
Antibacterial Activity and Membrane-Disruptive Mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic Acid, a Novel Phenolic Compound from Pine Needles of Cedrus deodara, against Staphylococcus aureus

Abstract
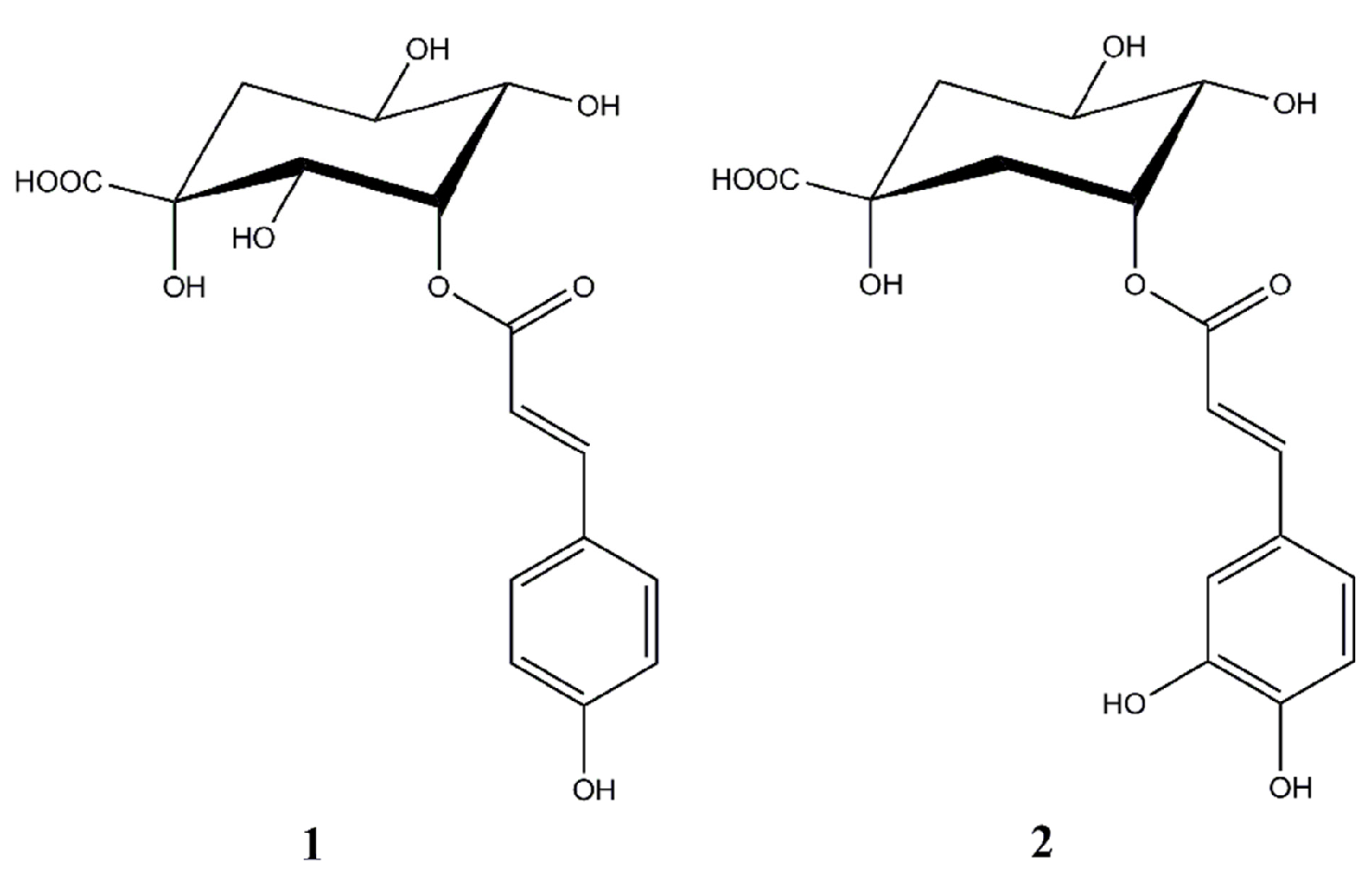
:1. Introduction
2. Results
2.1. Antibacterial Activities of CHQA
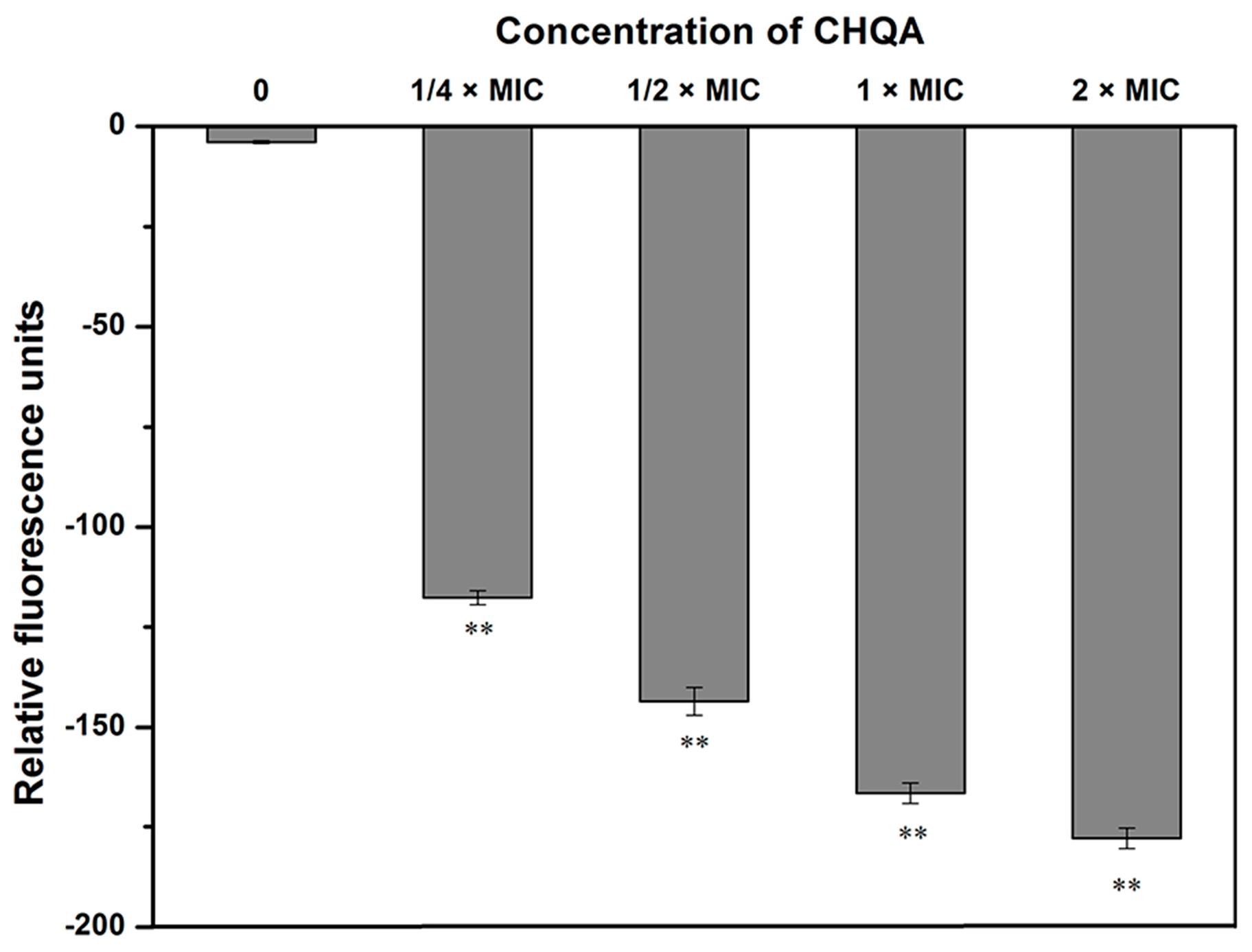
2.2. Effect of CHQA on Membrane Potential
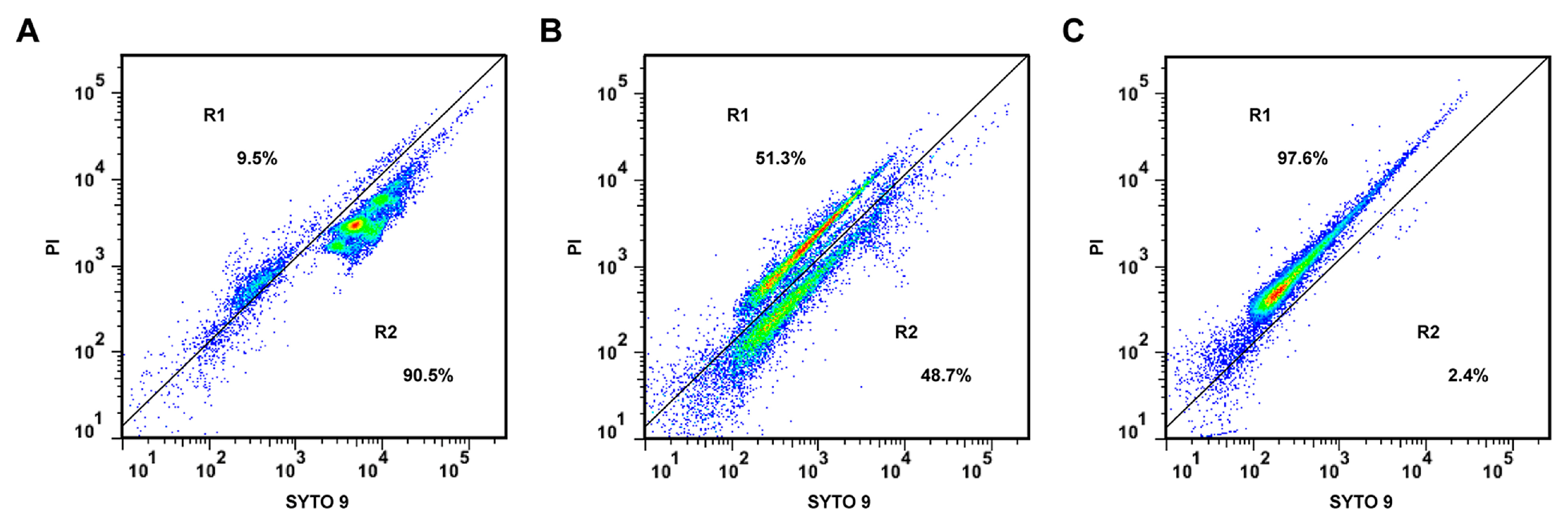
2.3. Effect of CHQA on Membrane Integrity
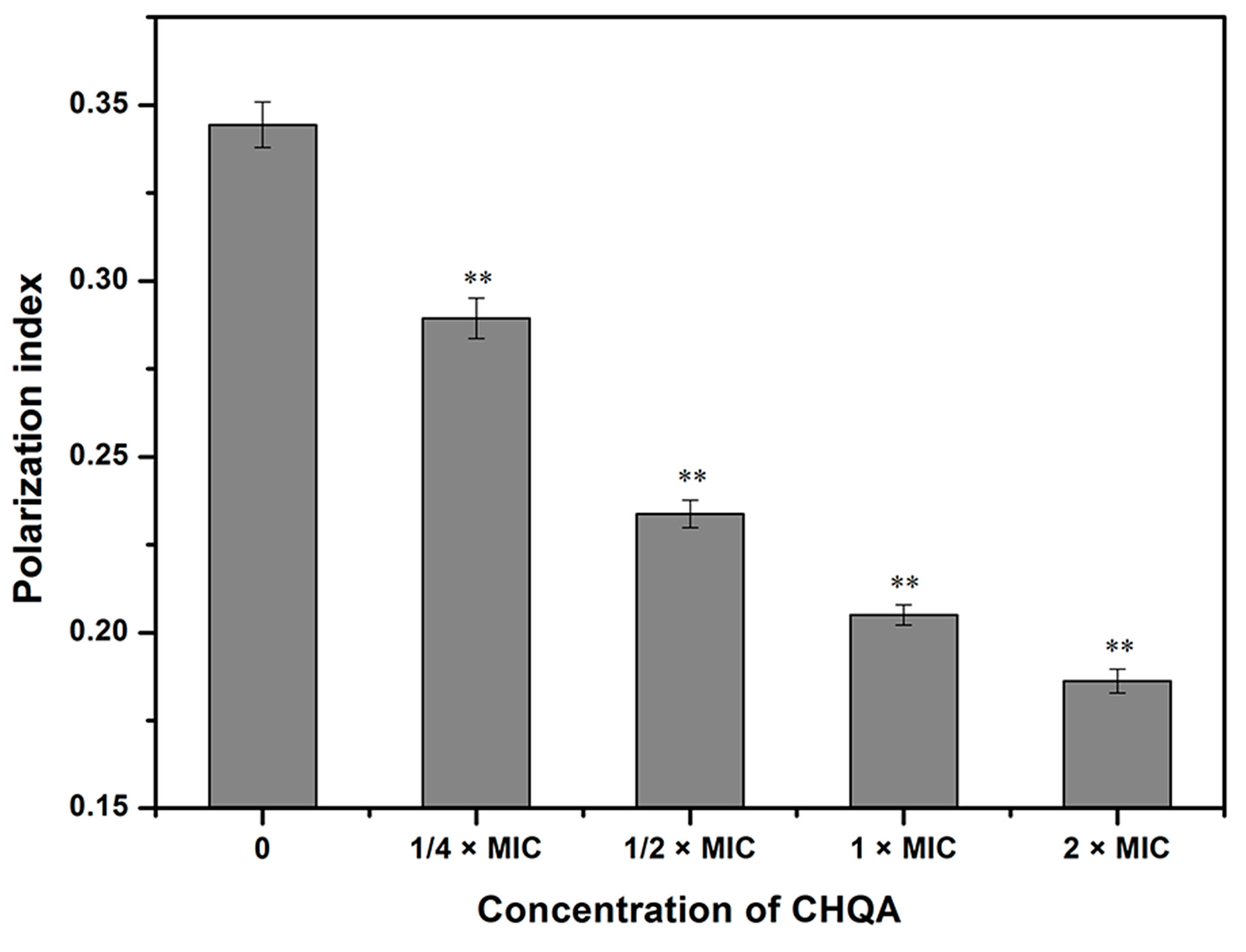
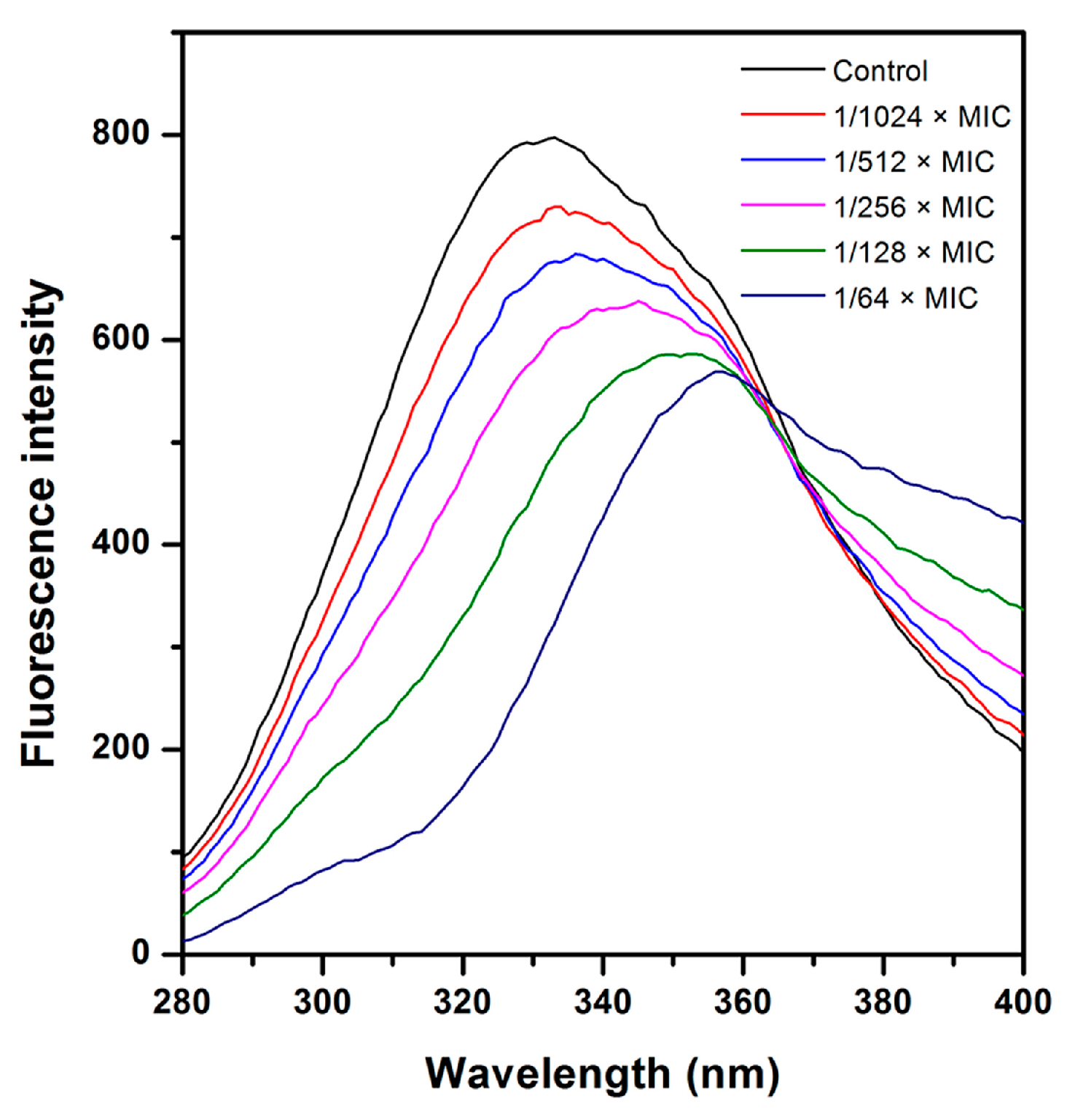
2.4. Effect of CHQA on Membrane Fluidity
2.5. Effect of CHQA on Membrane Protein
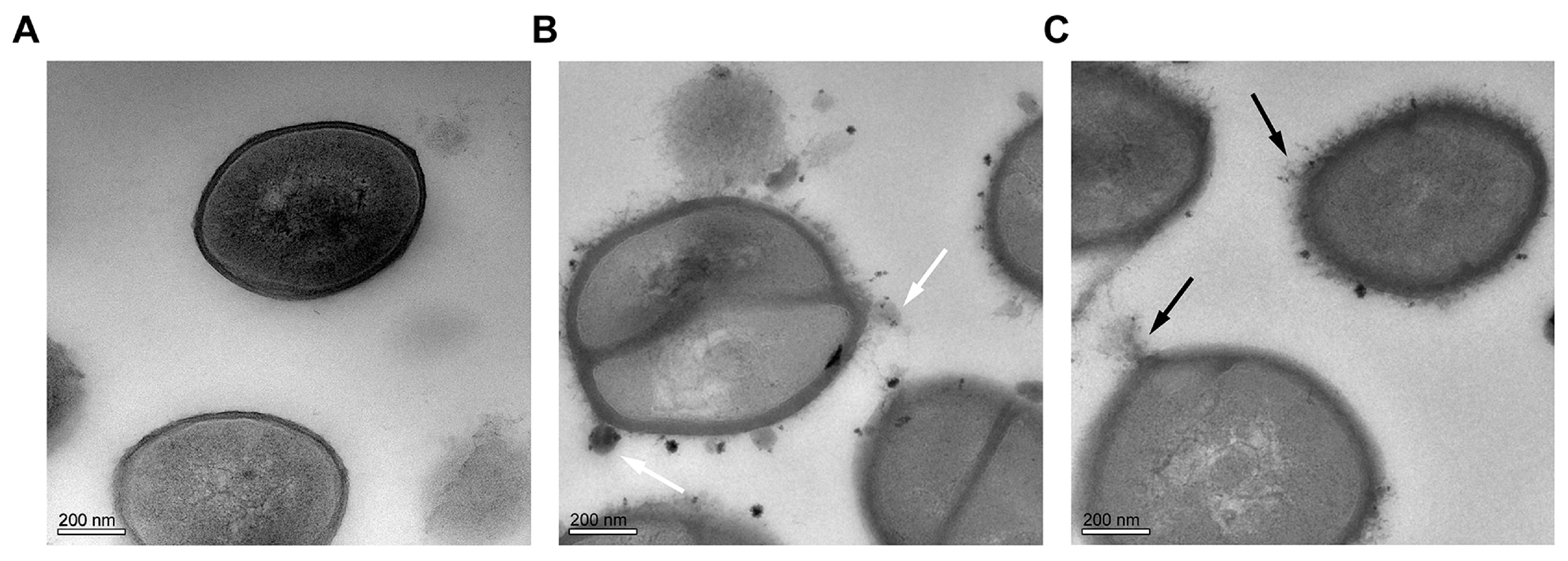
2.6. Effect of CHQA on Cell Morphology
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Bacterial Strains and Culture Conditions
4.3. Determination of the Minimum Inhibitory Concentration (MIC)
4.4. Membrane Potential
4.5. Flow Cytometric Analysis
4.6. Membrane Fluidity
4.7. Membrane Protein
4.8. Transmission Electron Microscope (TEM)
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar] [PubMed]
- Lee, S.Y.; Lim, S.I.; Ahn, D.H. Antibacterial mechanism of Myagropsis myagroides extract on Listeria monocytogenes. Food Control 2014, 42, 23–28. [Google Scholar] [CrossRef]
- Rozemeijer, W.; Fink, P.; Rojas, E.; Jones, C.H.; Pavliakova, D.; Giardina, P.; Murphy, E.; Liberator, P.; Jiang, Q.; Girgenti, D. Evaluation of approaches to monitor Staphylococcus aureus virulence factor expression during human disease. PLoS ONE 2015, 10, e0116945. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Sun, Y.; Zheng, Z.; Zhang, X.; Song, K.; Jia, Z.; Chen, Y.; Yang, M.; Liu, X.; Dong, R. Antimicrobial activity of syringic acid against Cronobacter sakazakii and its effect on cell membrane. Food Chem. 2016, 197, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Shen, S.; Xu, J.; Lin, S.; Yuan, Y.; Jones, G.S. Synergistic interactions of cinnamaldehyde in combination with carvacrol against food-borne bacteria. Food Control 2013, 34, 619–623. [Google Scholar] [CrossRef]
- Sanchez Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Plant defence mechanisms and enzymatic transformation products and their potential applications in food preservation: Advantages and limitations. Trends Food Sci. Technol. 2015, 46, 49–59. [Google Scholar] [CrossRef]
- Wu, Y.P.; Liang, X.; Liu, X.Y.; Zhong, K.; Gao, B.; Huang, Y.N.; Gao, H. Cedrus deodara pine needle as a potential source of natural antioxidants: Bioactive constituents and antioxidant activities. J. Funct. Foods 2015, 14, 605–612. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. p-Coumaric acid kills bacteria through dual damage mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Chen, J.; Yang, M.L.; Zeng, J.; Gao, K. Antimicrobial activity of Araucaria cunninghamii sweet and the chemical constituents of its twigs and leaves. Phytochem. Lett. 2013, 6, 41–45. [Google Scholar] [CrossRef]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2014, 238, 589–596. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef] [PubMed]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.U.; Egli, T. Assessment and interpretation of bacterial viability by using the live/dead baclight kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Lu, Y.H. Comparison of inhibitory activities and mechanisms of five mulberry plant bioactive components against α-glucosidase. J. Agric. Food Chem. 2013, 61, 8110–8119. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ren, C.; Zhang, Y.; Liu, X.; Yao, X.; Hu, Z. Spectroscopic studies on binding of puerarin to human serum albumin. J. Mol. Struct. 2008, 885, 64–69. [Google Scholar] [CrossRef]
- Lin, Y.T.; Labbe, R.G.; Shetty, K. Inhibition of Listeria monocytogenes in fish and meat systems by use of oregano and cranberry phytochemical synergies. Appl. Environ. Microbiol. 2004, 70, 5672–5678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.; Wang, Y.; Jiang, P.; Quek, S. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Control 2016, 59, 282–289. [Google Scholar] [CrossRef]
- Figueiredo, A.R.; Campos, F.; de Freitas, V.; Hogg, T.; Couto, J.A. Effect of phenolic aldehydes and flavonoids on growth and inactivation of Oenococcus oeni and Lactobacillus hilgardii. Food Microbiol. 2008, 25, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.G.; Wyllie, S.G.; Markham, J.L. Antimicrobially active terpenes cause K+ leakage in E. coli cells. J. Essent. Oil Res. 2005, 17, 686–690. [Google Scholar] [CrossRef]
- Stojković, D.; Petrović, J.; Soković, M.; Glamočlija, J.; Kukić-Marković, J.; Petrović, S. In situ antioxidant and antimicrobial activities of naturally occurring caffeic acid, p-coumaric acid and rutin, using food systems. J. Sci. Food Agric. 2013, 93, 3205–3208. [Google Scholar] [CrossRef] [PubMed]
- Char, C.D.; Guerrero, S.N.; Alzamora, S.M. Mild thermal process combined with vanillin plus citral to help shorten the inactivation time for Listeria innocua in orange juice. Food Bioprocess Technol. 2010, 3, 752–761. [Google Scholar] [CrossRef]
- Whiteaker, K.L.; Gopalakrishnan, S.M.; Groebe, D.; Shieh, C.C.; Warrior, U.; Burns, D.J.; Coghlan, M.J.; Scott, V.E.; Gopalakrishnani, M. Validation of FLIPR membrane potential dye for high throughput screening of potassium channel modulators. J. Biomol. Screen. 2001, 6, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Bot, C.; Prodan, C. Probing the membrane potential of living cells by dielectric spectroscopy. Eur. Biophys. J. 2009, 38, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.D.; Mann, C.M.; Markham, J.L.; Gustafson, J.E.; Warmington, J.R.; Wyllie, S.G. Determining the antimicrobial actions of tea tree oil. Molecules 2001, 6, 87–91. [Google Scholar] [CrossRef]
- Sánchez, E.; García, S.; Heredia, N. Extracts of edible and medicinal plants damage membranes of Vibrio cholerae. Appl. Environ. Microbiol. 2010, 76, 6888–6894. [Google Scholar] [CrossRef] [PubMed]
- Mykytczuk, N.C.S.; Trevors, J.T.; Leduc, L.G.; Ferroni, G.D. Fluorescence polarization in studies of bacterial cytoplasmic membrane fluidity under environmental stress. Prog. Biophys. Mol. Biol. 2007, 95, 60–82. [Google Scholar] [CrossRef] [PubMed]
- Cartron, M.L.; England, S.R.; Chiriac, A.I.; Josten, M.; Turner, R.; Rauter, Y.; Hurd, A.; Sahl, H.G.; Jones, S.; Foster, S.J. Bactericidal activity of the human skin fatty acid cis-6-hexadecanoic acid on Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 3599–3609. [Google Scholar] [CrossRef] [PubMed]
- Cherrat, L.; Dumas, E.; Bakkali, M.; Degraeve, P.; Laglaoui, A.; Oulahal, N. Effect of essential oils on cell viability, membrane integrity and membrane fluidity of Listeria innocua and Escherichia coli. J. Essent. Oil Bear. Plants 2016, 19, 155–166. [Google Scholar] [CrossRef]
- Nowotarska, S.W.; Nowotarski, K.J.; Friedman, M.; Situ, C. Effect of structure on the interactions between five natural antimicrobial compounds and phospholipids of bacterial cell membrane on model monolayers. Molecules 2014, 19, 7497–7515. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Li, X.; Yuan, L.; Ge, L.; Zhang, B.; Zhou, S. Interaction of houttuyfonate homologues with the cell membrane of gram-positive and gram-negative bacteria. Colloid Surf. A Physicochem. Eng. Asp. 2007, 301, 412–418. [Google Scholar] [CrossRef]
- Ye, X.; Xu, L.; Li, X.; Chen, Z.; Zhang, B.; Yuan, L.; Chen, X.; Zhang, H.; Chang, W.; Sun, S. Antibacterial mechanism of houttuyfonate homologues against Bacillus subtilis. Colloid Surf. A Physicochem. Eng. Asp. 2009, 350, 130–135. [Google Scholar] [CrossRef]
- Bai, J.R.; Wu, Y.P.; Liu, X.Y.; Zhong, K.; Huang, Y.N.; Gao, H. Antibacterial activity of shikimic acid from pine needles of Cedrus deodara against Staphylococcus aureus through damage to cell membrane. Int. J. Mol. Sci. 2015, 16, 27145–27155. [Google Scholar] [CrossRef] [PubMed]
- Teng, D.; Wang, X.; Xi, D.; Mao, R.; Zhang, Y.; Guan, Q.; Zhang, J.; Wang, J. A dual mechanism involved in membrane and nucleic acid disruption of avbd103b, a new avian defensin from the king penguin, against Salmonella enteritidis cvcc3377. Appl. Microbiol. Biotechnol. 2014, 98, 8313–8325. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.L.; Wu, J.; Chen, G.Q.; Cui, F.Z.; Kim, T.N.; Kim, J.O. A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mater. Res. 2000, 52, 662–668. [Google Scholar] [CrossRef]
- Sun, Y.; Dong, W.; Sun, L.; Ma, L.; Shang, D. Insights into the membrane interaction mechanism and antibacterial properties of chensinin-1b. Biomaterials 2015, 37, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.C.; Cunningham, T.M.; Hansen, M.R.; Haydel, S.E. Effects of antibacterial mineral leachates on the cellular ultrastructure, morphology, and membrane integrity of Escherichia coli and methicillin-resistant Staphylococcus aureus. Ann. Clin. Microbiol. Antimicrob. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.C.; He, Q.; Sun, Q.; Zhong, K.; Gao, H. Antibacterial activity of water-soluble extract from pine needles of Cedrus deodara. Int. J. Food Microbiol. 2012, 153, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compound 3-p-trans-coumaroyl-2-hydroxyquinic acid are available from the authors.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | MIC (mg/mL) | |
|---|---|---|
| 1 | 2 | |
| Gram-negative bacteria | ||
| Escherichia coli ATCC 11229 | 10 | 10 |
| Salmonella enterica ATCC 6539 | 5 | 5 |
| Vibrio parahaemolyticus ATCC 17802 | 5 | 5 |
| Gram-positive bacteria | ||
| Bacillus cereus ATCC 14579 | 2.5 | 2.5 |
| Clostridium perfringens ATCC 13124 | 5 | 5 |
| Staphylococcus aureus ATCC 6538 | 5 | 5 |
| Staphylococcus aureus ATCC 25923 | 5 | 5 |
| Staphylococcus aureus ATCC 29213 | 5 | 5 |
| Staphylococcus aureus ATCC 27217 | 5 | 5 |
| Staphylococcus aureus ATCC 29247 | 5 | 5 |
| Staphylococcus aureus ATCC 9144 | 5 | 5 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Qi, H.; Jiang, Y.; Gao, H. Antibacterial Activity and Membrane-Disruptive Mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic Acid, a Novel Phenolic Compound from Pine Needles of Cedrus deodara, against Staphylococcus aureus. Molecules 2016, 21, 1084. https://doi.org/10.3390/molecules21081084
Wu Y, Bai J, Zhong K, Huang Y, Qi H, Jiang Y, Gao H. Antibacterial Activity and Membrane-Disruptive Mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic Acid, a Novel Phenolic Compound from Pine Needles of Cedrus deodara, against Staphylococcus aureus. Molecules. 2016; 21(8):1084. https://doi.org/10.3390/molecules21081084
Chicago/Turabian StyleWu, Yanping, Jinrong Bai, Kai Zhong, Yina Huang, Huayi Qi, Yan Jiang, and Hong Gao. 2016. "Antibacterial Activity and Membrane-Disruptive Mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic Acid, a Novel Phenolic Compound from Pine Needles of Cedrus deodara, against Staphylococcus aureus" Molecules 21, no. 8: 1084. https://doi.org/10.3390/molecules21081084