
Solanum nigrum Protects against Hepatic Fibrosis via Suppression of Hyperglycemia in High-Fat/Ethanol Diet-Induced Rats

 ,
,
Abstract
:1. Introduction
2. Results
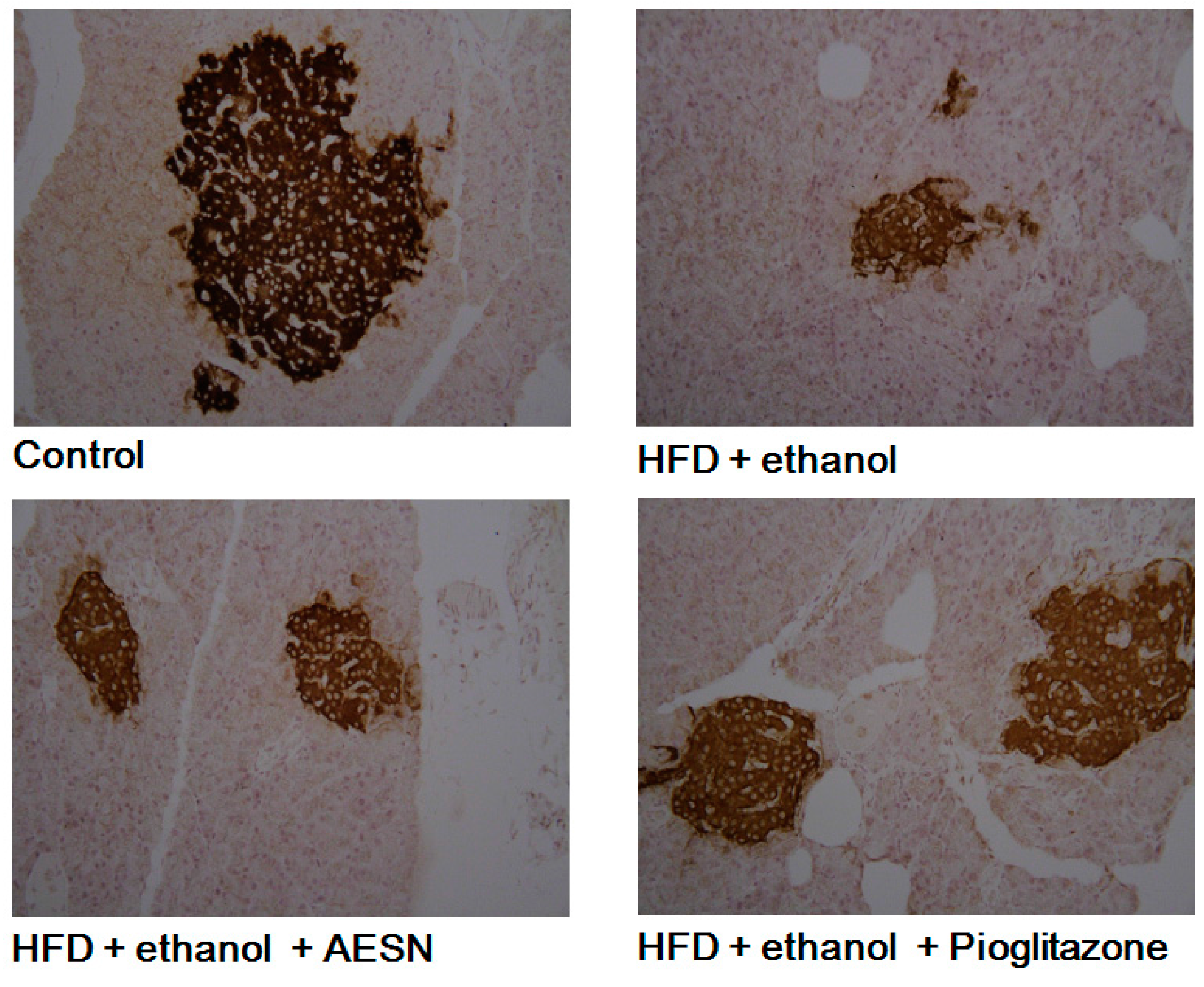
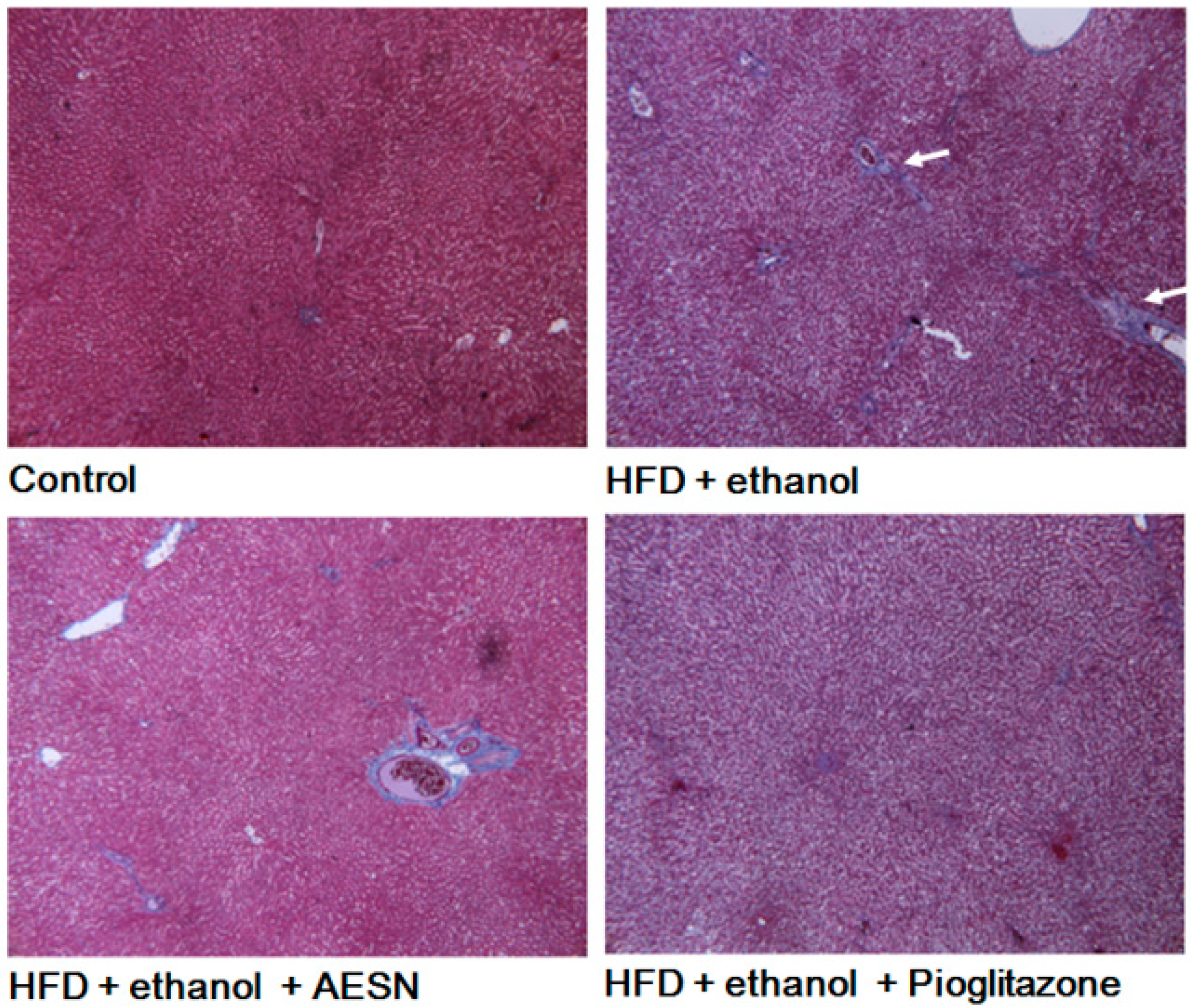
2.1. AESN Inhibits Hepatic Fibrosis
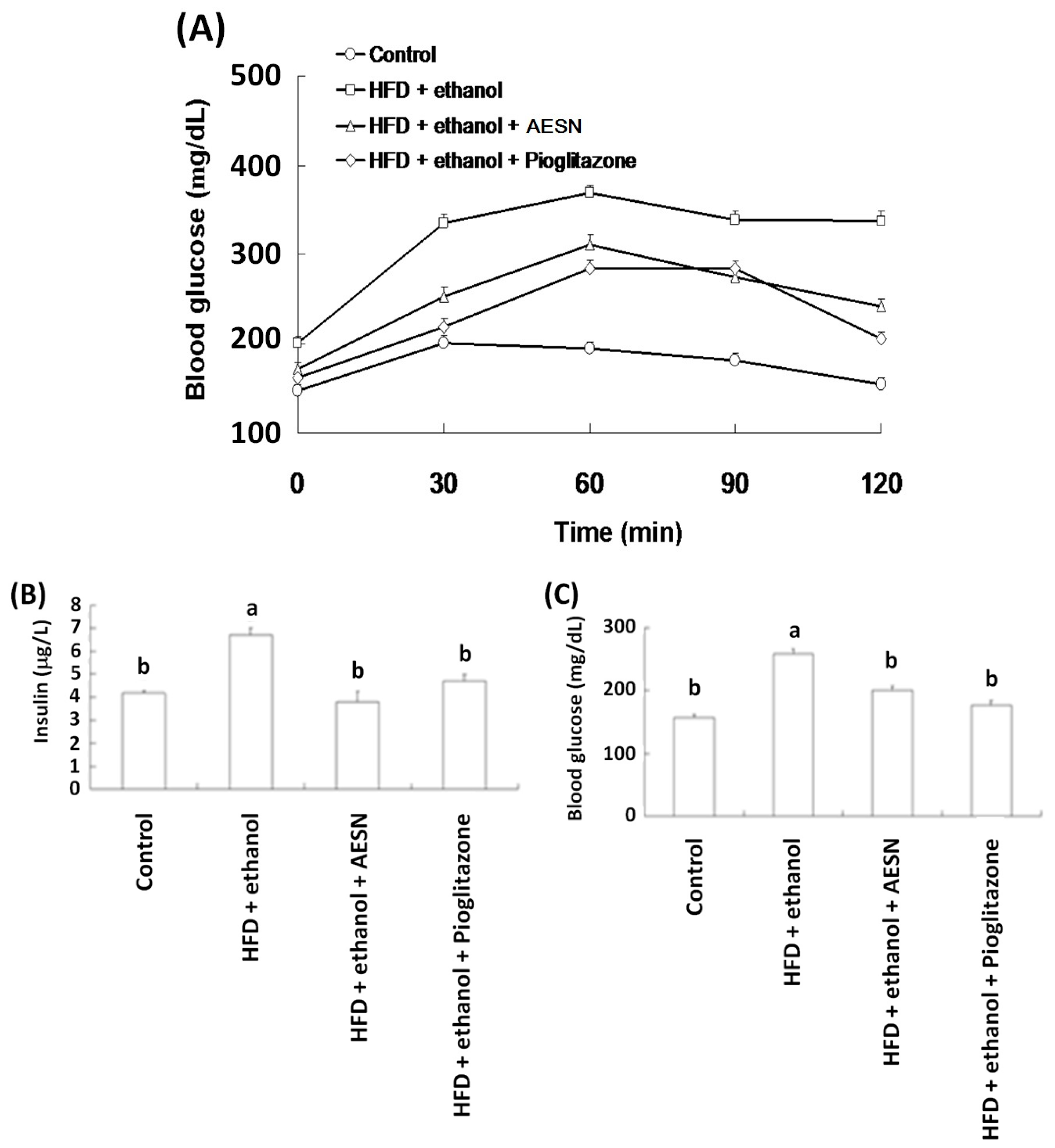
2.2. Anti-Diabetic Activity of AESN
2.3. AESN Protects against Liver Damage Induced by HFD/Ethanol Treatment
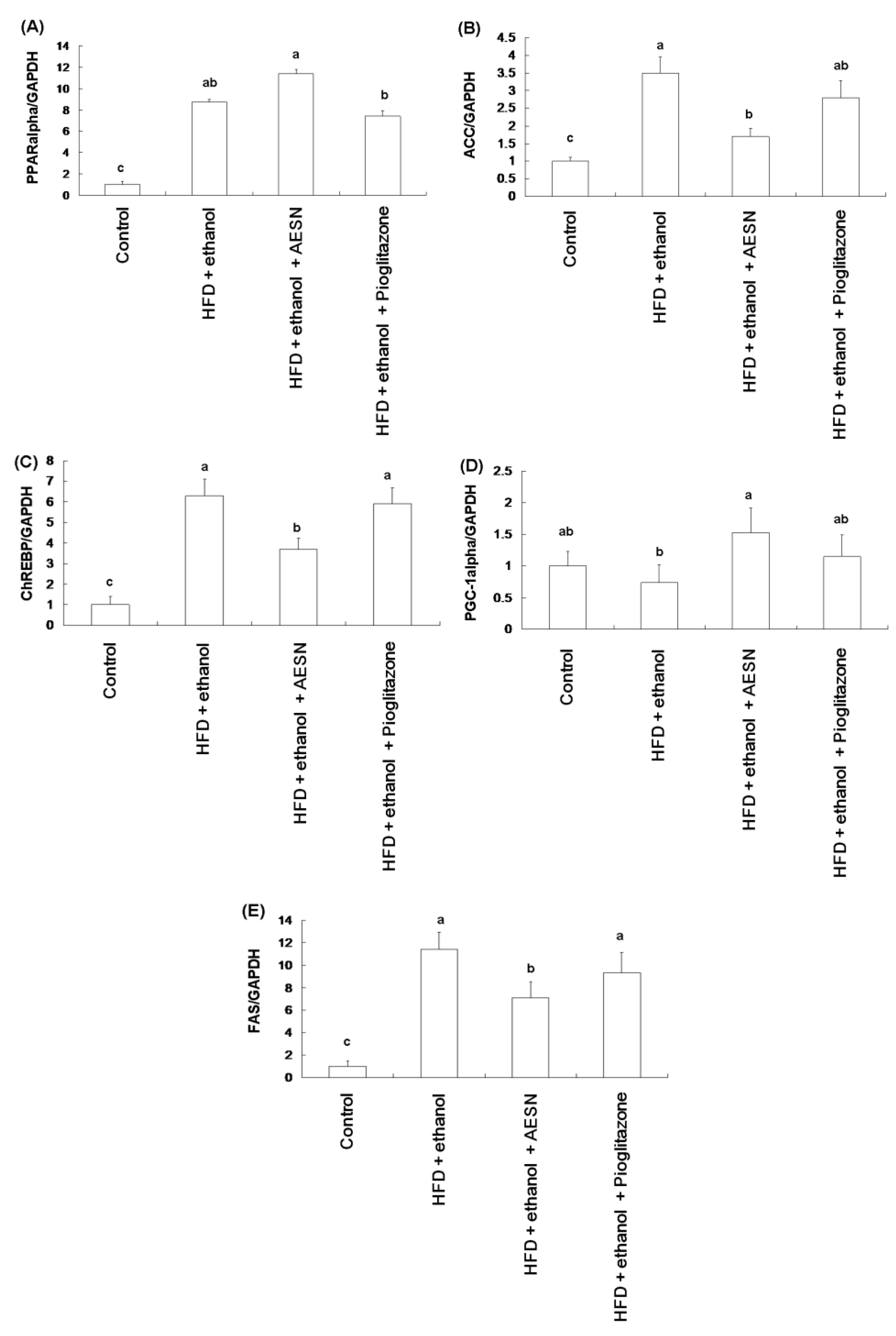
2.4. AESN Improves Fatty Liver
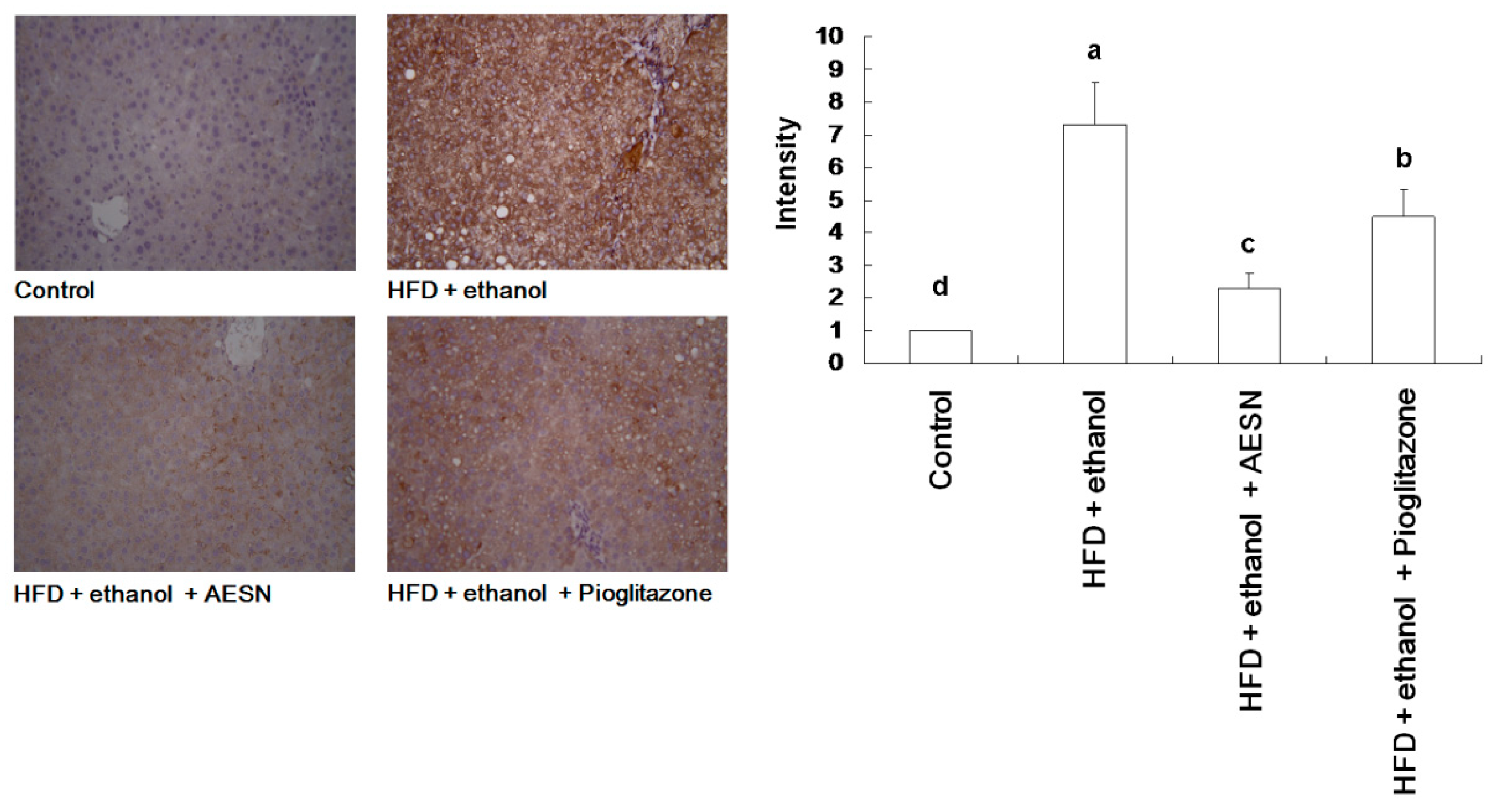
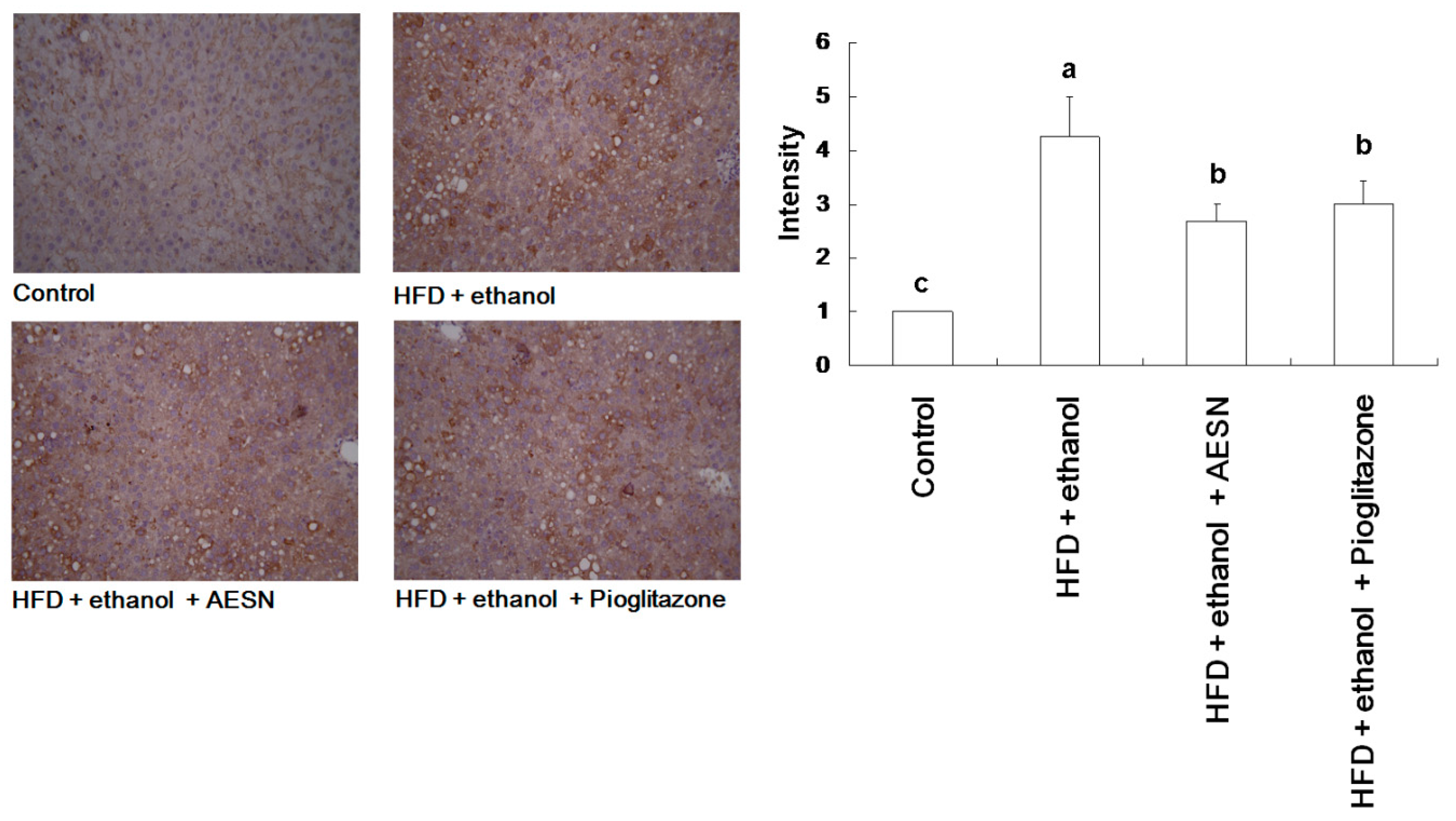
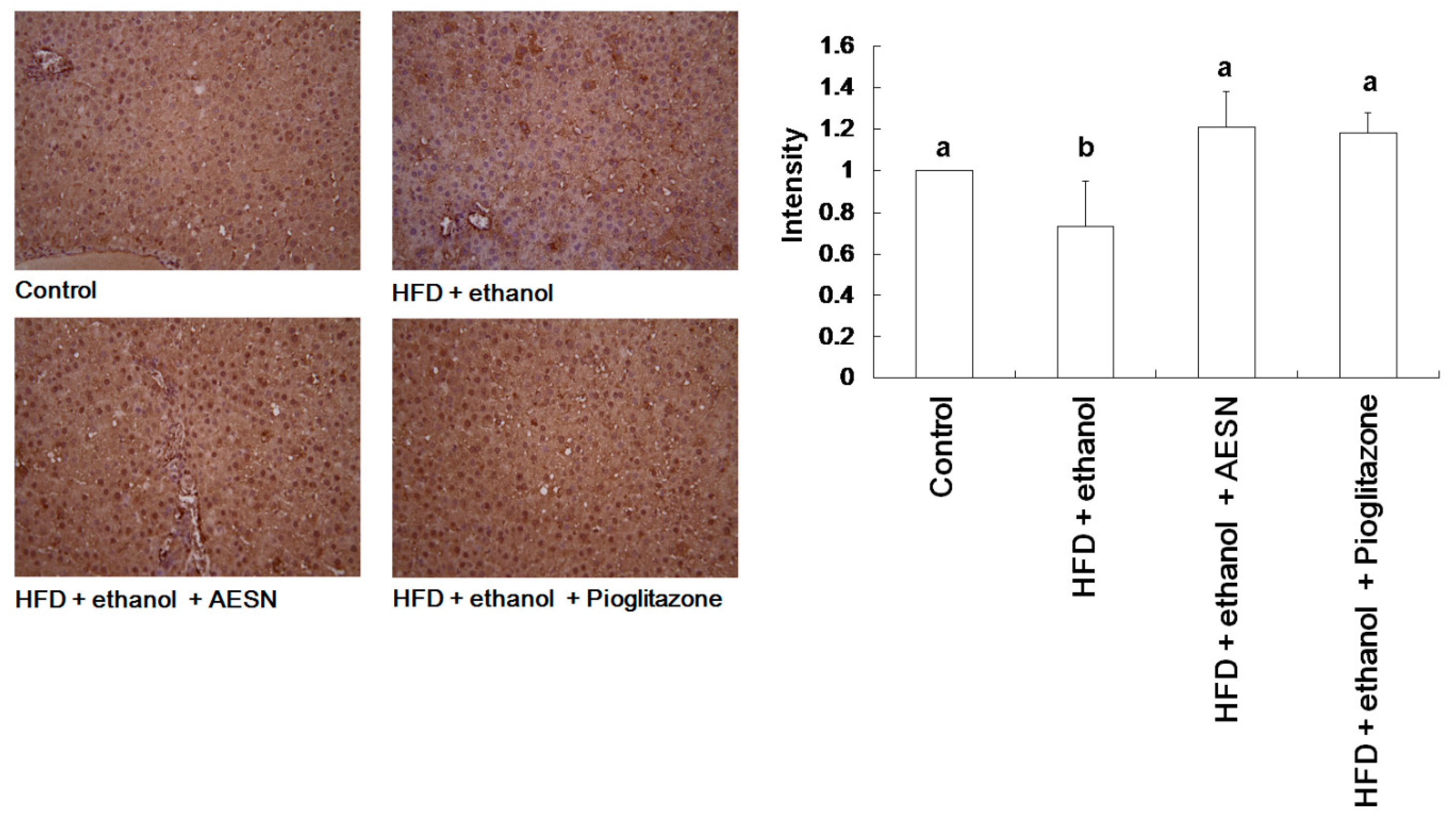
2.5. AESN Inhibits RAGE in HFD/Ethanol Treated-Rats
3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. Animal Treatments
4.3. Oral Glucose Tolerance Test (OGTT)
4.4. Assays for Serum Insulin, Aspartate Transaminase (AST), and Alanine Transaminase (ALT)
4.5. RNA Preparation and Real-Time PCR
4.6. Hepatic Fibrosis Stain
4.7. Immunohistochemistry (IHC) Stain
4.8. HSCs-t6 Cell Culture
4.9. Western Blot
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACC | acetyl-CoA carboxylase |
| AESN | aqueous extracts from Solanum nigrum |
| AGEs | Advanced glycation end products |
| ChREBP | carbohydrate response element-binding protein |
| FAS | fatty acid synthase |
| FBS | fetal bovine serum |
| FFA | free fatty acids |
| HASH | nonalcoholic steatohepatitis |
| HDL-C | high density lipoprotein cholesterol |
| HFD | high-fat diet |
| HSCs | hepatic stellate cells |
| ITT | insulin tolerance test |
| Nrf2 | nuclear factor-erythroid 2-related factor 2 |
| OGTT | oral glucose tolerance test |
| PGC-1α | PPARγ co-activator (PGC-1) |
| PPARα | peroxisome proliferator-activated receptor α |
| RAGE | Receptor for AGEs |
| TC | cholesterol |
| TG | triglycerides |
References
- Salt, W.B. Nonalcoholic fatty liver disease (NAFLD): A comprehensive review. J. Insur. Med. 2004, 36, 27–41. [Google Scholar] [PubMed]
- Lalor, P.F.; Faint, J.; Aarbodem, Y.; Hubscher, S.G.; Adams, D.H. The role of cytokines and chemokines in the development of steatohepatitis. Semin. Liver Dis. 2007, 27, 173–193. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Chen, T.H.; Lee, B.H.; Hsu, Y.W.; Pan, T.M. Monascin and ankaflavin act as natural AMPK activators with PPARalpha agonist activity to down-regulate nonalcoholic steatohepatitis in high-fat diet-fed C57BL/6 mice. Food Chem. Toxicol. 2014, 64, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Lee, B.H.; Chang, Y.Y.; Hsu, Y.W.; Pan, T.M. A novel natural Nrf2 activator with PPARgamma-agonist (monascin) attenuates the toxicity of methylglyoxal and hyperglycemia. Appl. Toxicol. Pharmacol. 2013, 272, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Lee, B.H.; Hsu, Y.W.; Pan, T.M. Peroxisome proliferator-activated receptor-gamma activators monascin and rosiglitazone attenuate carboxymethyllysine-induced fibrosis in hepatic stellate cells through regulating the oxidative stress pathway but independent of the receptor for advanced glycation end products signaling. J. Agric. Food Chem. 2013, 61, 6873–6879. [Google Scholar] [PubMed]
- Bacon, B.R.; Farahvash, M.J.; Janney, C.G.; Neuschwander-Tetri, B.A. Nonalcoholic steatohepatitis: An expanded clinical entity. Gastroenterology 1994, 107, 1103–1109. [Google Scholar] [PubMed]
- Ludwig, J.; Viggiano, T.R.; McGill, D.B.; Oh, B.J. Nonalcoholic steatohepatitis: Mayo Clinic experiences with a hitherto unnamed disease. Mayo Clin. Proc. 1980, 55, 434–438. [Google Scholar] [PubMed]
- Dam-Larsen, S.; Franzmann, M.; Andersen, I.B.; Christoffersen, P.; Jensen, L.B.; Sorensen, T.I.; Becker, U.; Bendtsen, F. Long term prognosis of fatty liver: Risk of chronic liver disease and death. Gut 2004, 53, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Zhao, Y.; Tang, Y. Chronic alcohol exposure stimulates adipose tissue lipolysis in mice: Role of reverse triglyceride transport in the pathogenesis of alcoholic steatosis. Am. J. Pathol. 2012, 180, 998–1007. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, M.; Kojima, T.; Ohbora, A. Protective effect of alcohol consumption for fatty liver but not metabolic syndrome. World J. Gastroenterol. 2012, 18, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Hsu, W.H.; Hsu, Y.W.; Pan, T.M. Suppression of dimerumic acid on hepatic fibrosis caused from carboxymethyl-lysine (CML) by attenuating oxidative stress depends on Nrf2 activation in hepatic stellate cells (HSCs). Food Chem. Toxicol. 2013, 62, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.; Lin, Y.F.; Tai, C.J.; Wang, C.W.; Chang, Y.J.; Choong, C.Y.; Lin, C.S.; Tai, C.J.; Chang, C.C. Integrated treatment of aqueous extract of Solanum nigrum-potentiated cisplatin- and doxorubicin-induced cytotoxicity in human hepatocellular carcinoma cells. Evid. Based Complement. Altern. Med. 2015, 2015, 675270. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.W.; Chen, C.L.; Wang, C.K.; Chang, Y.J.; Jian, J.Y.; Lin, C.S.; Tai, C.J.; Tai, C.J. Cisplatin-, doxorubicin- and docetaxel-induced cell death promoted by the aqueous extract of Solanum nigrum in human ovarian carcinoma cells. Integr. Cancer Ther. 2015, 14, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.J.; Wang, C.K.; Chang, Y.J.; Lin, C.S.; Tai, C.J. Aqueous extract of Solanum nigrum leaf activates autophagic cell death and enhances docetaxel-induced cytotoxicity in human endometrial carcinoma cells. Evid. Based Complement. Altern. Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.J.; Wang, C.K.; Tai, C.J.; Lin, Y.F.; Lin, C.S.; Jian, J.Y.; Chang, Y.J.; Chang, C.C. Aqueous extract of Solanum nigrum leaves induces autophage and enhances cytotoxicity of cisplatin, doxorubicin, docetaxel, and 5-fluorouracil in human colorectal carcinoma cells. Evid. Based Complement. Altern. Med. 2013, 2013, 514719. [Google Scholar]
- Lin, H.M.; Tseng, H.C.; Wang, C.J.; Lin, J.J.; Lo, C.W.; Chou, F.P. Hepatoprotective effects of Solanum nigrum Linn extract against CCl(4)-induced oxidative damage in rats. Chem. Biol. Interact. 2008, 171, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Fang, H.L.; Lina, W.C. Inhibitory effect of Solanum nigrum on thioacetamide-induced liver fibrosis in mice. J. Ethnopharmacol. 2008, 119, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Hsu, W.H.; Huang, T.; Chang, Y.Y.; Hsu, Y.W.; Pan, T.M. Monascin improves diabetes and dyslipidemia by regulating PPARgamma and inhibiting lipogenesis in fructose-rich diet-induced C57BL/6 mice. Food Funct. 2013, 4, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.I.; Dong, C.; Gardener, H. Serum levels of soluble receptor for advanced glycation end-products and metabolic syndrome: The Northern Manhattan study. Metabolism 2014, 63, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Max, M.; Thamm, K. RAGE influences obesity in mice. Effects of the presence of RAGE on weight gain, AGE accumulation, and insulin levels in mice on a high fat diet. Z. Gerontol. Geriatr. 2012, 45, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Hsu, W.H.; Chang, Y.Y.; Kuo, H.F.; Hsu, Y.W.; Pan, T.M. Ankaflavin: A natural novel PPARγ agonist upregulates Nrf2 to attenuate methylglyoxal-induced diabetes in vivo. Free Radic. Biol. Med. 2012, 53, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Vander-Jagt, D.; Hunsaker, L. Methylglyoxal metabolism and diabetic complications: Roles of aldose reductase, glyxalase-I, betaine aldehyde dehydrogenase and oxoaldehyde dehydrogenase. Chem. Biol. Int. 2003, 143, 341–351. [Google Scholar] [CrossRef]
- Weng, C.J.; Chen, M.J.; Yeh, C.T.; Yen, G.C. Hepatoprotection of quercetin against oxidative stress by induction of metallothionein expression through activating MAPK and PI3K pathways and enhancing Nrf2 DNA-binding activity. New Biotechnol. 2011, 28, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Sohrabipour, S.; Kharazmi, F.; Soltani, N.; Kamalinejad, M. Effect of the administration of Solanum nigrum fruit on blood glucose, lipid profiles, and sensitivity of the vascular mesenteric bed to phenylephrine in streptozotocin-induced diabetic rats. Med. Sci. Monit. Basic Res. 2013, 19, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Sohrabipour, S.; Kharazmi, F.; Soltani, N.; Kamalinejad, M. Biphasic effect of Solanum nigrum fruit aqueous extract on vascular mesenteric beds in non-diabetic and streptozotocin-induced diabetic rats. Pharmacogn. Res. 2014, 6, 148–152. [Google Scholar]
- Uyeda, K.; Repa, J.J. Carbohydrate response element binding protein, ChREBP, a transcription factor coupling hepatic glucose utilization and lipid synthesis. Cell Metab. 2006, 4, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, K.; Horikawa, Y. ChREBP: A glucose-activated transcription factor involved in the development of metabolic syndrome. Endocr. J. 2008, 55, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Repa, J.J.; Liang, G.; Ou, J. Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRalpha and LXRbeta. Genes Dev. 2000, 14, 2819–2830. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Yang, R.; Tarr, P.T. Hyperlipidemic effects of dietary saturated fats mediated through PGC-1beta coactivation of SREBP. Cell 2005, 120, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.H.; Liao, A.C.; Hung, J.H.; Lee, W.J.; Hu, K.C.; Lin, P.T.; Liao, R.F.; Chen, P.S. Alpha-Solanine inhibits invasion of human prostate cancer cells by suppressing epithelial-mesenchymal transition and MMPs expression. Molecules 2014, 19, 11896–11914. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Jeong, H.D.; Choi, H.Y. The chloroform fraction of Solanum nigrum suppresses nitric oxide and tumor necrosis factor-alpha in LPS-stimulated mouse peritoneal macrophages through inhibition of p38, JNK and ERK1/2. Am. J. Chin. Med. 2011, 39, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Analysis of biologically active compounds in potatoes (Solanum tuberosum), tomatoes (Lycopersicon esculentum), and jimson weed (Datura stramonium) seeds. J. Chromatogr. A 2004, 1054, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chang, T. Methylglyoxal content in drinking coffee as a cytotoxic factor. J. Food Sci. 2010, 75, H167–H171. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.J.; Manley-Harris, M.; Molan, P.C. The origin of methylglyoxal in New Zealand manuka (Leptospermum scoparium) honey. Carbohydr. Res. 2009, 344, 1050–1053. [Google Scholar] [CrossRef] [PubMed]
- Arrbias-Lorenzo, G.; Morales, F.J. Analysis, distribution, and dietary exposure of glyoxal and methylglyoxal in cookies and their relationship with other heat-induced contaminants. J. Agric. Food Chem. 2010, 58, 2966–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sample Availability: Not available.





| Mean Score | SD | |
|---|---|---|
| Control | 0 | 0 |
| HFD + Ethanol | 3.26 | 0.45 |
| HFD + Ethanol + AESN | 2.73 | 0.38 |
| HFD + Ethanol + Pioglitazone | 2.15 | 0.52 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | TC (mg/dL) | TG (mg/dL) | HDL-C (mg/dL) | FFA (mg/dL) |
|---|---|---|---|---|
| Control | 83.5 ± 13.0 c | 38.7 ± 7.8 d | 27.6 ± 3.7 a | 0.76 ± 0.04 c |
| HFD + Ethanol | 187.6 ± 17.2 a | 89.1 ± 5.7 a | 14.5 ± 1.3 b | 4.63 ± 0.16 a |
| HFD + Ethanol + AESN | 137.3 ± 14.1 b | 57.3 ± 7.8 c | 13.3 ± 2.2 b | 2.45 ± 0.15 b |
| HFD + Ethanol + Pioglitazone | 164.2 ± 15.2 a,b | 69.0 ± 8.4 b | 15.7 ± 3.2 b | 3.41 ± 0.11 a |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tai, C.-J.; Choong, C.-Y.; Shi, Y.-C.; Lin, Y.-C.; Wang, C.-W.; Lee, B.-H.; Tai, C.-J. Solanum nigrum Protects against Hepatic Fibrosis via Suppression of Hyperglycemia in High-Fat/Ethanol Diet-Induced Rats. Molecules 2016, 21, 269. https://doi.org/10.3390/molecules21030269
Tai C-J, Choong C-Y, Shi Y-C, Lin Y-C, Wang C-W, Lee B-H, Tai C-J. Solanum nigrum Protects against Hepatic Fibrosis via Suppression of Hyperglycemia in High-Fat/Ethanol Diet-Induced Rats. Molecules. 2016; 21(3):269. https://doi.org/10.3390/molecules21030269
Chicago/Turabian StyleTai, Cheng-Jeng, Chen-Yen Choong, Yeu-Ching Shi, Yu-Chun Lin, Chia-Woei Wang, Bao-Hong Lee, and Chen-Jei Tai. 2016. "Solanum nigrum Protects against Hepatic Fibrosis via Suppression of Hyperglycemia in High-Fat/Ethanol Diet-Induced Rats" Molecules 21, no. 3: 269. https://doi.org/10.3390/molecules21030269