deber, C.M. Roles of hydrophobicity and charge distribution of cationic antimicrobial peptides in peptide-membrane interactions. J. Biol. Chem. 2012, 287, 7738–7745. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Villegas, E.; Montoya-Rosales, A.; Rivas-Santiago, B.; Corzo, G. Characterization of antibacterial and hemolytic activity of synthetic pandinin 2 variants and their inhibition against mycobacterium tuberculosis. PLoS ONE 2014, 9, e101742. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vasquez, L.; Silva-Sanchez, J.; Jimenez-Vargas, J.M.; Rodriguez-Romero, A.; Munoz-Garay, C.; Rodriguez, M.C.; Gurrola, G.B.; Possani, L.D. Enhanced antimicrobial activity of novel synthetic peptides derived from vejovine and hadrurin. Biochim. Biophys. Acta 2013, 1830, 3427–3436. [Google Scholar] [CrossRef] [PubMed]
- Carmona, G.; Rodriguez, A.; Juarez, D.; Corzo, G.; Villegas, E. Improved protease stability of the antimicrobial peptide pin2 substituted with d-amino acids. Protein J. 2013, 32, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Corzo, G.; Escoubas, P.; Villegas, E.; Barnham, K.J.; He, W.; Norton, R.S.; Nakajima, T. Characterization of unique amphipathic antimicrobial peptides from venom of the scorpion pandinus imperator. Biochem. J. 2001, 359, 35–45. [Google Scholar] [CrossRef] [PubMed]
- CLSI, C.A.L.S.I. Performance standards for antimicrobial susceptibility testing. In NCCLS Document 31M100-S21; Standards, N.C.C.L., Ed.; National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2011. [Google Scholar]
- Sample Availability: Samples of the compounds Pin2[G], Pin2[14], P18K and FA1 are available from the authors.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequences | GRAVY | AMF | µH | Q | RT (min) | Molecular Mass (Da) | |
|---|---|---|---|---|---|---|---|---|
| Theoretical | Experimental § | |||||||
| Pin2[G] | FWGALAKGALKLIGSLFSSFSKKD | 0.379 | 0.429 | 0.465 | +3 | 51.5 | 2572.0 | 2572.4 |
| Pin2[14] | FWGLKGLKKFSKKL | −0.357 | 0.427 | 0.417 | +5 | 30.5 | 1680.1 | 1680.3 |
| P18K | GILKTIKSIASKLKRKAK | −0.328 | 0.455 | 0.573 | +7 | 33.5 | 1983.5 | 1984.0 |
| FA1 | GILKTIKSIASKVANTVQKLKRKAKNAV | −0.171 | 0.442 | 0.535 | +8 | 42.0 | 3008.6 | 3009.5 |
| Antimicrobials | Minimal Inhibitory Concentrations (µg/mL) a | |||
|---|---|---|---|---|
| E. coli | P. aureginosa | S. aureus | K. pneumoniae | |
| Peptides | ||||
| Pin2[14] | >50 | >50 | >50 | >50 |
| P18K | >50 | >50 | >50 | >50 |
| Pin2[G] | 50 | >50 | 25 | >50 |
| FA1 | 6.25 | 25 | >50 | >50 |
| Antibiotics | ||||
| Amoxicillin | 12.5 | >50 | 1.5 | >50 |
| Gentamicin | 1.5 | 1.5 | 1.5 | 12.5 |
| Vancomycin | >50 | >50 | 1.5 | >50 |
| Ceftriaxone | 1.5 | 12.5 | 1.5 | >50 |
| Sulfamethoxazole | >50 | >50 | >50 | >50 |
| Levofloxacin | 1.5 | 1.5 | 1.5 | 50 |
| Azitromicin | 1.5 | 12.5 | 3.1 | >50 |
| Trimethoprim | 1.5 | >50 | 1.5 | >50 |
| Peptide-Antibiotic | Best Concentration of Mixtures a (MIC, µg/mL) | Peptide b (MIC, µg/mL) | Antibiotic c (MIC, µg/mL) | FIC Index d | |
|---|---|---|---|---|---|
| Peptide | Antibiotic | ||||
| FA1-Amoxicillin | >50 | >50 | >50 | >50 | 2> |
| FA1-Azithromycin | 25 | 1.2 | >50 | >50 | <0.524 |
| FA1-Ceftriaxone | 25 | 1.2 | >50 | >50 | <0.524 |
| FA1-Gentamicin | 50 | 3.1 | >50 | 12.5 | <1.25 |
| FA1-Levofloxacin | 12.5 | 12.5 | >50 | 50 | <0.48 |
| FA1-Sulfamethoxazole | 6.25 | 1.2 | >50 | >50 | <0.15 |
| FA1-Trimethoprim | 50 | 3.1 | >50 | >50 | <1.062 |
| FA1-Vancomycin | 12.5 | 25 | >50 | >50 | <0.75 |
| Pin2[G]-Amoxicillin | >50 | >50 | >50 | >50 | 2> |
| Pin2[G]-Azithromycin | 25 | 25 | >50 | >50 | <1 |
| Pin2[G]-Ceftriaxone | 50 | 12.5 | >50 | >50 | <1.25 |
| Pin2[G]-Gentamicin | 6.25 | 12.5 | >50 | 12.5 | <1.25 |
| Pin2[G]-Levofloxacin | 25 | 25 | >50 | 50 | <1 |
| Pin2[G]-Sulfamethoxazole | 50 | 1.2 | >50 | >50 | <1.024 |
| Pin2[G]-Trimethoprim | >50 | >50 | >50 | >50 | 2> |
| Pin2[G]-Vancomycin | 50 | 50 | >50 | >50 | <2 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arenas, I.; Villegas, E.; Walls, O.; Barrios, H.; Rodríguez, R.; Corzo, G. Antimicrobial Activity and Stability of Short and Long Based Arachnid Synthetic Peptides in the Presence of Commercial Antibiotics. Molecules 2016, 21, 225. https://doi.org/10.3390/molecules21020225
Arenas I, Villegas E, Walls O, Barrios H, Rodríguez R, Corzo G. Antimicrobial Activity and Stability of Short and Long Based Arachnid Synthetic Peptides in the Presence of Commercial Antibiotics. Molecules. 2016; 21(2):225. https://doi.org/10.3390/molecules21020225
Chicago/Turabian StyleArenas, Ivan, Elba Villegas, Oliver Walls, Humberto Barrios, Ramon Rodríguez, and Gerardo Corzo. 2016. "Antimicrobial Activity and Stability of Short and Long Based Arachnid Synthetic Peptides in the Presence of Commercial Antibiotics" Molecules 21, no. 2: 225. https://doi.org/10.3390/molecules21020225