
Cyanidin-3-O-glucoside: Physical-Chemistry, Foodomics and Health Effects

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Physical Chemistry of Cy3G
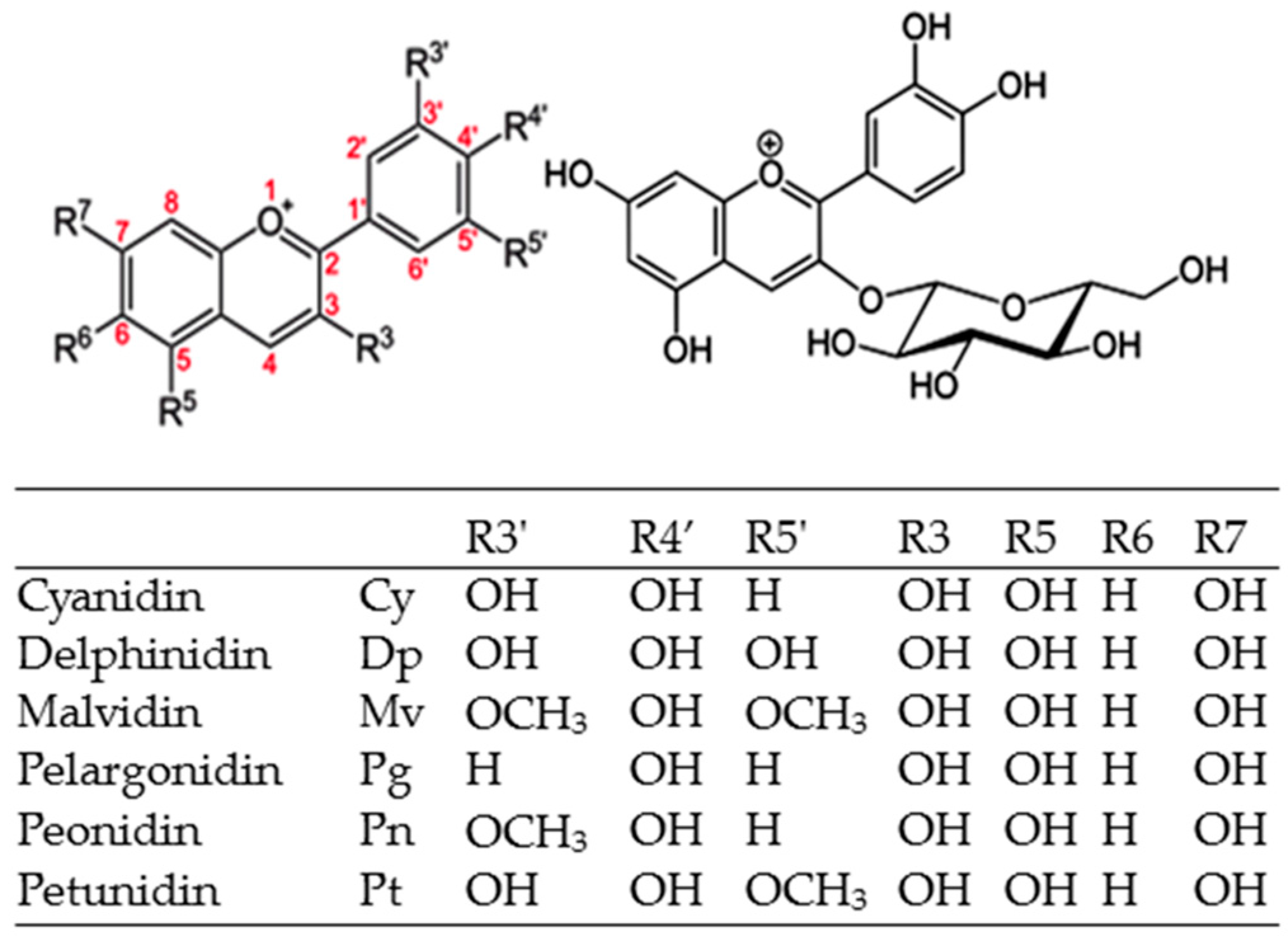
2.1. Chemical Structure
2.2. Color
2.3. Temperature
2.4. pH
2.5. Antioxidant Capacity
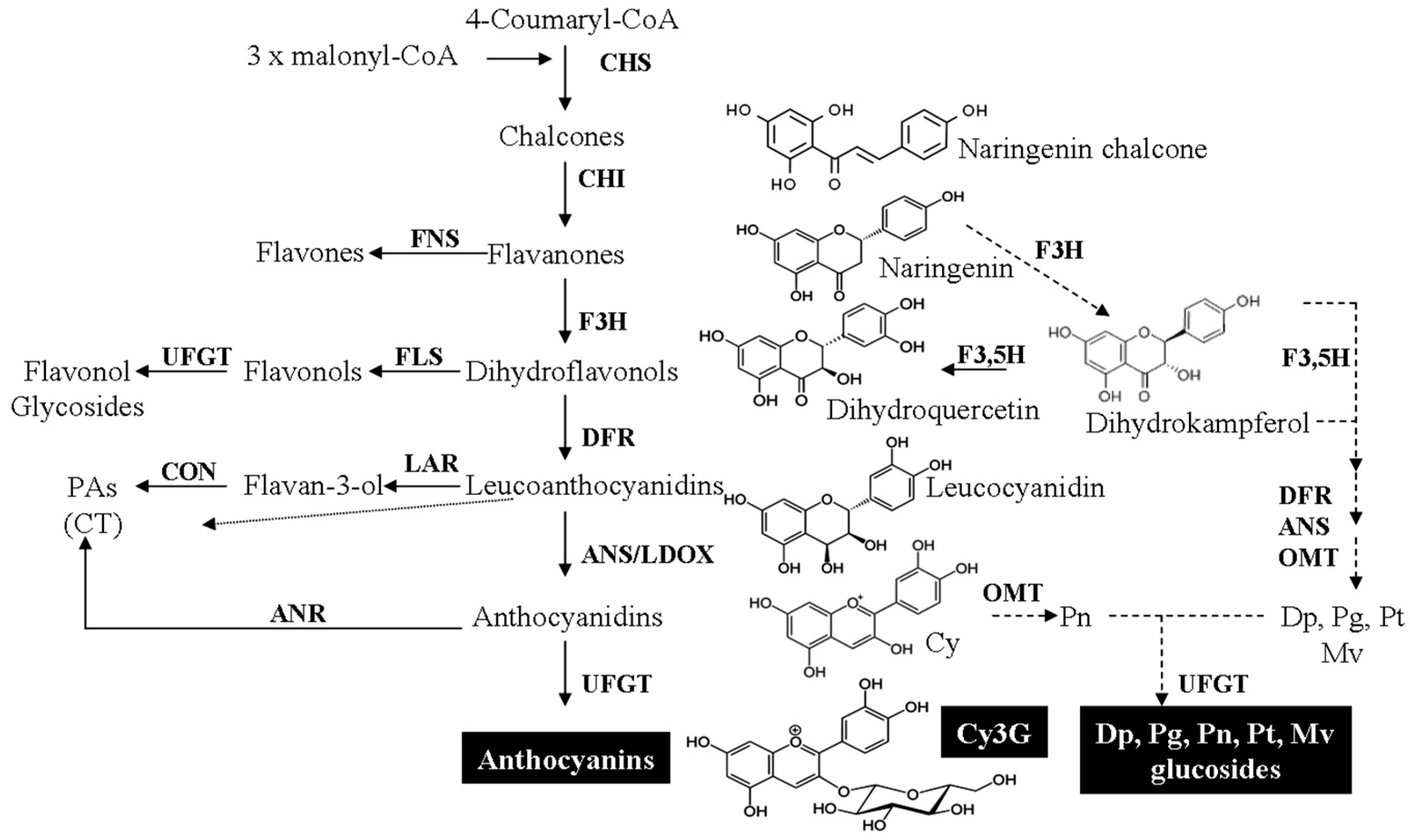
3. Cy3G in Plant Biochemistry
3.1. Biosynthesis
3.2. Dietary Sources
4. Foodomics
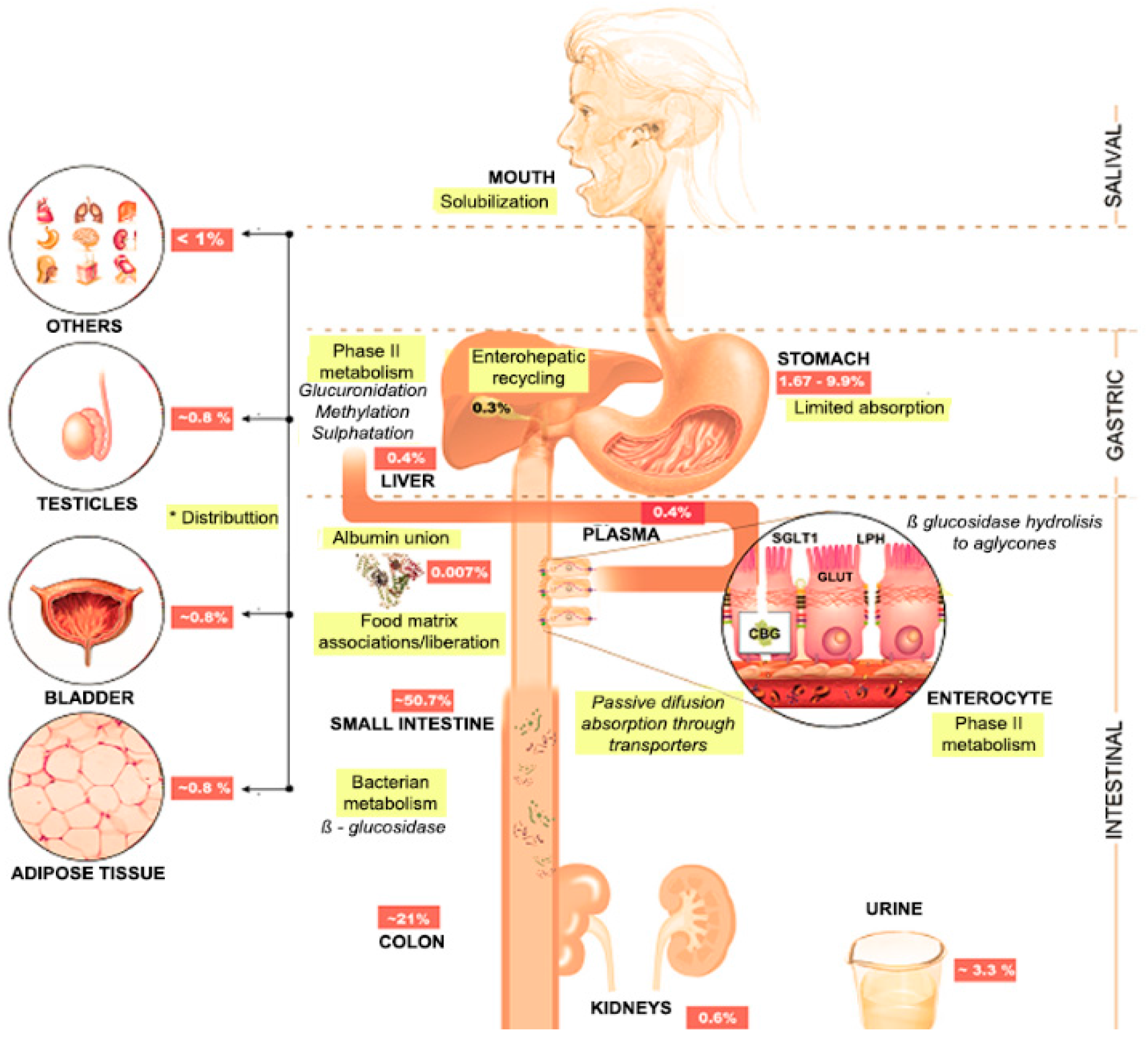
4.1. Oral Cavity
4.2. Stomach
4.3. Small Bowel
4.4. Large Bowel
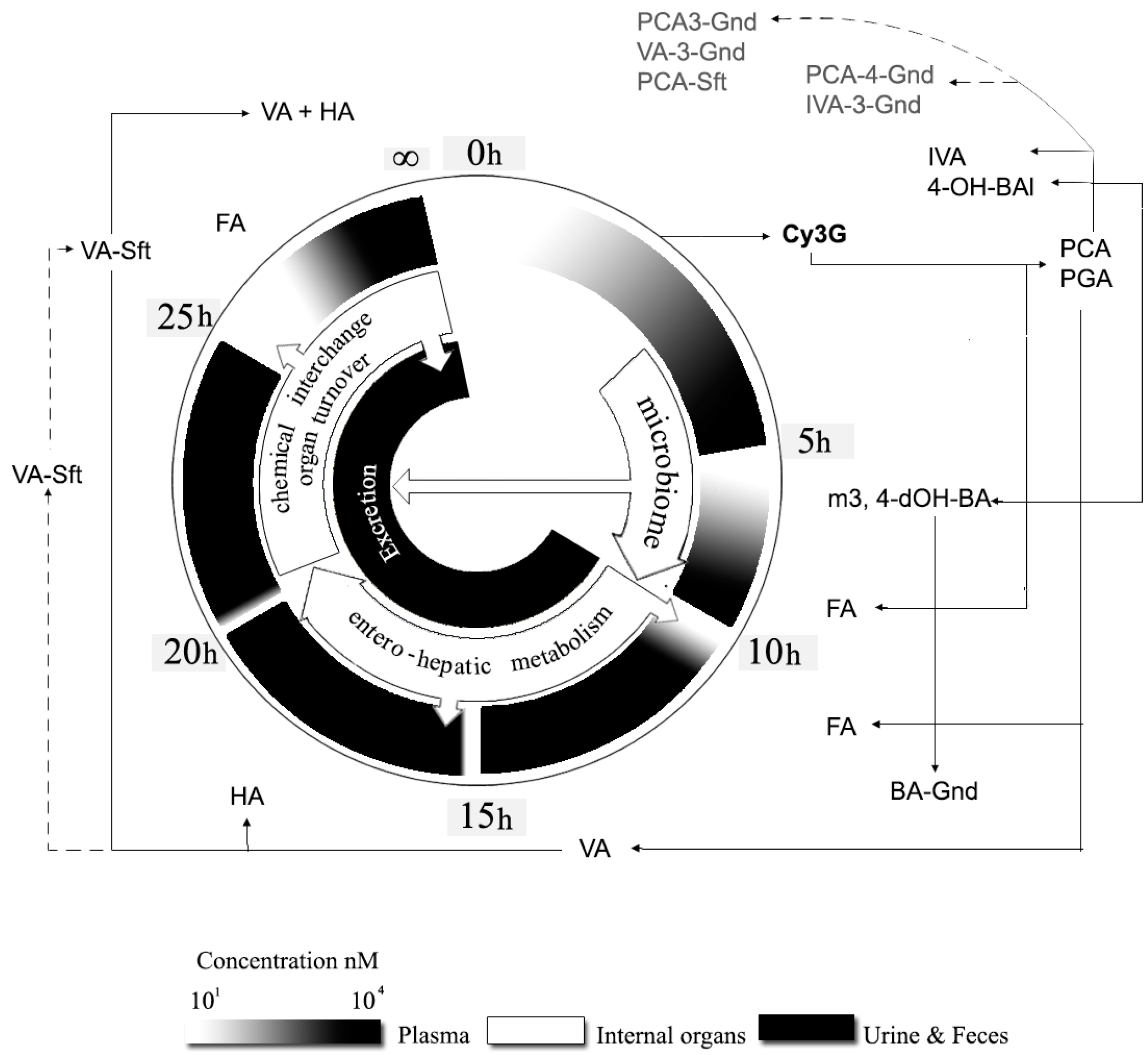
4.5. Splanchnic Metabolism
5. Cy3G Health Effects
5.1. Human Studies
5.2. Cell Lines and Rodent Models
5.3. Physiologically-Relevant Molecular Mechanisms
6. Future Prospects
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ΔA | Hyperchromic effect |
| Δλ | Bathochromic shift |
| ADME | Absorption, distribution, metabolism and excretion |
| ANR | Anthocyanidin reductase |
| ANS | Anthocyanidin synthase |
| ACNs | Anthocyanins |
| Å2 | Polar surface area |
| AOX | Antioxidants |
| ASSWV | Abrasive stripping square wave voltammetry |
| βRA | β-Resorcylic acid |
| BA | Benzoic acid-4-glucoronide |
| BA-Gnd | Benzoic acid-4-glucoronide |
| BRB | Black raspberry |
| BSA | Bovine serum albumin |
| CHI | Chalcone isomerase |
| CHS | Chalcone synthase |
| COMT | Catechol-O-methyltransferase |
| CON | Unknown condensing enzyme(s) |
| CT | Condensed tannins |
| CV | Cyclic voltammetry |
| CVD | Cardiovascular disease |
| Cy | Cyanidin |
| Cy3G | Cyanidin-3-O-glucoside |
| DFR | Dihydroflavonol-4-reductase |
| Dp | Delphinidin |
| Dp3G | Delphinidin-3-O-glucoside |
| Dp3Ga | Delphinidin-3-O-galactoside |
| Dp3R | Delphinidin-3-O-rutinoside |
| DPV | Differential of pulses voltammetry |
| EHM | Entero-hepatic metabolism |
| Epa | Anodic peak potential |
| F3,5H | Flavanone-3,5-hidroxylase |
| F3H | Flavanone-3-hidroxylase |
| FA | Ferulic acid |
| FLS | Flavonol synthase |
| FNS | Flavone synthase |
| GI | Gastrointestinal |
| GLUT1,2,3 | Glucose transporter 1,2,3 |
| HA | Hippuric acid |
| HAS | Human serum albumin |
| Hb | Hemoglobin |
| IVA | Isovainillinic acid |
| IVA-3-Gnd | IVA-3-glucoronide |
| LAR | Leucoanthocyanidin reductase |
| LDOW | Leucoanthocyanidin dioxigenase |
| LogP | Octanol/water partition coefficient |
| LPH | Lactase-phlorizin hydrolase |
| Mb | Myoglobin |
| MCP-1/CCL2 | Monocyte chemoattractant protein-1 |
| MCT1 | Mono-carboxylated transporter 1 |
| Mv | Malvidin |
| Mv3G | Malvidin-3-O-glucoside |
| MW | Molecular weight |
| NCCD | Non-communicable chronic diseases |
| OMT | Malvidin (Mv), O-methyltransferase |
| Pas | Proanthocyanidins |
| PC | Phenolic compounds |
| PCA | Protocatechuic acid |
| PCA-3-Gnd | PCA-3-glucoronide |
| PCA-3-Gnd | PCA-4-glucoronide |
| PCA-Sft | PCA-sulfate |
| Pg | Pelargoddin |
| PGA | Phloroglucinaldehyde |
| PgANS | Pelargodin specific-anthocyanidin synthase |
| Pn | Peonidin |
| Pn3Ga | Peonidin-3-O-galactoside |
| PST | Phenyl sulfotranferases |
| Pt | Petunidin |
| RE | Reference electrode |
| ROS | Reactive oxygen species |
| RSC | Radical scavenging capacity |
| SGLT1 | Na+-glucose transporter |
| UFGT | UDP-glucose-flavonoid-3-o-glucosyltransferase |
| UGT | Uridine-5′-diphosphate glucoronosyltransferases |
| VA | Vainillinic acid |
| VA-3-Gnd | VA-3-glucoronide |
| VA-Sft | VA-sulfate |
| WE | Working electrode |
| 3,4-dOH-BA | Methyl-3,4-dihydroxybenzoate |
| 3RT | Anthocyanidin-3-glycoside rhamnosyl-transferase |
| 4-OH-BA | 4-Hydroxy-benzaldehyde |
References
- Duchnowicz, P.; Broncel, M.; Podsędek, A.; Koter-Michalak, M. Hypolipidemic and antioxidant effects of hydroxycinnamic acids, quercetin, and cyanidin 3-glucoside in hypercholesterolemic erythrocytes (in vitro study). Eur. J. Nutr. 2012, 51, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, M.F.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant. Sci. 2012, 3, 1–15. [Google Scholar]
- Pratheeshkumar, P.; Son, Y.O.; Wang, X.; Divya, S.P.; Joseph, B.; Hitron, J.A.; Wang, L.; Kim, D.; Yin, Y.; Roy, R.V.; et al. Cyanidin-3-glucoside inhibits UVB-induced oxidative damage and inflammation by regulating MAP kinase and NF-κB signaling pathways in SKH-1 hairless mice skin. Toxicol. Appl. Pharmacol. 2014, 280, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, J.; Tanabe, S.; Bergeron, C.; Gafner, S.; Grenier, D. Anthocyanin-rich black currant extract and cyanidin-3-O-glucoside have cytoprotective and anti-inflammatory properties. J. Med. Food 2012, 15, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Yu, S.W.; Baek, S.H.; Nair, K.M.; Bae, O.N.; Bhatt, A.; Kassab, M.; Majid, A. Neuroprotective effect of cyanidin-3-O-glucoside anthocyanin in mice with focal cerebral ischemia. Neurosci. Lett. 2011, 500, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Kovinich, N.; Kayanja, G.; Chanoca, A.; Riedl, K.; Otegui, M.S.; Grotewol, E. Not all anthocyanins are born equal: Distinct patterns induced by stress in Arabidopsis. Planta 2014, 240, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Dong, X.; Zhou, W. Combined effect of pH and high temperature on the stability and antioxidant capacity of two anthocyanins in aqueous solution. Food Chem. 2014, 163, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Marques, F.; de Freitas, V.; Mateus, N. Antioxidant and antiproliferative properties of methylated metabolites of anthocyanins. Food Chem. 2013, 141, 2923–2933. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Gómez, C.; Ageorges, A. Flavonoids: Anthocyanins. In Handbook of Analysis of Active Compounds in Functional Foods, 1st ed.; Nollet, L.M.C., Toldrá, F., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2012; pp. 379–403. [Google Scholar]
- Patras, A.; Brunton, N.P.; O′Donnell, C.; Tiwari, B.K. Effect of thermal processing on anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends. Food Sci. Technol. 2010, 21, 3–11. [Google Scholar] [CrossRef]
- Fang, J. Bioavailability of anthocyanins. Drug. Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Some anthocyanins could be efficiently absorbed across the gastrointestinal mucosa: Extensive presystemic metabolism reduces apparent bioavailability. J. Agric. Food Chem. 2014, 62, 3904–3911. [Google Scholar] [CrossRef] [PubMed]
- De Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Prior, R.L. Systematic identification and characterization of anthocyanins by HPLC-ESI-MS/MS in common foods in the United States: Fruits and berries. J. Agric. Food Chem. 2005, 53, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentration of anthocyanins in common foods in the United States and estimation of normal consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef] [PubMed]
- Pala, Ç.U.; Toklucu, A.K. Effect of UV-C light on anthocyanin content and other quality parameters of pomegranate juice. J. Food Comp. Anal. 2011, 24, 790–795. [Google Scholar] [CrossRef]
- Hernández-Herrero, J.A.; Frutos, M.J. Influence of rutin and ascorbic acid in colour, plum anthocyanins and antioxidant capacity stability in model juices. Food Chem. 2015, 173, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Mansour, R.; Ezzli, B.; Farouk, M. Dyeing properties of wool fabrics dyed with Vitis vinifera L. (black grenache) leaves extract. Fibers Polym. 2013, 14, 786–792. [Google Scholar] [CrossRef]
- Pacheco-Palencia, L.A.; Talcott, S.T. Chemical stability of açai fruit (Euterpe oleracea Mart.) anthocyanins as influenced by naturally occurring and externally added polyphenolic cofactors in model systems. Food Chem. 2010, 118, 17–25. [Google Scholar]
- Kamiloglu, S.; Capanoglu, E.; Grootaert, C.; Van Camp, J. Anthocyanin absorption and metabolism by human intestinal Caco-2 cells-A review. Int. J. Mol. Sci. 2015, 16, 21555–21574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Lee, S.G.; Vance, T.M.; Wang, Y.; Kim, B.; Lee, J.Y.; Chun, O.K.; Bolling, B.W. Bioavailability of anthocyanins and colonic polyphenol metabolites following consumption of Aronia berry extract. Food Chem. 2016, 211, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Ovando, A.; Pacheco-Hernández, M.D.L.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Flamini, R.; de Rosso, M.; Bavaresco, L. Study of grape polyphenols by liquid chromatography-high-resolution mass spectrometry (UHPLC/QTOF) and suspect screening analysis. J. Anal. Methods Chem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A (13)C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, T.; Nakagawa, K.; Kudo, M.; Muraishi, K.; Someva, K. Direct intestinal absorption of red fruit anthocyanins, cyanidin-3-glucoside and cyanidin-3,5-diglucoside into rats and humans. J. Agric. Food Chem. 1999, 47, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Felgines, C.; Krisa, S.; Mauray, A.; Besson, C.; Lamaison, J.L.; Scalbert, A.; Mérillon, J.M.; Texier, O. Radiolabelled cyanidin 3-O-glucoside is poorly absorbed in the mouse. Br. J. Nutr. 2010, 103, 1738–1745. [Google Scholar] [CrossRef] [PubMed]
- Phan, A.D.T.; Netzel, G.; Wang, D.; Flanagan, B.M.; D’Arcy, B.R.; Gidley, M.J. Binding of dietary polyphenols to cellulose: Structural and nutritional aspects. Food Chem. 2015, 171, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.; Pintado, M. In vitro evaluation of the effects of protein-polyphenol-polysaccharide interactions on (+)-catechin and cyanidin-3-glucoside bioaccesibility. Food Funct. 2015, 6, 3444–3453. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zuo, H.; Li, S. Comparison of the interaction between three anthocyanins and human serum albumins by spectroscopy. J. Lumin. 2014, 153, 54–63. [Google Scholar] [CrossRef]
- Tang, L.; Li, S.; Bi, H.; Gao, X. Interaction of cyanidin-3-O-glucoside with three proteins. Food Chem. 2016, 196, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Molinspiration Cheminformatics. Available online: http://www.molinspiration.com/about.html (accessed on 25 July 2016).
- Kosińska-Cagnazzo, A.; Diering, S.; Prim, D.; Andlauer, W. Identification of bioaccessible and uptaken phenolic compounds from strawberry fruits in in vitro digestion/Caco-2 absorption model. Food Chem. 2015, 170, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.; Brás, N.F.; Mateus, N.; de Freitas, V. Understanding the molecular mechanisms of anthocyanin binding to pectin. Langmuir 2014, 30, 8516–8527. [Google Scholar] [CrossRef] [PubMed]
- Urías-Lugo, D.A.; Heredia, J.B.; Muy-Rangel, M.D.; Valdez-Torres, J.B.; Serna-Saldívar, S.O.; Gutiérrez-Uribe, J.A. Anthocyanins and phenolic acids of hybrid and native blue maize (Zea mays L.) extracts and their antiproliferative activity in mammary (MCF7), liver (HepG2), colon (Caco2 and HT29) and prostate (PC3) cancer cells. Plant. Foods Hum. Nutr. 2015, 70, 193–199. [Google Scholar]
- Zhao, X.; Yuan, Z.; Feng, L.; Fang, Y. Cloning and expression of anthocyanin biosynthetic genes in red and white pomegranate. J. Plant Res. 2015, 128, 687–696. [Google Scholar] [CrossRef] [PubMed]
- García-Alonso, M.; Rimbach, G.; Sasai, M.; Nakahara, M.; Matsugo, S.; Uchida, Y.; Rivas-Gonzalo, J.C.; de Pascual-Teresa, A. Electron spin resonance spectroscopy studies on the free radical scavenging activity of wine anthocyanins and pyranoanthocyanins. Mol. Nutr. Food Res. 2015, 49, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Bakowska, A.; Kucharska, A.Z.; Oszmiański, J. The effects of heating, UV irradiation, and storage on stability of the anthocyanin–Polyphenol copigment complex. Food Chem. 2003, 81, 349–355. [Google Scholar] [CrossRef]
- Boulton, R. The copigmentation of anthocyanins and its role in the color of red wine: A critical review. Am. J. Enol. Vitic. 2001, 52, 67–87. [Google Scholar]
- Fazaeli, M.; Yousefi, S.; Emam-Djomeh, Z. Investigation on the effects of microwave and conventional heating methods on the phytochemicals of pomegranate (Punica granatum L.) and black mulberry juices. Food Res. Int. 2013, 50, 568–573. [Google Scholar] [CrossRef]
- Cavalcanti, R.N.; Santos, D.R.; Meireles, M.A.A. Non-thermal stabilization mechanisms of anthocyanins in model and food systems—An overview. Food Res. Int. 2011, 44, 499–509. [Google Scholar] [CrossRef]
- Mildner-Szkudlarz, S.; Bajerska, J.; Górnaś, P.; Segliņa, D.; Pilarska, A.; Jesionowski, T. Physical and bioactive properties of muffins enriched with raspberry and cranberry pomace powder: A promising application of fruit by-products rich in biocompounds. Plant Foods Hum. Nutr. 2016, 71, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Slavin, M.; Lu, Y.; Kaplan, N.; Yu, L.L. Effects of baking on cyanidin-3-glucoside content and antioxidant properties of black and yellow soybean crackers. Food Chem. 2013, 141, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Rawson, A.; Patras, A.; Tiwari, B.K.; Noci, F.; Koutchma, T.; Brunton, N. Effect of thermal and non-thermal processing technologies on the bioactive content of exotic fruits and their products: Review of recent advances. Food Res. Int. 2011, 44, 1875–1887. [Google Scholar] [CrossRef]
- Wang, H. Rapid quantitative analysis of individual anthocyanin content based on high-performance liquid chromatography with diode array detection with the pH differential method. J. Sep. Sci. 2014, 37, 2535–2544. [Google Scholar] [CrossRef] [PubMed]
- Janeiro, P.; Brett, A.M.O. Redox behavior of anthocyanins present in Vitis vinifera L. Electroanalysis 2007, 19, 1779–1786. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; pp. 77–79. [Google Scholar]
- Moran, V.F.E.; Boggetti, H.J.; Zampini, I.C.; Ordoñez, R.M.; Isla, M.I.; Alvarez, R.M.S.; de Rosso, V.; Mercadante, A.Z.; Borsarelli, C.D. Singlet oxygen quenching and radical scavenging capacities of structurally-related flavonoids present in Zuccagnia punctata Cav. Free Radic. Res. 2009, 43, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Komorsky-Lovric, S.; Novak, I. Abrasive stripping square-wave voltammetry of blackberry, raspberry, strawberry, pomegranate, and sweet and blue potatoes. J. Food Sci. 2011, 76, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.L.; Yu, Y.Q.; Chen, Z.J.; Wen, G.S.; Wei, F.G.; Zheng, Q.; Wang, C.-D.; Xiao, X.L. Stability-increasing effects of anthocyanin glycosyl acylation. Food Chem. 2017, 214, 119–128. [Google Scholar] [CrossRef] [PubMed]
- De Lima, A.A.; Sussuchi, E.M.; Giovani, W.F. Electrochemical and antioxidant properties of anthocyanins and anthocyanidins. Croatia Chem. Acta 2007, 80, 29–34. [Google Scholar]
- Moncada, M.C.; de Mesquita, M.F.; dos Santos, M.M.C. Electrochemical oxidation of the synthetic anthocyanin analogue 4-methyl-7,8-dihydroxyflavylium salt. J. Electroanal. Chem. 2009, 636, 60–67. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Yordi, E.G.; Pérez, E.M.; Matos, M.J.; Villares, E.U. Antioxidant and pro-oxidant effects of polyphenolic compounds and structure-activity relationship evidence. In Nutrition, Well-Being and Health; Bouayed, J., Bohn, T., Eds.; InTech: Rijeka, Croatia, 2012; pp. 23–48. [Google Scholar]
- Hassimotto, N.M.A.; Genovese, M.I.; Lajolo, F.M. Absorption and metabolism of cyanidin-3-glucoside and cyanidin-3-rutinoside extracted from wild mulberry (Morus nigra L.) in rats. Nutr. Res. 2008, 28, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Wu, V.C.; Tyler, S.; Edwards, K. Antimicrobial action of the American cranberry constituents; phenolics, anthocyanins, and organic acids against Escherichia coli O157:H7. Int. J. Food Microbiol. 2010, 139, 102–107. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Mu, L.; Yang, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [PubMed]
- Ploenlap, P.; Pattanagul, W. Effects of exogenous abscisic acid on foliar anthocyanin accumulation and drought tolerance in purple rice. Biologia 2015, 70, 915–921. [Google Scholar] [CrossRef]
- Cisowska, A.; Wojnicz, D.; Hendrich, A.B. Anthocyanins as antimicrobial agents of natural plant origin. Nat. Prod. Commun. 2011, 6, 149–156. [Google Scholar] [PubMed]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Avramenko, T.V.; Tsitsiashvili, G.S. Critical analysis of protein signaling networks involved in the regulation of plant secondary metabolism: Focus on anthocyanins. Crit. Rev. Biotechnol. 2016, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dossett, M.; Lee, J.; Finn, C.E. Characterization of a novel anthocyanin profile in wild black raspberry mutants: An opportunity for studying the genetic control of pigment and color. J. Funct. Foods 2011, 3, 207–214. [Google Scholar] [CrossRef]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.J.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 2007, 49, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Pierantoni, L.; Dondini, L.; De Franceschi, P.; Musacchi, S.; Winkel, B.S.J.; Sansavini, S. Mapping of an anthocyanin-regulating MYB transcription factor and its expression in red and green pear, Pyrus communis. Plant Physiol. Biochem. 2010, 48, 1020–1026. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Du, L.; Li, L.; Kalt, W.; Palmer, L.C.; Fillmore, S.; Zhang, Y.; Zang, Z.Q.; Li, X. Quantitative changes in proteins responsible for flavonoid and anthocyanin biosynthesis in strawberry fruit at different ripening stages: A targeted quantitative proteomic investigation employing multiple reaction monitoring. J. Proteom. 2015, 122, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, L.; Perez-Lopez, P.; Giampieri, F.; Alvarez-Suarez, J.M.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Quiles, J.L.; Mezzetti, B.; Battino, M. The genetic aspects of berries: From field to health. J. Sci. Food Agric. 2016, 96, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Vogiatzoglou, A.; Mulligan, A.A.; Lentjes, M.A.; Luben, R.N.; Spencer, J.P.; Schroeter, H.; Khaw, H.; Kuhnle, G.G. Flavonoid intake in European adults (18 to 64 years). PLoS ONE 2015, 10, e0128132. [Google Scholar] [CrossRef] [PubMed]
- Department of Agriculture, Agricultural Research Service. USDA National Nutrient Database for Standard Reference, Release 28. 2005–2011. Available online: http://www.ars.usda.gov/Services/docs.htm?docid = 8964 (accessed on 30 July 2016). [Google Scholar]
- Diaconeasa, Z.; Leopold, L.; Rugină, D.; Ayvaz, H.; Socaciu, C. Antiproliferative and antioxidant properties of anthocyanin rich extracts from blueberry and blackcurrant Juice. Int. J. Mol. Sci. 2015, 16, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Niño-Medina, G.; Muy-Rangel, D.; Gardea-Bejar, A.; González-Aguilar, G.; Heredia, B.; Baez-Sanudo, M.; Siller-Cepeda, J.; Velez de la Rocha, R. Nutritional and nutraceutical components of commercial eggplant types grown in Sinaloa, México. Not. Bot. Horti Agrobot. Cluj-Napoca 2014, 42, 538–544. [Google Scholar] [CrossRef]
- Nimptsch, K.; Zhang, X.; Cassidy, A.; Song, M.; O’Reilly, É.J.; Lin, J.H.; Pischon, T.; Rimm, E.B.; Willet, W.C.; Fuchs, C.S.; et al. Habitual intake of flavonoid subclasses and risk of colorectal cancer in 2 large prospective cohorts. Am. J. Clin. Nutr. 2016, 103, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastian, R.S.; Enns, C.W.; Goldman, J.D.; Martin, C.L.; Steinfeldt, L.C.; Murayi, T.; Moshfegh, A.J. A new database facilitates characterization of flavonoid intake, sources, and positive associations with diet quality among US Adults. J. Nutr. 2015, 145, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Stephania, U.; Micek, A.; Stefler, D.; Bobak, M.; Paja, A. Dietary polyphenols are inversely associated with metabolic syndrome in Polish adults of the HAPIEE study. Eur. J. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.; Shin, S.; Joung, H. Estimation of dietary flavonoid intake and major food sources of Korean adults. Br. J. Nutr. 2016, 115, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.J.; Dwyer, J.T.; Jacques, P.F.; McCullough, M.L. Improving the estimation of flavonoid intake for study of health outcomes. Nutr. Rev. 2015, 73, 553–576. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, F.; Bordoni, A. Foodomics: A new comprehensive approach to food and nutrition. Genes Nutr. 2013, 8, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Velderrain-Rodríguez, G.; Palafox, H.; Wall-Medrano, A.; Ayala-Zavala, J.F.; Chen, C.Y.O.; Robles-Sánchez, R.M.; Astiazaran-García, H.; Álvarez-Parrilla, H.; González-Aguilar, G.A. Phenolic compounds: Their journey after intake. Food Funct. 2014, 5, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Kopf-Bolanz, K.A.; Schwander, F.; Gijs, M.; Vergères, G.; Portmann, R.; Egger, L. Validation of an in vitro digestive system for studying macronutrient decomposition in humans. J. Nutr. 2012, 142, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Food oral processing: Some important underpinning principles of eating and sensory perception. Food Struct. 2014, 1, 91–105. [Google Scholar] [CrossRef]
- Carpenter, G.H. The secretion, components, and properties of saliva. Ann. Rev. Food Sci. Technol. 2013, 4, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Wiese, S.; Gärtner, S.; Rawel, H.M.; Winterhalter, P.; Kulling, S.E. Protein interactions with cyanidin-3-glucoside and its influence on α-amylase activity. J. Sci. Food Agric. 2009, 89, 33–40. [Google Scholar] [CrossRef]
- Kamonpatana, K.; Giusti, M.M.; Chitchumroonchokchai, C.; Moren Cruz, M.; Riedl, K.M.; Kumar, P.; Failla, M.L. Susceptibility of anthocyanins to ex vivo degradation in human saliva. Food Chem. 2012, 135, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Mallery, S.R.; Budendorf, D.E.; Larsen, M.P.; Pei, P.; Tong, M.; Holpuch, A.S.; Larsen, P.E.; Stoner, G.D.; Fields, H.W.; Chan, K.K.; et al. Effects of human oral mucosal tissue, saliva, and oral microflora on intraoral metabolism and bioactivation of black raspberry anthocyanins. Cancer Prev. Res. 2011, 4, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, T.J.; Uhrig, L.K.; Pearl, D.K.; Casto, B.C.; Warner, B.M.; Clinton, S.K.; Sardo-Molmenti, C.L.; Ferguson, J.M.; Daly, B.T.; Riedl, K.; et al. Suppression of pro-inflammatory and pro-survival biomarkers in oral cancer patients consuming a black raspberry phytochemical-rich troche. Cancer Prev. Res. 2016, 9, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Fernandes, I.; Brás, N.F.; Faria, A.; De Freitas, V.; Calhau, C.; Mateus, N. Experimental and theoretical data on the mechanism by which red wine anthocyanins are transported through a human MKN-28 gastric cell model. J. Agric. Food Chem. 2015, 63, 7685–7692. [Google Scholar] [CrossRef] [PubMed]
- Del Bo’, C.; Riso, P.; Brambilla, A.; Gardana, C.; Rizzolo, A.; Simonetti, P.; Bertolo, G.; Klimis-Zacas, D.; Porrini, M. Blanching improves anthocyanin absorption from highbush blueberry (Vaccinium corymbosum L.) purée in healthy human volunteers: A pilot study. J. Agric. Food Chem. 2012, 60, 9298–9304. [Google Scholar] [PubMed]
- McGhie, T.K.; Walton, M.C. The bioavailability and absorption of anthocyanins: Towards a better understanding. Mol. Nutr. Food Res. 2007, 51, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Passamonti, S.; Terdoslavich, M.; Franca, R.; Vanzo, A.; Tramer, F.; Braidot, E.; Petrussa, E.; Angelo, V. Bioavailability of flavonoids: A review of their membrane transport and the function of bilitranslocase in animal and plant organisms. Curr. Drug Metab. 2009, 10, 369–394. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.B.; Feng, D.; Song, G.; Li, H.W.; Tang, H.W.; Ling, W.H. The role of sodium-dependent glucose transporter 1 and glucose transporter 2 in the absorption of cyanidin-3-O-β-glucoside in Caco-2 cells. Nutrients 2014, 6, 4165–4177. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Woo, H.; Park, M.; Rhee, K.J.; Moon, C.; Lee, D.; Seo, W.D.; Kim, J.B. Cyanidin 3-O-glucoside reduces Helicobacter pylori VacA-induced cell death of gastric KATO III cells through inhibition of the SecA pathway. Int. J. Med. Sci. 2014, 11, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Effects of anthocyanidin on the inhibition of proliferation and induction of apoptosis in human gastric adenocarcinoma cells. Food Chem. Toxicol. 2005, 43, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Felgines, C.; Talavera, S.; Texier, O.; Bensson, C.; Fogliano, V.; Lamaison, J.L.; la Fauci, L.; Galvano, G.; Remes, C.; Galvano, F. Absorption and metabolism of red orange anthocyanins in rats. Br. J. Nutr. 2006, 95, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mateos, A.; Vauzour, D.; Krueger, C.G.; Shanmuganayagam, D.; Reed, J.; Calani, L.; Mena, P.; del Rio, D.; Crozier, A. Bioavailability, bioactivity and impact on health of dietary flavonoids and related compounds: An update. Arch. Toxicol. 2014, 88, 1803–1853. [Google Scholar] [CrossRef] [PubMed]
- Woodward, G.M.; Needs, P.W.; Kay, C.D. Anthocyanin-derived phenolic acids form glucuronides following simulated gastrointestinal digestion and microsomal glucuronidation. Mol. Nutr. Food Res. 2011, 55, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, S.; Rudloff, S.; Asseburg, H.; Borsch, C.; Fröhling, B.; Unger, F.; Dold, S.; Spengler, B.; Romp, A.; Kunz, C. Uptake and bioavailability of anthocyanins and phenolic acids from grape/blueberry juice and smoothie in vitro and in vivo. Br. J. Nutr. 2015, 113, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Ribnicky, D.M.; Roopchand, D.E.; Oren, A.; Grace, M.; Poulev, A.; Lila, M.A.; Havenaar, R.; Raskin, I. Effects of a high fat meal matrix and protein complexation on the bioaccessibility of blueberry anthocyanins using the TNO gastrointestinal model (TIM-1). Food Chem. 2014, 142, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Zhang, Y.; Zhou, W. In vitro and in silico studies of the inhibition of anthocyanins against pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Cardona, J.A.; Mertens-Talcott, S.U.; Talcott, S.T. Phospholipids and terpenes modulate Caco-2 transport of açaí anthocyanins. Food Chem. 2015, 175, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.C.; McGhie, T.K.; Reynolds, G.W.; Hendriks, W.H. The flavonol quercetin-3-glucoside inhibits cyanidin-3-glucoside absorption in vitro. J. Agric. Food Chem. 2006, 54, 4913–4920. [Google Scholar] [CrossRef] [PubMed]
- Hanske, L.; Engst, W.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Contribution of gut bacteria to the metabolism of cyanidin 3-glucoside in human microbiota-associated rats. Br. J. Nutr. 2013, 109, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Valdés, L.; Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. The relationship between phenolic compounds from diet and microbiota: Impact on human health. Food Funct. 2015, 6, 2424–2439. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, O.; Botting, N.P.; Kay, C. A gram scale synthesis of a multi-13C-labelled anthocyanin, [6,8,10,3′,5′-13C5] cyanidin-3-glucoside, for use in oral tracer studies in humans. Chem. Commun. 2011, 47, 10596–10598. [Google Scholar] [CrossRef] [PubMed]
- Khymenets, O.; Andres-Lacueva, C.; Urpi-Sarda, M.; Vázquez-Fresno, R.; Mart, M.M.; Reglero, G.; Torres, M.; Llorach, R. Metabolic fingerprint after acute and under sustained consumption of a functional beverage based on grape skin extract in healthy human subjects. Food Funct. 2015, 6, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). Scientific opinion on the re-evaluation of anthocyanins (e 163) as food additives. EFSA J. 2013, 11, 3145. [Google Scholar]
- Charoensin, S.; Taya, S.; Wongpornchai, S.; Wongpoomchai, R. Assessment of genotoxicity and antigenotoxicity of an aqueous extract of Cleistocalyx nervosum var. paniala in in vitro and in vivo models. Interdiscip. Toxicol. 2012, 5, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Rogers, G.; Peterson, J.J.; Dwyer, J.T.; Lin, H.; Jacques, P.F. Higher dietary anthocyanin and flavonol intakes are associated with anti-inflammatory effects in a population of US adults. Am. J. Clin. Nutr. 2015, 102, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.J.; Cassidy, A.; Litonjua, A.A.; Sparrow, D.; Vokonas, P.; Schwartz, J. Dietary anthocyanin intake and age-related decline in lung function: Longitudinal findings from the VA Normative Aging Study. Am. J. Clin. Nutr. 2016, 103, 542–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponzo, V.; Goitre, I.; Fadda, M.; Gambino, R.; De Francesco, A.; Soldati, L.; Gentile, L.; Magistroni, P.; Cassander, M.; Bo, S. Dietary flavonoid intake and cardiovascular risk: A population-based cohort study. J. Transl. Med. 2015, 13, 218. [Google Scholar] [CrossRef] [PubMed]
- Hassellund, S.S.; Flaa, A.; Kjeldsen, S.E.; Seljeflot, I.; Karlsen, A.; Erlund, I.; Rostrup, M. Effects of anthocyanins on cardiovascular risk factors and inflammation in pre-hypertensive men: A double-blind randomized placebo-controlled crossover study. J. Hum. Hypertens. 2013, 27, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Bertoglio, J.C.; Zarrelli, A.; Pina, R.; Scapagnini, G. A randomized clinical trial evaluating the efficacy of an anthocyanin–maqui berry extract (Delphinol®) on oxidative stress biomarkers. J. Am. Coll. Nutr. 2015, 34. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y.; Li, G.; Xiao, Y.; et al. Anti-inflammatory effect of purified dietary anthocyanin in adults with hypercholesterolemia: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J.; Sandhya, K.; Ellis, C.L.; Tadapaneni, R.; Kappagoda, C.T.; Burton-Freeman, B.M. Strawberry anthocyanin and its association with postprandial inflammation and insulin. Br. J. Nutr. 2011, 106, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mateos, A.; Rendeiro, C.; Bergillos-Meca, T.; Tabatabaee, S.; George, T.W.; Heiss, C.; Spencer, J.P. Intake and time dependence of blueberry flavonoid-induced improvements in vascular function: A randomized, controlled, double-blind, crossover intervention study with mechanistic insights into biological activity. Am. J Clin. Nutr. 2013, 98, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xia, M.; Yang, Y.; Liu, F.; Li, Z.; Hao, Y.; Mi, M.; Jinm, T.; Ling, W. Purified anthocyanin supplementation improves endothelial function via NO-cGMP activation in hypercholesterolemic individuals. Clin. Chem. 2011, 57, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Arnold, M.; Huang, Y.W.; Sardo, C.; Seguin, C.; Martin, E.; Huang, T.H.; Riedl, K.; Schwartz, S.; Frankel, W.; et al. Modulation of genetic and epigenetic biomarkers of colorectal cancer in humans by black raspberries: A phase I pilot study. Clin. Cancer Res. 2011, 17, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, R.; Russo, A.; Galvano, F.; Galvano, G.; Barcellona, M.L.; Volti, G.L.; Vanella, A. Cyanidin and cyanidin 3-O-β-d-glucoside as DNA cleavage protectors and antioxidants. Cell Biol. Toxicol. 2003, 19, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.Q.; Wu, X.Y.; Zhou, X.; Zhu, J.; Ma, L.Y. Cyanidin-3-glucoside attenuates body weight gain, serum lipid concentrations and insulin resistance in high-fat diet-induced obese rats. Chin. J. Contemp. Pediatr. 2014, 16, 534–538. [Google Scholar]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Guo, J.; Jiang, X.; Li, Z.; Ling, W. Cyanidin-3-O-β-glucoside, a typical anthocyanin, exhibits antilipolytic effects in 3T3-L1 adipocytes during hyperglycemia: Involvement of FoxO1-mediated transcription of adipose triglyceride lipase. Food Chem. Toxicol. 2012, 50, 3040–3047. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Liu, G.; Zhong, R.; Wang, Y.; Wang, D.; Xia, M. Cyanidin-3-O-β-glucoside regulates fatty acid metabolism via an AMP-activated protein kinase-dependent signaling pathway in human HepG2 cells. Lipids Health Dis. 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Ziberna, L.; Tramer, F.; Moze, S.; Vrhovsek, U.; Mattivi, F.; Pssamonti, S. Transport and bioactivity of cyanidin 3-glucoside into the vascular endothelium. Free Radic. Biol. Med. 2012, 52, 1750–1759. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Ueno, Y.; Yoshikawa, T.; Kojo, H.; Osawa, T. Microarray profiling of gene expression in human adipocytes in response to anthocyanins. Biochem. Pharmacol. 2006, 71, 1184–1197. [Google Scholar] [CrossRef] [PubMed]
- Björk, C.; Wilhelm, U.; Mandrup, S.; Larsen, B.D.; Bordoni, A.; Hedén, P.; Rydén, M.; Arner, P.; Laurencikiene, J. Effects of selected bioactive food compounds on human white adipocyte function. Nutr. Metab. (Lond.) 2016, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, T.; Ueno, Y.; Kojo, H.; Yoshikawa, T.; Osawa, T. Gene expression profile of isolated rat adipocytes treated with anthocyanins. Biochim. Biophys. Acta 2005, 1733, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Pimpão, R.C.; Ventura, M.R.; Ferreira, R.B.; Williamson, G.; Santos, C.N. Phenolic sulfates as new and highly abundant metabolites in human plasma after ingestion of a mixed berry fruit purée. Br. J. Nutr. 2015, 113, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Chemaxon. Available online: https://www.chemaxon.com/ (accessed on 10 August 2016).
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0—The human metabolome database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Phenol-Explorer. Data Base on Polyphenol Content in Foods, Version 3.6 (Internet Database). Available online: http://phenol-explorer.eu/ (accessed on 10 August 2016).
- Xu, M.; Bower, K.A.; Wang, S.; Frank, J.A.; Chen, G.; Ding, M.; Luo, J. Cyanidin-3-glucoside inhibits ethanol-induced invasion of breast cancer cells overexpressing ErbB2. Mol. Cancer 2010, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Pyo, M.Y.; Yoon, S.J.; Yu, Y.; Park, S.; Jin, M. Cyanidin-3-glucoside suppresses Th2 cytokines and GATA-3 transcription factor in EL-4 T cells. Biosci. Biotechnol. Biochem. 2014, 78, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Yang, J.S.; Ha, T.J.; Yoon, S.H. Cyanidin-3-glucoside inhibits ATP-induced intracellular free Ca2+ concentration, ROS formation and mitochondrial depolarization in PC12 cells. Korean J. Physiol. Pharmacol. 2014, 18, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.; Paixão, J.; Nunes, C.; Dinis, T.C.; Almeida, L.M. Cyanidin-3-glucoside suppresses cytokine-induced inflammatory response in human intestinal cells: Comparison with 5-aminosalicylic acid. PLoS ONE 2013, 8, e73001. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.A.; Xia, M.; Gao, S.; Li, D.; Zhang, Y.; Kin, T.; Ling, W. Cyanidin-3-O-β-glucoside upregulates hepatic cholesterol 7α-hydroxylase expression and reduces hypercholesterolemia in mice. Mol. Nutr. Food Res. 2012, 56, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Skemiene, K.; Liobikas, J.; Borutait, V. Anthocyanins as substrates for mitochondrial complex I—protective effect against heart ischemic injury. FEBS J. 2015, 282, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Nair, M.G.; Claycombe, K.J. Synergistic inhibition of interleukin-6 production in adipose stem cells by tart cherry anthocyanins and atorvastatin. Phytomedicine 2012, 19, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Gao, J.; Zhang, R. Study on anti-tumor effect of cyanidin-3-glucoside on ovarian cancer. Zhongguo Zhong Yao Za Zhi 2012, 37, 1651–1654. [Google Scholar] [PubMed]
- Wu, T.; Qi, X.; Liu, Y.; Guo, J.; Zhu, R.; Chen, W.; Yu, T. Dietary supplementation with purified mulberry (Morus australis Poir) anthocyanins suppresses body weight gain in high-fat diet fed C57BL/6 mice. Food Chem. 2013, 141, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Wang, Y.; Liu, Y.; Xia, M. Supplementation of cyanidin-3-O-β-glucoside promotes endothelial repair and prevents enhanced atherogenesis in diabetic apolipoprotein E–deficient mice. J. Nutr. 2013, 143, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Hassimotto, N.M.; Moreira, V.; do Nascimento, N.G.; Souto, P.C.; Teixeira, C.; Lajolo, F.M. Inhibition of carrageenan-induced acute inflammation in mice by oral administration of anthocyanin mixture from wild mulberry and cyanidin-3-glucoside. BioMed Res. Int. 2013, 2013, 146716. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.D.; Zhang, B.; Zhang, J.K.; Xu, C.J.; Wu, Y.L.; Li, X.; Chen, K.S. Cyanidin-3-glucoside-rich extract from Chinese bayberry fruit protects pancreatic β cells and ameliorates hyperglycemia in streptozotocin-induced diabetic mice. J. Med. Food 2012, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Wang, D.; Yang, Y.; Xia, M.; Li, D.; Li, G.; Zhu, Y.; Xiao, Y.; Ling, W. Cyanidin-3-O-β-glucoside improves obesity and triglyceride metabolism in KK-Ay mice by regulating lipoprotein lipase activity. J. Sci. Food Agric. 2011, 91, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhou, J.; Wu, Z.; Wang, X.; Liu, L.; Yao, C. Cyanidin-3-O-β-Glucoside ameliorates ethanol-induced acute liver injury by attenuating oxidative stress and apoptosis: The role of SIRT1/FOXO1 signaling. Alcohol. Clin. Exp. Res. 2016, 40, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jeong, E.; Paik, S.S.; Jeon, J.H.; Jung, S.W.; Kim, H.B.; Kim, M.; Chun, M.H.; Kim, I.B. Cyanidin-3-glucoside extracted from mulberry fruit can reduce N-methyl-N-nitrosourea-induced retinal degeneration in rats. Curr. Eye Res. 2013, 39, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ke, Z.; Liu, Y.; Wang, X.; Fan, Z.; Chen, G.; Xu, M.; Bower, K.A.; Frank, J.A.; Ou, X.; Shi, X.; et al. Cyanidin-3-glucoside ameliorates ethanol neurotoxicity in the developing brain. J. Neurosci. Res. 2011, 89, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Zhang, J.; Qin, M. Protective effect of cyanidin 3-O-glucoside on beta-amyloid peptide-induced cognitive impairment in rats. Neurosci. Lett. 2013, 534, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.; Burkard, M.; Leischner, C.; Lauer, U.M.; Frank, J.; Venturelli, S. Epigenetic activities of flavonoids in the prevention and treatment of cancer. Clin. Epigenetics 2015, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kiemlian Kwee, J. Yin and Yang of polyphenols in cancer prevention: A short review. Anti-Cancer Agents Med. Chem. 2016, 16, 832–840. [Google Scholar]
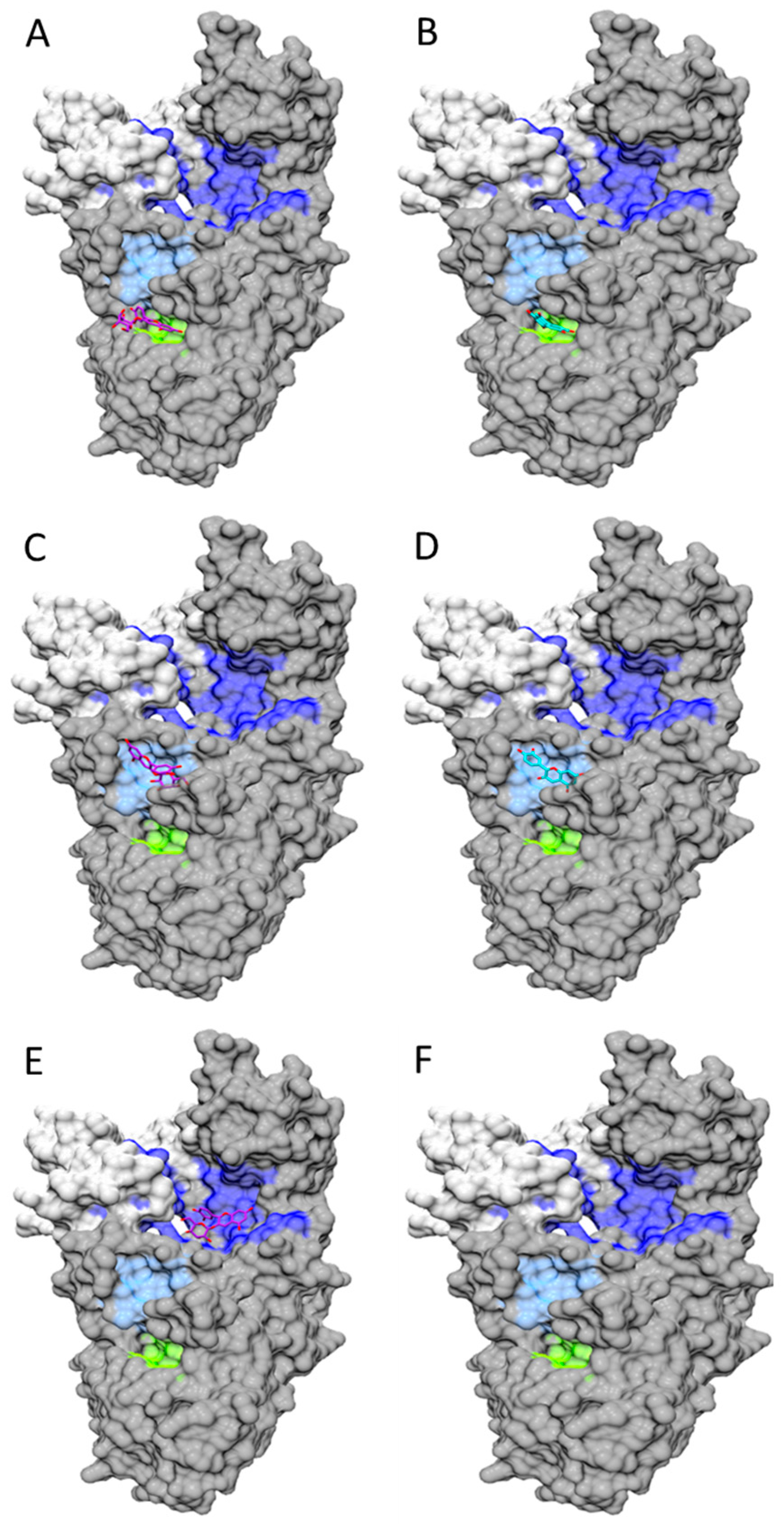
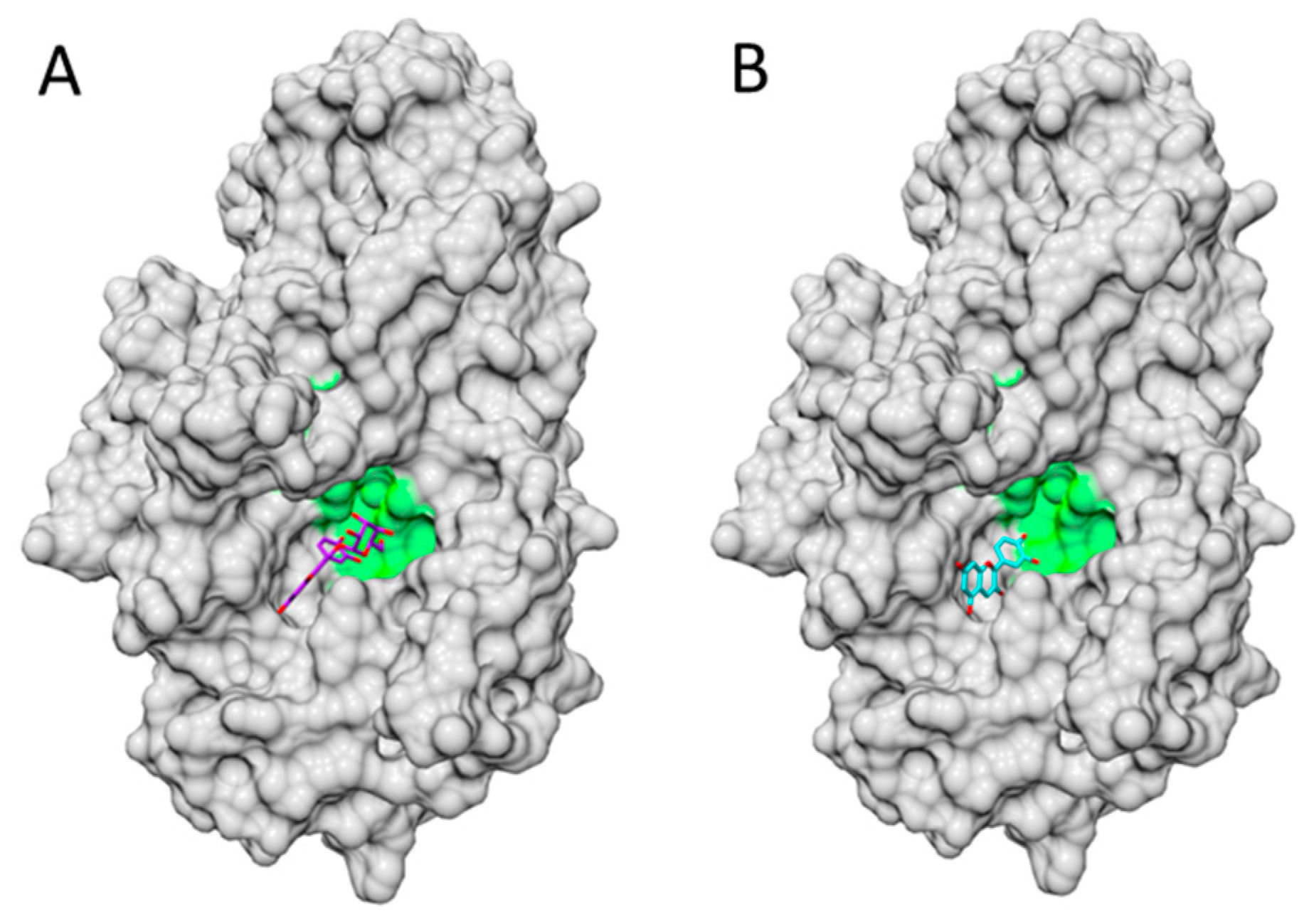
- Zhang, C.; Guo, X.; Cai, W.; Ma, Y.; Zhao, X. Binding characteristics and protective capacity of Cyanidin-3-glucoside and its aglycon to calf thymus DNA. J. Food Sci. 2016, 80, H889–H893. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.H.; Wang, J.; Zhu, Y.Y.; Chen, J. Characterization of intermolecular interaction between cyanidin-3-glucoside and bovine serum albumin: Spectroscopic and molecular docking methods. Luminescence 2014, 29, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Cahyana, Y.; Gordon, M.H. Interaction of anthocyanins with human serum albumin: Influence of pH and chemical structure on binding. Food Chem. 2013, 141, 2278–2285. [Google Scholar] [CrossRef] [PubMed]
- Sheng, F.; Wang, Y.; Zhao, X.; Tian, N.; Hu, H.; Li, P. Separation and identification of anthocyanin extracted from mulberry fruit and the pigment binding properties toward human serum albumin. J. Agric. Food Chem. 2014, 62, 6813–6819. [Google Scholar] [CrossRef] [PubMed]
- Adisakwattana, S.; Yibchok-Anun, S.; Charoenlertkul, P.; Wongsasiripat, N. Cyanidin alleviates postprandial hyperglycemia and its synergism with acarbose by inhibition of intestinal α glucosidase. J. Clin. Biochem. Nutr. 2011, 49, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Díaz, R.; Madrid-Espinoza, J.; Salinas-Cornejo, J.; González-Villanueva, E.; Ruiz-Lara, S. Differential roles for VviGST1, VviGST3, and VviGST4 in proanthocyanidin and anthocyanin transport in Vitis vinífera. Front Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Name | [M]+ (m/z) | MS/MS (m/z) | Solubility (mg/mL) | LogP | Å2 |
|---|---|---|---|---|---|---|
| Cy | Cyanidin (anthocyanidin) | 287 | 0.049 | 3.05 | 114.3 | |
| Cy3A | Cy-3-arabinoside (pentose) | 419 | 287 | 0.41 | 1.06 | 173.21 |
| Cy3X | Cy-3-xyloside | 419 | 287 | 0.41 | 1.06 | 173.2 |
| Cy3G | Cy-3-glucoside | 449 | 287 | 0.6 | 0.39 | 193.4 |
| Cy3Ga | Cy-3-galactoside | 449 | 287 | -- | 0.24 | 193.4 |
| Cy3Aga | Cy-3-(6′′-acetyl)-galactoside | 491 | 287 | 0.39 | 0.82 | 199.5 |
| Cy3Sa | Cy-3-sambubioside | 518 | 287 | -- | -- | -- |
| Cy3,3′′MG | Cy-3-(3′′-malonyl)-glucoside | 535 | 287 | 0.47 | 0.68 | 236.8 |
| Cy3,6′′MG | Cy-3-(6′′-malonyl)-glucoside | 535 | 449/287 | 0.45 | 0.68 | 236.8 |
| Cy3Sa | Cy-3-sambubioside | 581 | 287 | 1.17 | -1.1 | 252.4 |
| Cy3dOXG | Cy-3-(dioxaloyl)-glucoside | 593 | 287 | 0.17 | 2.55 | 280.2 |
| Cy3R | Cy-3-rutinoside | 595 | 449/287 | 0.9 | −1.64 | 252.4 |
| Cy3XR | Cy-3-xylosylrutinoside | 727 | 581/287 | 2.52 | −2.1 | 311.3 |
| Cy3GR | Cy-3-glucosylrutinoside | 757 | 287/611 | 3.85 | −2.8 | 331.5 |
| Cy3,5GG | Cy-3,5-diglucoside | 611 | 449/287 | -- | −2.3 | 272.6 |
| Cy3So | Cy-3-sophoroside | 611 | 287 | -- | -- | 260 |
| Cy3Sa5R | Cy-3-sambubioside-5-rhamnoside | 727 | 581/433/287 | -- | -- | -- |
| Cy3So5R | Cy-3-sophoroside-5-rhamnoside | 757 | 611/433/287 | -- | -- | -- |
| Molecule | pH | Epa/mV | Technique | Electrodes | Ref. |
|---|---|---|---|---|---|
| Cy3G | 3.5 | 490, 980 | DPV | WE: Glassy carbon RE: Ag|AgCl | [47] |
| 4.5 | 420, 815 | ||||
| 7.0 | 310, 500 | ||||
| Cy3G | 2.2 | 548 | CV | WE: Glassy carbon RE: Ag|AgCl | [52] |
| 4.8 | 400 | ||||
| 5.9 | 310 | ||||
| 6.9 | 230 | ||||
| Cy3G | 2.0 | 500 | ASSWV | WE: Paraffin rod impregnated RE: Ag|AgCl | [52] |
| Cy | 403 | ||||
| Cy3G | 1.0 | 617 | CV | WE: Platinum RE: SCE | [53] |
| Group | Fruit/Vegetable | Cy3G 2 | Major ACNs 3 |
|---|---|---|---|
| Fruits/Berries | Black elderberry | 794.13 | Cy3G |
| Blackberry raw | 138.72 | Cy3G | |
| Black Aestivalis grape | 18.72 | Cy3G | |
| Gooseberry | 2.95 | Cy3G | |
| Nectarine peeled | 0.56 | Cy3G | |
| Peach peeled | 0.28 | Cy3G | |
| Blackcurrant raw | 25.07 | Dp3R (304.91) | |
| Black chokeberry | 19.64 | Cy3A (252.76) | |
| Blueberry | 14.2 | Dp3G (22.6) | |
| Sweet cherry raw | 18.73 | Cy3R (143.27) | |
| Red raspberry | 14.89 | Cy3So (37.61) | |
| Raspberry pomace (dry) | 41.52 | Cy3So (100.1) | |
| Plum fresh | 8.63 | Cy3R (33.85) | |
| Lowbush blueberry | 7.5 | Mv3G (26.06) | |
| Redcurrant | 3.37 | Cy3XR (11.22) | |
| Strawberry | 2.88 | Pg3G (47.1) | |
| Lingonberry | 1.42 | Cy3Ga (48.69) | |
| Highbush blueberry | 1.37 | Dp3Ga (20.50) | |
| Sour cherry | 1.12 | Cy3GR (43.63) | |
| Black grape | 1.08 | Mv3G (39.23) | |
| American Cranberry | 0.74 | Pn3Ga (22.02) | |
| Cloudberry | 0.62 | Cy3R (1.86) | |
| Juices/wine | Pomegranate pure juice | 3.43 | Cy3G |
| Blood orange pure juice | 1.41 | Cy3MG (1.76) | |
| Red wine | 0.21 | Mv3G (9.97) | |
| Cereals/legumes | Black bean raw | 3.99 | Dp3G (20.50) |
| Blue maize hybrid | 2.25 | Cy3G | |
| Vegetables | Black Olive raw | 10.62 | Cy3R (72.35) |
| Red lettuce raw | 0.62 | Cy3MG (2.91) |
| Cell Line | Cy3G dose | Mechanism | Ref. |
|---|---|---|---|
| Erythrocytes | 10–100 μM | ↓ cholesterol and TBAR in cell membranes | [1] |
| Human adherent macrophages (from U937 cells), oral epithelial cells (GMSM-K) and gingival fibroblasts (HGF-1) | 5–25 μg/mL | ↓ IL-6 level (macrophages), cytoprotection (GMSM-K, HGF-1) against nicotine toxicity | [4] |
| Colon (Caco2), liver (HepG2), prostate (PC3) | Blue maize ACNs (189–500 μg/g)-extract | ↓ cell proliferation | [36] |
| Gastric cancer (KATO III) | 12.5 μM | ↓ Helicobacter pylori VacA-induced cell death | [91] |
| Adipocytes (3T3-L1) | 50 μM | ↓ FoxO1-mediated transcription of lipase | [123] |
| Hepatome (HepG2) | 1–100 μM | ↑ fatty acid oxidation and AMPK activity | [124] |
| Adipocyte | 0.5–50 μM + docosahexanoic acid | ↓ basal lipolysis , inflammatory markers | [127] |
| Breast cancer (BT474m MD-MB231, MCF7) | 10 μM | ↓ invasion / increased expression of ErB2 | [133] |
| Murine thymoma (EL-4T) | 2.5–5.0 μg/mL | ↓ Il-3 & IL-4 by GATA-3 inhibition | [134] |
| Pheochromocytoma (PC-12) | IC50, 15.3 μg/mL | ↓ ATP-induced [Ca2+] increase | [135] |
| Colon cancer (HT-29) | 25 μM | ↓ IL-8, nitrite, PGE2 | [136] |
| human aortic epithelial cells | 0.5–50 μM | ↑ oxiesterol efflux, ↑ABCG1/ABCA1 expression | [137] |
| Heart (isolated mitochondria) | 20 Μm | ↑ phosphorylation, ATP production, ↑e− carrier | [138] |
| Adipocytes (steam cells) | 100 μg/mL | ↓ IL-6 level | [139] |
| Ovarian cancer (HO-8910PM) | IC50, 13.8 μg/mL | ↑ apoptosis, ↓mucin 4 expression | [140] |
| Model | Protocol (Dose) | Effects | Ref. |
|---|---|---|---|
| Mice (nude), SKH-1 | Oral | ↓ lipid per oxidation ↑ Glutathione | [3] |
| Mice, C57BL/6 | Oral, 24 h before, (2 mg/kg) | ↓ Neuronal apoptosis reducing factor, superoxide level, infarct size | [5] |
| Mice (obese), C57BL/6 | Oral, 5 w, 0.02% diet, (ad libitum) | Antidiabetic by modulating c-jun N-terminal kinase | [122] |
| Mice, apoE (-) | Oral, 12 w, 0.06% diet, (ad libitum) | ↓ expression of hepatic cholesterol 7a-hydroxylase | [137] |
| Mice, ovarian cancer | Oral, 2 w, (5 mg/kg) | ↓ Growth of ovarian xenograft tumors | [140] |
| Mice, C57BL/6 | Oral, 12 w, (40–200 mg/kg) | ↓ weigh gain, insulin resistance, adiposity, leptin | [141] |
| Mice, apoE (-) | Oral, 8 w, 0.2% diet (ad libitum) | ↓ atherogenesis, ↑endothelial repair | [142] |
| Mice, with peritonitis/edema | Oral, 30–60 min before (40 mg/kg) | ↓ inflammation, expression COX-2/PGE2 | [143] |
| Mice, diabetic | Oral, 4 w, (300 µg/10g) | ↓ blood glucose | [144] |
| Mice, KK-Ay | Oral | ↓ visceral fat ↑ lipoprotein lipase | [145] |
| Mice, C57BL/6 w/acute alcohol-induced liver injury | Oral, 24–48 h, (10 mg/kg) | ↓ plasma IL-6 ,TNF-α, ALT and AST and ↑ SIRT1 p-c-Jun and Bax expression | [146] |
| Rats (retinal degeneration) | Oral, 5 w, 100 mg/kg | ↓ Loss of photoreceptors | [147] |
| Mice (fetus) | Intra-peritoneal 10–30 mg/kg | ↓Neuronal damage by caspase 3 inhibition | [148] |
| Rats (β-amyloidosis) | Oral, 30 day, 10 mg/kg | ↓ cognitive impairment induced by Aβ via the modulation of GSK-3β/tau | [149] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivas-Aguirre, F.J.; Rodrigo-García, J.; Martínez-Ruiz, N.D.R.; Cárdenas-Robles, A.I.; Mendoza-Díaz, S.O.; Álvarez-Parrilla, E.; González-Aguilar, G.A.; De la Rosa, L.A.; Ramos-Jiménez, A.; Wall-Medrano, A. Cyanidin-3-O-glucoside: Physical-Chemistry, Foodomics and Health Effects. Molecules 2016, 21, 1264. https://doi.org/10.3390/molecules21091264
Olivas-Aguirre FJ, Rodrigo-García J, Martínez-Ruiz NDR, Cárdenas-Robles AI, Mendoza-Díaz SO, Álvarez-Parrilla E, González-Aguilar GA, De la Rosa LA, Ramos-Jiménez A, Wall-Medrano A. Cyanidin-3-O-glucoside: Physical-Chemistry, Foodomics and Health Effects. Molecules. 2016; 21(9):1264. https://doi.org/10.3390/molecules21091264
Chicago/Turabian StyleOlivas-Aguirre, Francisco J., Joaquín Rodrigo-García, Nina Del R. Martínez-Ruiz, Arely I. Cárdenas-Robles, Sandra O. Mendoza-Díaz, Emilio Álvarez-Parrilla, Gustavo A. González-Aguilar, Laura A. De la Rosa, Arnulfo Ramos-Jiménez, and Abraham Wall-Medrano. 2016. "Cyanidin-3-O-glucoside: Physical-Chemistry, Foodomics and Health Effects" Molecules 21, no. 9: 1264. https://doi.org/10.3390/molecules21091264