
Antibacterial Activities and Possible Modes of Action of Acacia nilotica (L.) Del. against Multidrug-Resistant Escherichia coli and Salmonella


Abstract
:1. Introduction
2. Results and Discussion
2.1. Antibiogram of Salmonella and E. coli
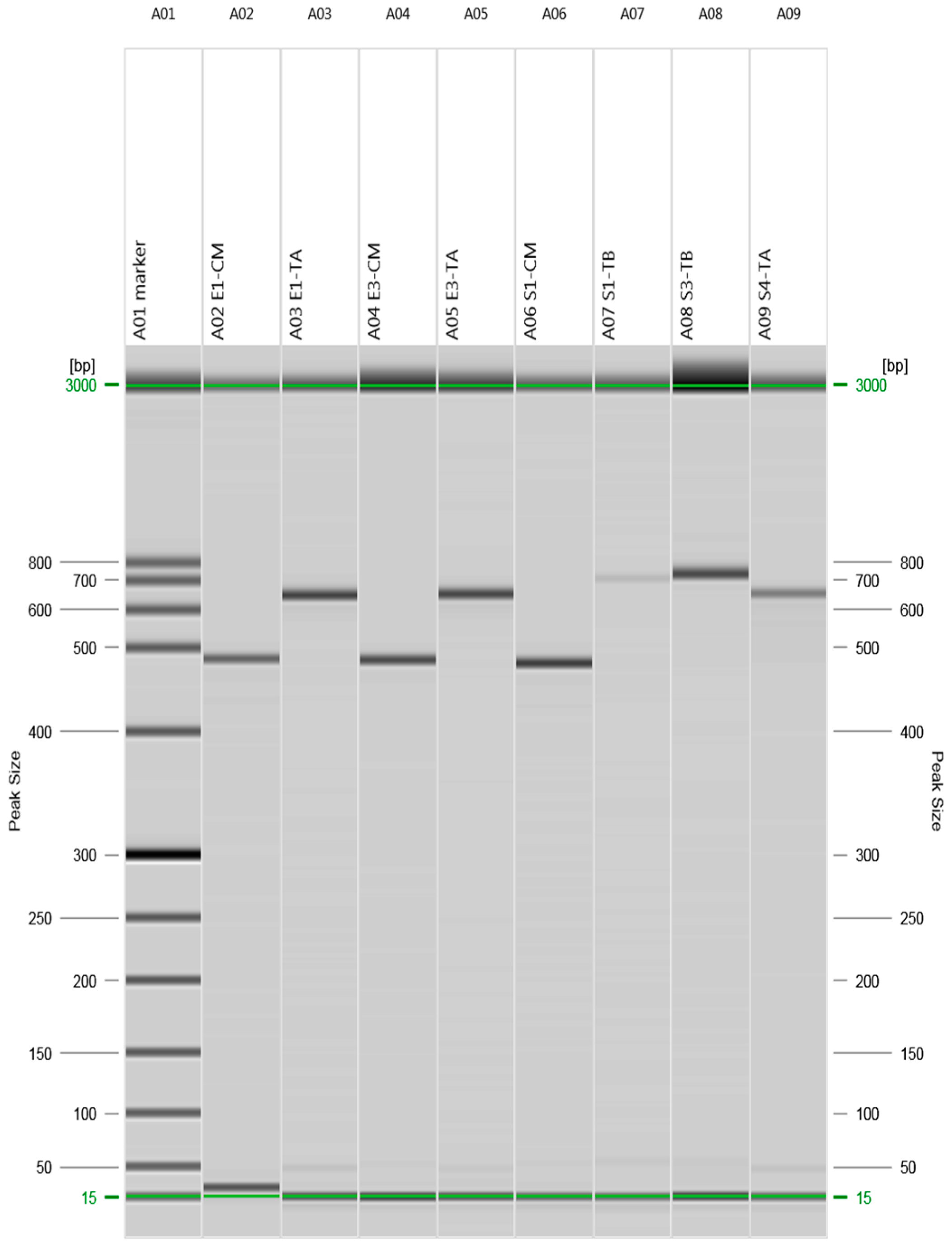
2.2. Detection of Beta-Lactams- and Tetracycline-Resistant Genes
2.3. Antimicrobial Activity of Acacia Extracts
2.4. Minimum Inhibitory Concentration and Minimum Bactericide Concentration of Acacia nilotica Extracts
2.5. Kill-Time Analysis
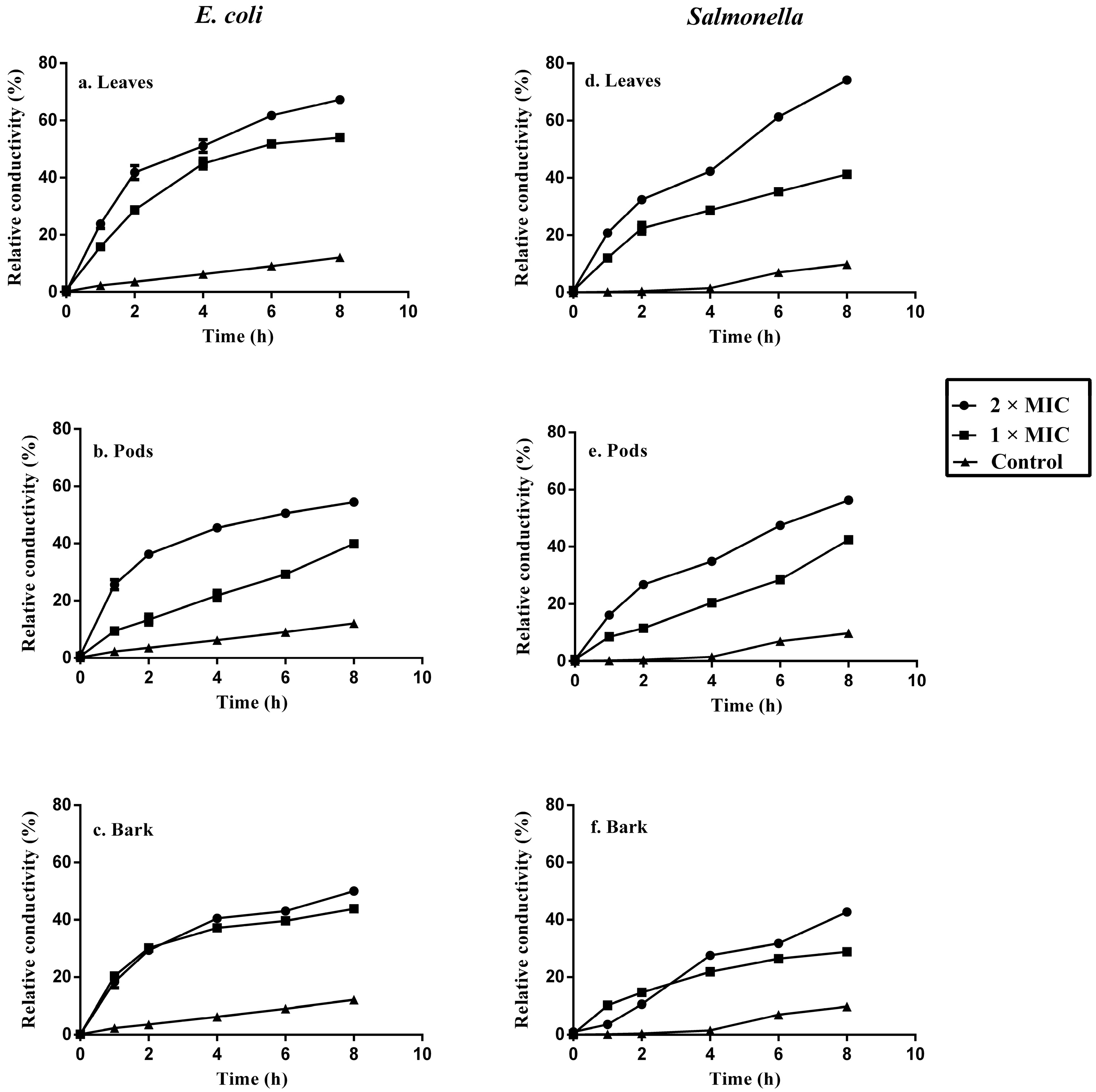
2.6. Bacterial Cell Membrane Permeability after Treatment with Acacia Extracts
2.7. Integrity of Bacterial Cell Membrane
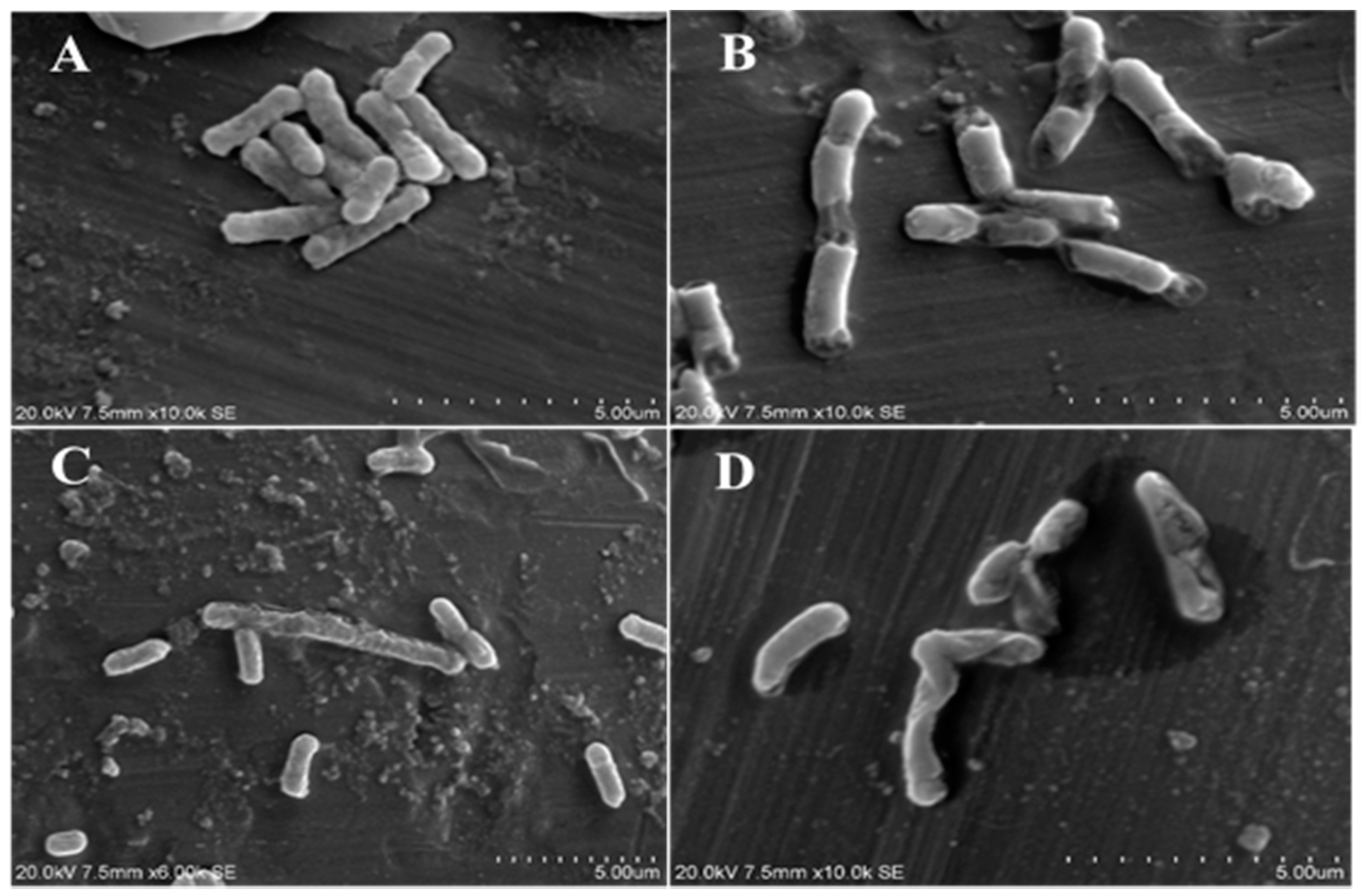
2.8. Scanning Electron Microscope Observations
3. Material and Methods
3.1. Preparation of Plant Extracts and Microbial Sample Collection and Identification
3.2. Antibiogram of Salmonella spp. and E. coli
3.3. Detection of Beta-Lactam- and Tetracycline-Resistant Genes
3.4. Antimicrobial Activity of Acacia Extracts
3.5. Determination of Minimum Inhibitory Concentration and Minimum Bactericide Concentration
3.6. Kill-Time Analysis
3.7. Bacterial Cell Membrane Permeability
3.8. Integrity of Bacterial Cell Membrane
3.9. Scanning Electron Microscope Observations
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harbottle, H.; Thakur, S.; Zhao, S.; White, D.G. Genetics of antimicrobial resistance. Anim. Biotechnol. 2006, 17, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Fernández, E.; Alonso-Calleja, C.; García-Fernández, C.; Capita, R. Prevalence and antimicrobial resistance of Salmonella serotypes isolated from poultry in Spain: Comparison between 1993 and 2006. Int. J. Food Microbiol. 2012, 153, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.F.; Paixao, T.A.; Tsolis, R.M.; Baeumler, A.J.; Santos, R.L. Salmonellosis in cattle: Advantages of being an experimental model. Res. Vet. Sci. 2012, 93, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Petri, W.A.; Miller, M.; Binder, H.J.; Levine, M.M.; Dillingham, R.; Guerrant, R.L. Enteric infections, diarrhea, and their impact on function and development. J. Clin. Investig. 2008, 118, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Anal, A.K. Prevalence and antibiogram study of Salmonella and Staphylococcus aureus in poultry meat. Asian Pac. J. Trop. Biomed. 2013, 3, 163–168. [Google Scholar] [CrossRef]
- Koga, V.L.; Rodrigues, G.R.; Scandorieiro, S.; Vespero, E.C.; Oba, A.; de Brito, B.G.; Kobayashi, R.K. Evaluation of the antibiotic resistance and virulence of Escherichia coli strains isolated from chicken carcasses in 2007 and 2013 from Paraná, Brazil. Foodborne Pathog. Dis. 2015, 12, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Anal, A.K. Isolation of Salmonella from ready-to-eat poultry meat and evaluation of its survival at low temperature, microwaving and simulated gastric fluids. J. Food Sci. Technol. 2014, 52, 3051–3057. [Google Scholar] [CrossRef] [PubMed]
- Coates, A.; Hu, Y.; Bax, R.; Page, C. The future challenges facing the development of new antimicrobial drugs. Nat. Rev. Drug. Discov. 2002, 1, 895–910. [Google Scholar] [CrossRef] [PubMed]
- Braga, L.C.; Leite, A.A.; Xavier, K.G.; Takahashi, J.A.; Bemquerer, M.P.; Chartone-Souza, E.; Nascimento, A.M. Synergic interaction between pomegranate extract and antibiotics against Staphylococcus aureus. Can. J. Microbiol. 2005, 51, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Akinmoladun, A.C.; Ibukun, E.; Afor, E.; Akinrinlola, B.; Onibon, T.; Akinboboye, A. Chemical constituents and antioxidant activity of Alstonia boonei. Afr. J. Biotechnol. 2007, 6, 1197–1201. [Google Scholar]
- Ambasta, S.P. The Useful Plants of India: National Institute of Science Communication; Council of Scientific & Industrial Research: New Delhi, India, 1992. [Google Scholar]
- Oladosu, P.; Isu, N.R.; Ibrahim, K.; Okolo, P.; Oladepo, D.K. Time kill-kinetics antibacterial study of Acacia nilotica. Afr. J. Microbiol. Res. 2013, 7, 5248–5252. [Google Scholar] [CrossRef]
- Saini, M.L.; Saini, R.; Roy, S.; Kumar, A. Comparative pharmacognostical and antimicrobial studies of acacia species (Mimosaceae). J. Med. Plant Res. 2008, 2, 378–386. [Google Scholar]
- Deshpande, S.N. Preliminary phytochemical analysis and in vitro investigation of antibacterial activity of Acacia nilotica against clinical isolates. J. Pharmacogn. Phytochem. 2013, 1, 23–27. [Google Scholar]
- Addis, Z.; Kebede, N.; Sisay, Z.; Alemayehu, H.; Wubetie, A.; Kassa, T. Prevalence and antimicrobial resistance of Salmonella isolated from lactating cows and in contact humans in dairy farms of Addis Ababa: A cross sectional study. BMC Infect. Dis. 2011, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.E.; van Asselt, E.D.; Zwietering, M.H.; Nauta, M.J.; de Jonge, R. Extreme heat resistance of food borne pathogens Campylobacter jejuni, Escherichia coli, and Salmonella Typhimurium on chicken breast fillet during cooking. Int. J. Microbiol. 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Skočková, A.; Koláčková, I.; Bogdanovičová, K.; Karpíšková, R. Characteristic and antimicrobial resistance in Escherichia coli from retail meats purchased in the Czech Republic. Food Control 2015, 47, 401–406. [Google Scholar] [CrossRef]
- Kavitha, P.A.; Kumar, P.; Murthy, T.P.N.; Gopinath, S.M. Methanolic extract of Acacia nilotica and antibacterial activity against Hospital isolates of Bengaluru district. Int. J. Latest Res. Sci. Technol. 2013, 2, 522–524. [Google Scholar]
- Khan, R.; Islam, B.; Akram, M.; Shakil, S.; Ahmad, A.A.; Ali, S.M.; Khan, A.U. Antimicrobial activity of five herbal extracts against multi drug resistant (MDR) strains of bacteria and fungus of clinical origin. Molecules 2009, 14, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Okoro, S.O.; Kawo, A.H.; Arzai, A.H. Phytochemical Screening, Antibacterial and Toxicological Activities of Acacia nilotica Extracts. Bayero J. Pure Appl. Sci. 2014, 7, 105–115. [Google Scholar] [CrossRef]
- Sharma, C.; Aneja, K.R.; Surain, P.; Dhiman, R.; Jiloha, P.; Meashi, V.; Kaur, M. In vitro evaluation of anti-microbial spectrum of Acacia nilotica leaves and bark extracts against pathogens causing otitis infection. J. Innov. Biol. 2014, 1, 051–056. [Google Scholar]
- National Committee for Clinical Laboratory Standards. Methods for Determining Bactericidal Activity of Antimicrobial Agents. Tentative Guidelines; National Committee for Clinical Laboratory Standards Document M26-T; NCCLS: Villanova, PA, USA, 1992; Volume 12, No. 19. [Google Scholar]
- Yadav, A.; Yadav, M.; Kumar, S.; Yadav, J.P. Bactericidal effect of Acacia nilotica: In Vitro antibacterial and time kill kinetic studies. Int. J. Curr. Res. 2015, 7, 22289–22294. [Google Scholar]
- Cox, S.D.; Mann, C.M.; Markham, J.L.; Gustafson, J.E.; Warmington, J.R.; Wyllie, S.G. Determining the antimicrobial actions of tea tree oil. Molecules 2001, 6, 87–91. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, M.; Zhao, Z.; Yu, S. The antibiotic activity and mechanisms of sugarcane (Saccharum officinarum L.) bagasse extract against food-borne pathogens. Food Chem. 2015, 185, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Diao, W.R.; Hu, Q.P.; Zhang, H.; Xu, J.G. Chemical composition, antibacterial activity and mechanism of action of essential oil from seeds of fennel (Foeniculum vulgare Mill.). Food Control 2014, 35, 109–116. [Google Scholar]
- Bajpai, V.K.; Sharma, A.; Baek, K.H. Antibacterial mode of action of Cudrania tricuspidata fruit essential oil, affecting membrane permeability and surface characteristics of food-borne pathogens. Food Control 2013, 32, 582–590. [Google Scholar] [CrossRef]
- Sadiq, M.B.; Hanpithakpong, W.; Tarning, J.; Anal, A.K. Screening of phytochemicals and in vitro evaluation of antibacterial and antioxidant activities of leaves, pods and bark extracts of Acacia nilotica (L.) Del. Ind. Crops Prod. 2015, 77, 873–882. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Adwan, G.; Abu-Shanab, B.; Adwan, K. Antibacterial activities of some plant extracts alone and in combination with different antimicrobials against multidrug-resistant Pseudomonas aeruginosa strains. Asian Pac. J. Trop. Med. 2010, 3, 266–269. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty Third Informational Supplement; CLSI Documents: Wayne, PA, USA, 2013. [Google Scholar]
- Van, T.T.H.; Chin, J.; Chapman, T.; Tran, L.T.; Coloe, P.J. Safety of raw meat and shellfish in Vietnam: An analysis of Escherichia coli isolations for antibiotic resistance and virulence genes. Int. J. Food Microb. 2008, 124, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.P.; Cooles, S.W.; Osborn, M.K.; Piddock, L.J.V.; Woodward, M.J. Antibiotic resistance genes, integrons and multiple antibiotic resistance in thirty-five serotypes of Salmonella enterica isolated from humans and animals in the UK. J. Antimicrob. Chemother. 2004, 53, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Duraipandiyan, V.; Ayyanar, M.; Ignacimuthu, S. Antimicrobial activity of some ethnomedicinal plants used by Paliyar tribe from Tamil Nadu, India. BMC Complement. Altern Med. 2006, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Fujita, K.I.; Kubo, A.; Nihei, K.I.; Ogura, T. Antibacterial activity of coriander volatile compounds against Salmonella choleraesuis. J. Agric. Food Chem. 2004, 52, 3329–3332. [Google Scholar] [CrossRef] [PubMed]
- Joray, M.B.; del Rollán, M.R.; Ruiz, G.M.; Palacios, S.M.; Carpinella, M.C. Antibacterial activity of extracts from plants of central Argentina—Isolation of an active principle from Achyrocline satureioides. Planta Med. 2011, 77, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Chen, X.G.; Liu, C.S.; Liu, C.G.; Meng, X.H.; Yu, L.J. Antibacterial mechanism of chitosan microspheres in a solid dispersing system against E. coli. Colloids Surf. B Biointerfaces 2008, 65, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Sun, C.; Liang, Z.; Han, Y.; Yu, J. Antibacterial activity of hypocrellin A against Staphylococcus aureus. World J. Microbiol. Biotechnol. 2012, 28, 3151–3157. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Resistance | Susceptible |
|---|---|---|
| E1 | Amp, Aml, Chl, Gen, Tet, Cip, Ctx | Amk, Str, Sxt |
| E2 | Amp, Aml, Chl, Gen, Amk, Str, Tet, Sxt, Cip, Ctx | |
| E3 | Amp, Aml, Chl, Tet | Gen, Amk, Str, Sxt, Cip, Ctx |
| S1 | Amp, Aml, Str, Tet | Chl, Gen, Amk, Sxt, Cip, Ctx |
| S2 | Amp, Aml, Chl, Gen, Amk, Str, Tet, Sxt, Cip, Ctx | |
| S3 | Tet, Chl | Amp, Aml, Gen, Amk, Str, Sxt, Cip, Ctx |
| S4 | Amp, Aml, Tet | Gen, Amk, Str, Cip, Ctx, Chl, Sxt |
| Sample | Conc. (mg/disc) | Zone of Inhibition (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| E1 | E2 | E3 | Mean of E. coli Strains | S1 | S2 | S3 | S4 | Mean of Salmonella Strains | ||
| Leaves | 5 | 17.0 ± 1.0 | 16.7 ± 1.15 | 16.7 ± 0.58 | 16.78 ± 0.83 a | 12.3 ± 0.58 | 11.3 ± 1.53 | 11.3 ± 2.08 | 11.7 ± 1.15 | 11.67 ± 1.30 a |
| 10 | 21.7 ± 0.6 | 21.0 ± 1.0 | 20.7 ± 1.53 | 21.11 ± 1.05 b | 17.7 ± 0.58 | 16.7 ± 0.58 | 16.0 ± 1.0 | 17.0 ± 1.0 | 16.83 ± 0.94 b | |
| Pods | 5 | 9.3 ± 0.58 | 8.3 ± 0.58 | 7.7 ± 0.58 | 8.44 ± 0.88 | 8.3 ± 0.58 | 8.7 ± 1.15 | 7.7 ± 1.15 | 8.3 ± 0.58 | 8.25 ± 0.86 |
| 10 | 19.0 ± 1.0 | 18.3 ± 0.58 | 15.3 ± 0.58 | 17.56 ± 1.81 b | 15.7 ± 1.53 | 13.7 ± 0.58 | 14.7 ± 1.15 | 15.0 ± 1.0 | 14.75 ± 1.21 b | |
| Bark | 5 | 8.7 ± 0.58 | 9.0 ± 1.0 | 8.3 ± 0.58 | 8.67 ± 0.71 | 5.3 ± 4.61 | 8.7 ± 0.58 | 8.7 ± 1.15 | 8.0 ± 1.0 | 7.67 ± 2.53 |
| 10 | 13.0 ± 1.0 | 12.3 ± 0.58 | 11.3 ± 1.53 | 12.22 ± 1.20 b | 10.3 ± 1.15 | 11.3 ± 1.15 | 11.7 ± 0.58 | 11.0 ± 2.0 | 11.08 ± 1.24 b | |
| Amikacin (Control) | 30 µg | 22.4 ± 1.51 | 23.8 ± 1.73 | 22.5 ± 1.1 | 22.8 ± 1.43 | 23.4 ± 2.14 | 23.5 ± 1.53 | 22.0 ± 1.0 | 23.0 ± 1.0 | 22.97 ± 1.24 |
| Microbial Strains | Minimum Inhibitory Concentration | Minimum Bactericide Concentration | ||||
|---|---|---|---|---|---|---|
| Leaves | Pods | Bark | Leaves | Pods | Bark | |
| (mg/mL) | (mg/mL) | (mg/mL) | (mg/mL) | (mg/mL) | (mg/mL) | |
| E1 | 3.12 | 3.12 | 6.25 | 6.25 | 12.5 | 12.5 |
| E2 | 3.12 | 6.25 | 6.25 | 3.12 | 12.5 | 12.5 |
| E3 | 3.12 | 3.12 | 6.25 | 6.25 | 12.5 | 12.5 |
| S1 | 1.56 | 3.12 | 3.12 | 3.12 | 6.25 | 6.25 |
| S2 | 1.56 | 3.12 | 3.12 | 3.12 | 6.25 | 6.25 |
| S3 | 1.56 | 3.12 | 3.12 | 3.12 | 6.25 | 12.5 |
| S4 | 1.56 | 3.12 | 3.12 | 3.12 | 6.25 | 6.25 |
| Sample | Conc. | Cell Constituents Release | |||
|---|---|---|---|---|---|
| E. coli | Salmonella typhimurium | ||||
| Protein (µg/mL) | Cell Constituents (OD260nm) | Protein (µg/mL) | Cell Constituents (OD260nm) | ||
| Acacia leaves | 2 × MIC | 62.70 ± 4.20 | 0.42 ± 0.007 | 48.26 ± 5.25 | 0.4 ± 0.006 |
| 1 × MIC | 32.33 ± 4.00 | 0.34 ± 0.007 | 25.67 ± 4.84 | 0.31 ± 0.001 | |
| Control | 8.63 ± 2.31 | 0.086 ± 0.013 | 10.11 ± 2.22 | 0.09 ± 0.004 | |
| Acacia pods | 2 × MIC | 30.85 ± 2.80 | 0.31 ± 0.029 | 18.63 ± 2.31 | 0.27 ± 0.01 |
| 1 × MIC | 17.52 ± 2.31 | 0.23 ± 0.036 | 12.33 ± 2.22 | 0.164 ± 0.025 | |
| Control | 6.78 ± 2.94 | 0.102 ± 0.006 | 5.67 ± 2.94 | 0.1 ± 0.017 | |
| Acacia bark | 2 × MIC | 39.74 ± 4.62 | 0.30 ± 0.013 | 31.96 ± 3.90 | 0.29 ± 0.006 |
| 1 × MIC | 22.33 ± 2.94 | 0.18 ± 0.016 | 18.67 ± 1.64 | 0.14 ± 0.009 | |
| Control | 5.67 ± 2.22 | 0.088 ± 0.011 | 7.18 ± 1.65 | 0.1 ± 0.01 | |
| Antimicrobial Agent | Resistant Gene | Sequence | Size (bp) | Annealing Temp (°C) | Reference |
|---|---|---|---|---|---|
| Beta-Lactams | bla CMY F | TGGCCAGAACTGACAGGCAAA | 462 | 65.2 | [33] |
| bla CMY R | TTTCTCCTGAACGTGGCTGGC | ||||
| bla SHV F | TCGCCTGTGTATTATCTCCC | 768 | 57.2 | [33] | |
| bla SHV R | CGCAGATAAATCACCACAATG | ||||
| Tetracycline | tet (A) F | GGTTCACTCGAACGACGTCA | 577 | 61.1 | [34] |
| tet (A) R | CTGTCCGACAAGTTGCATGA | ||||
| tet (B) F | CCTCAGCTTCTCAACGCGTG | 634 | 60.3 | [34] | |
| tet (B) R | GCACCTTGCTGATGACTCTT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadiq, M.B.; Tarning, J.; Aye Cho, T.Z.; Anal, A.K. Antibacterial Activities and Possible Modes of Action of Acacia nilotica (L.) Del. against Multidrug-Resistant Escherichia coli and Salmonella. Molecules 2017, 22, 47. https://doi.org/10.3390/molecules22010047
Sadiq MB, Tarning J, Aye Cho TZ, Anal AK. Antibacterial Activities and Possible Modes of Action of Acacia nilotica (L.) Del. against Multidrug-Resistant Escherichia coli and Salmonella. Molecules. 2017; 22(1):47. https://doi.org/10.3390/molecules22010047
Chicago/Turabian StyleSadiq, Muhammad Bilal, Joel Tarning, Tay Zar Aye Cho, and Anil Kumar Anal. 2017. "Antibacterial Activities and Possible Modes of Action of Acacia nilotica (L.) Del. against Multidrug-Resistant Escherichia coli and Salmonella" Molecules 22, no. 1: 47. https://doi.org/10.3390/molecules22010047