
Low-Dose Endotoxin Induces Late Preconditioning, Increases Peroxynitrite Formation, and Activates STAT3 in the Rat Heart

 , ,
, ,
Abstract
:1. Introduction
2. Results
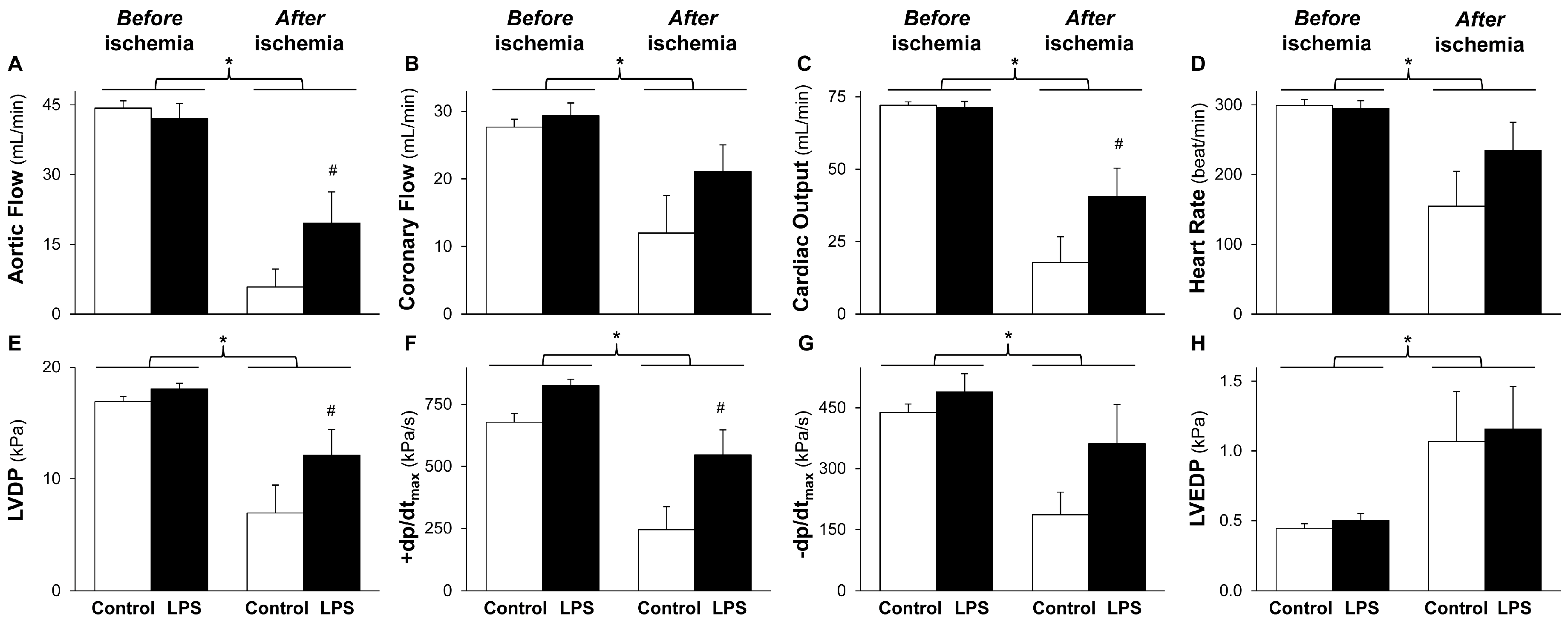
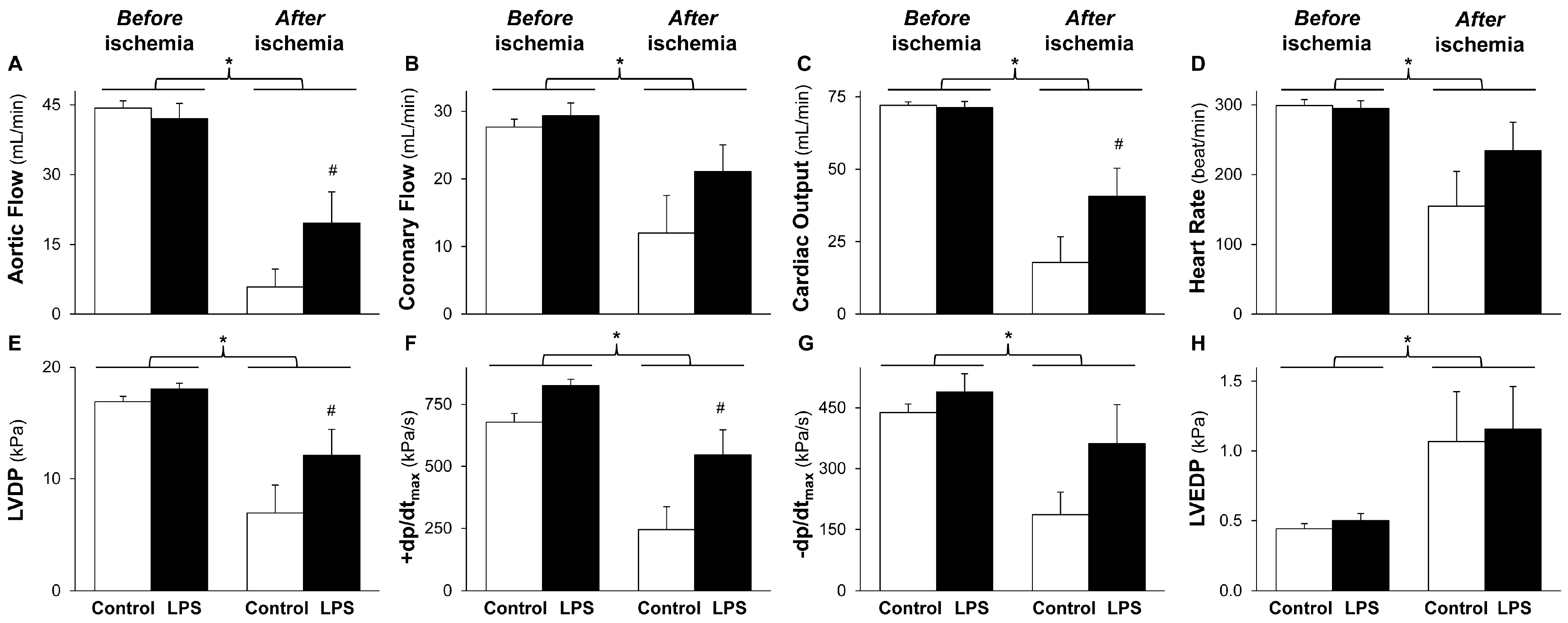
2.1. LPS Pretreatment Improves Post-Ischemic Cardiac Function and LDH Release: Evidence for Delayed Cardioprotection
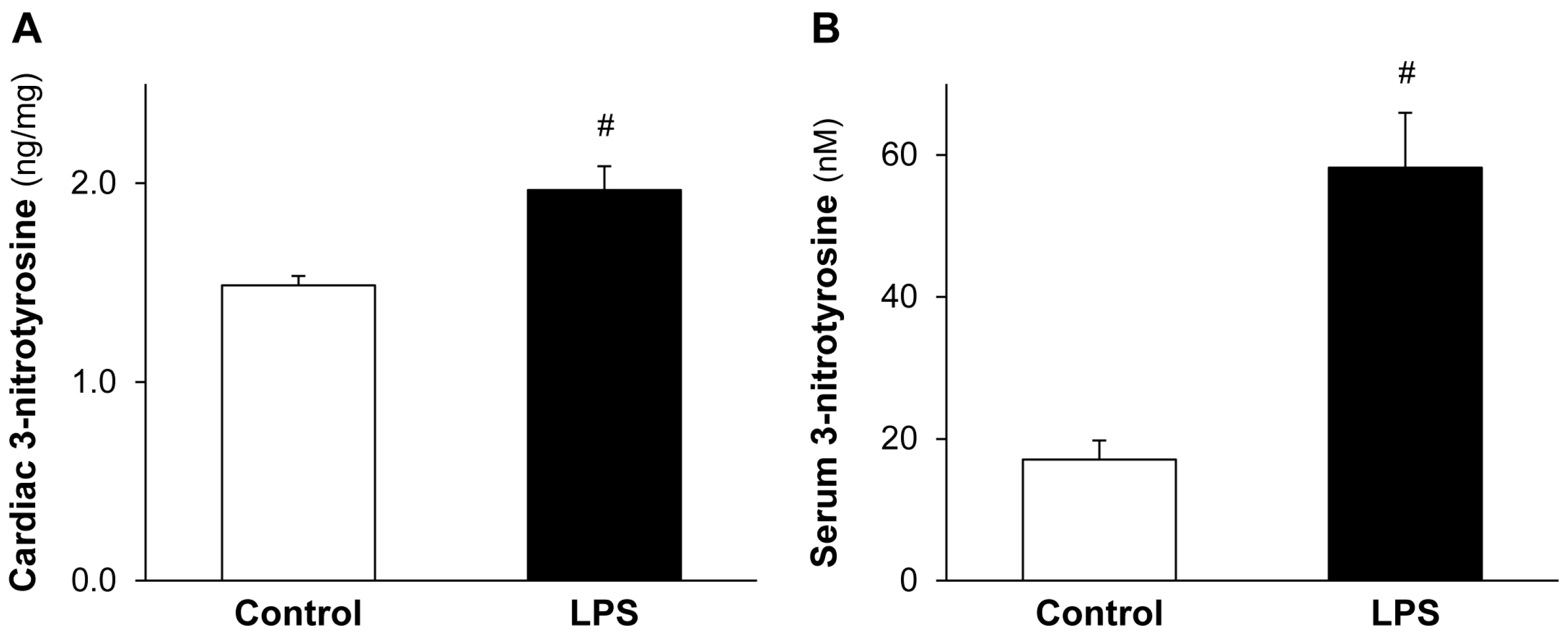
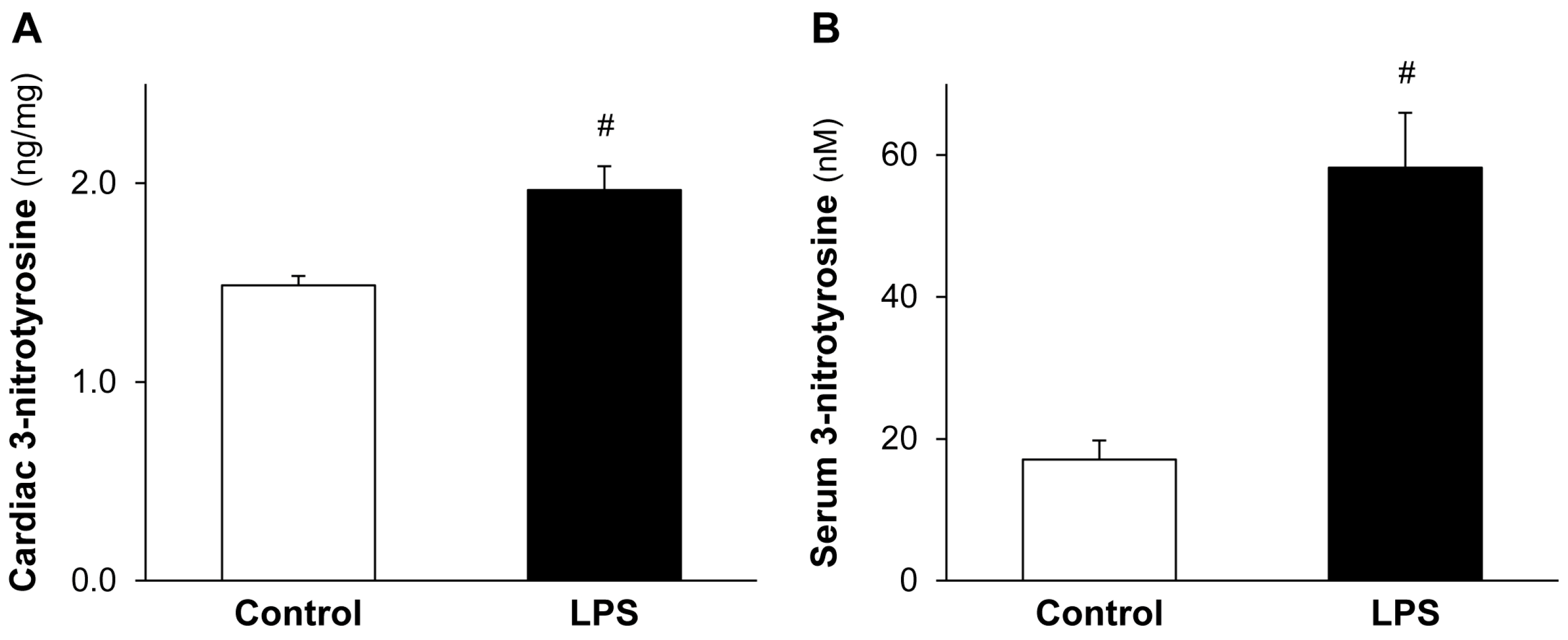
2.2. LPS Pretreatment Enhances Cardiac 3-Nitrotyrosine Formation
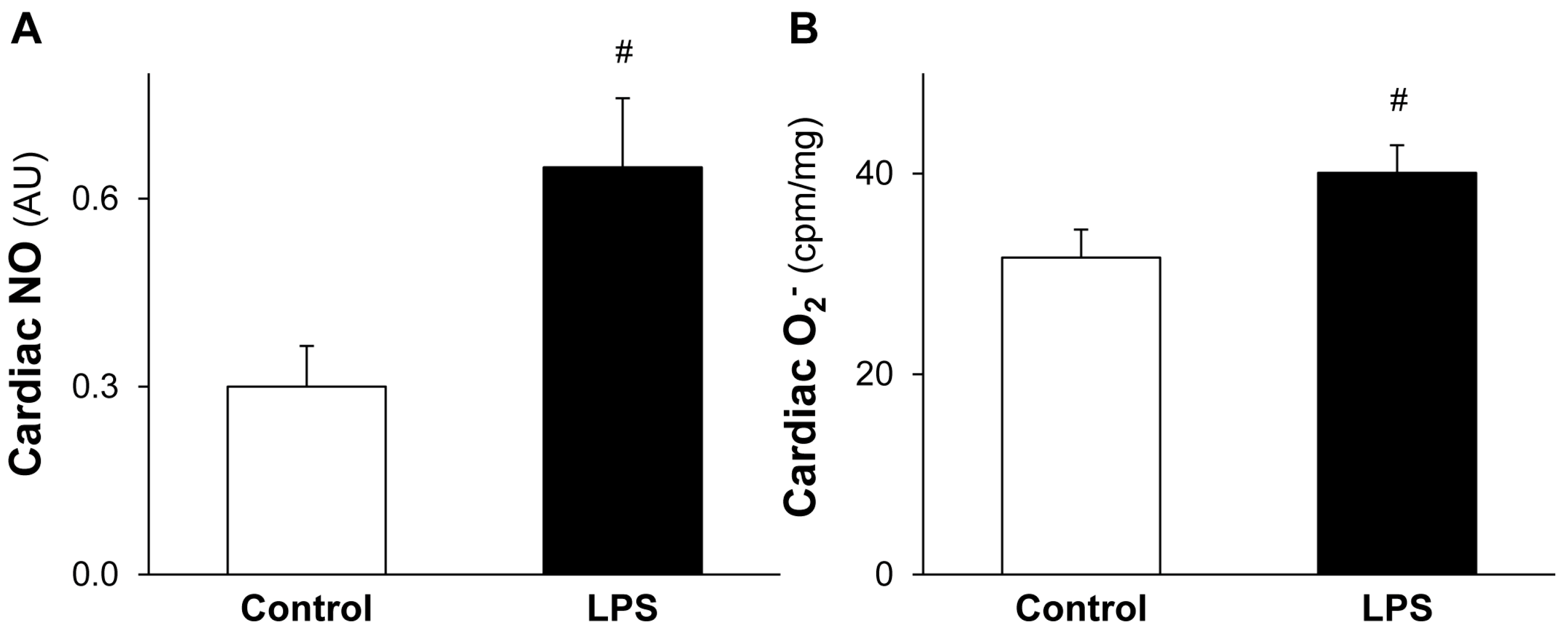
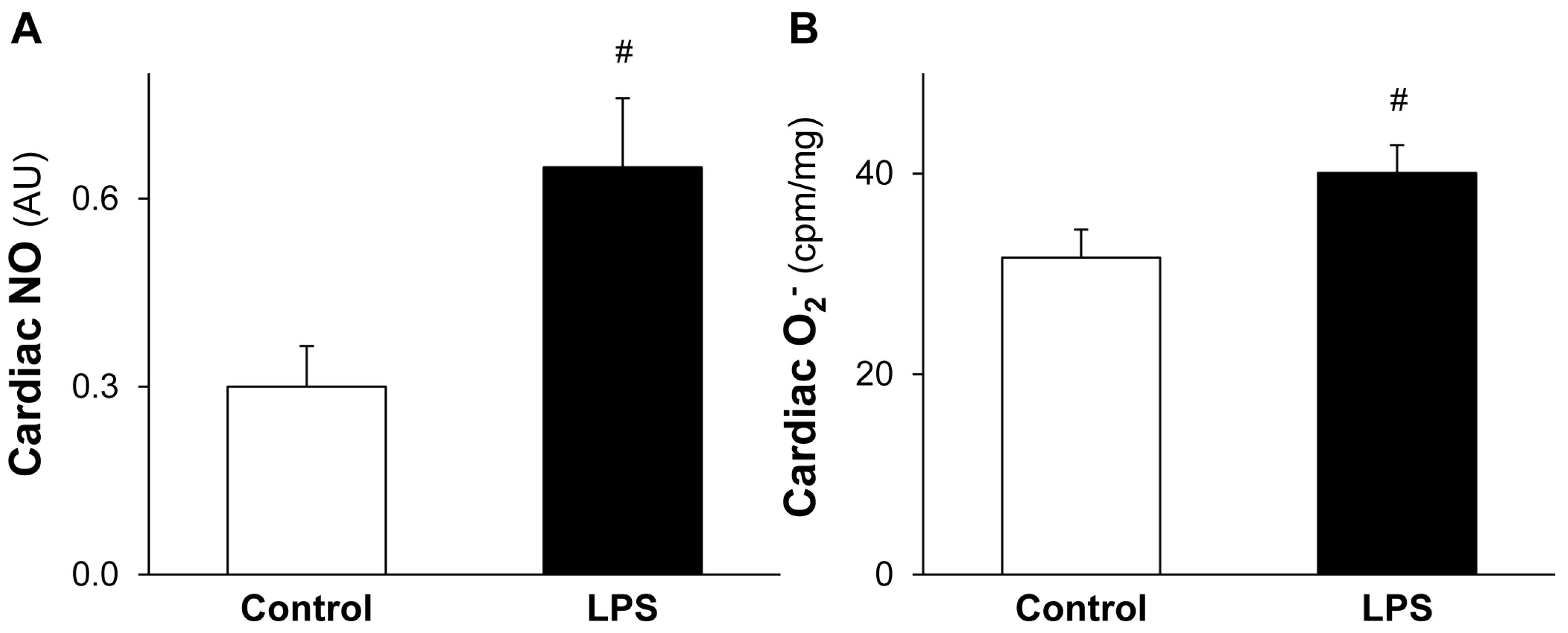
2.3. LPS Pretreatment Leads to Increased Level of Cardiac NO and O2•−
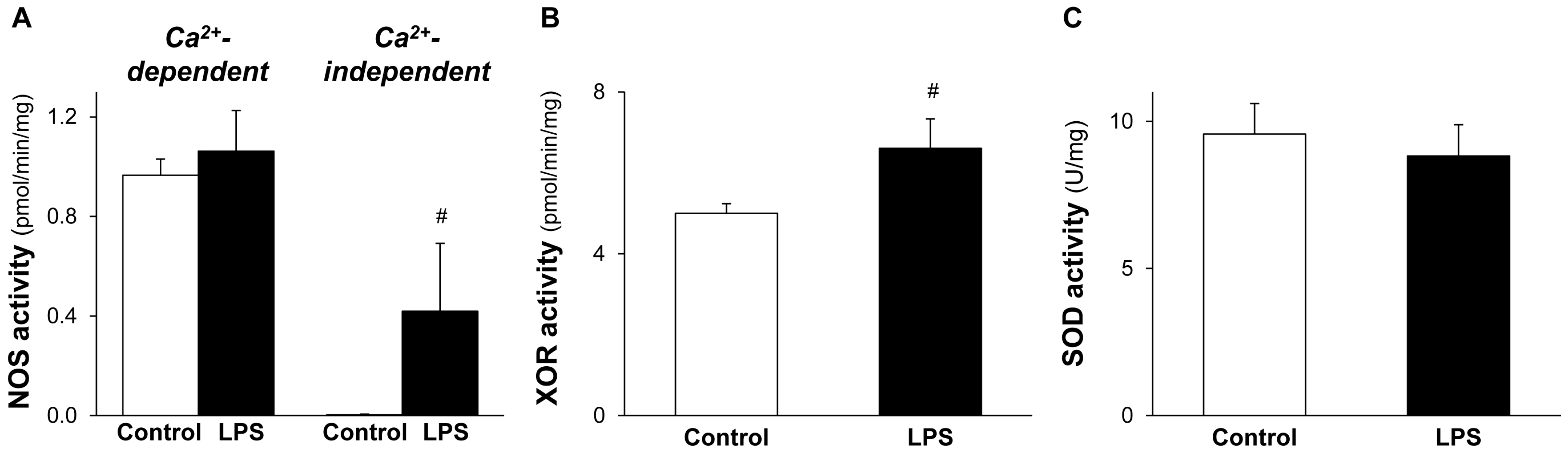
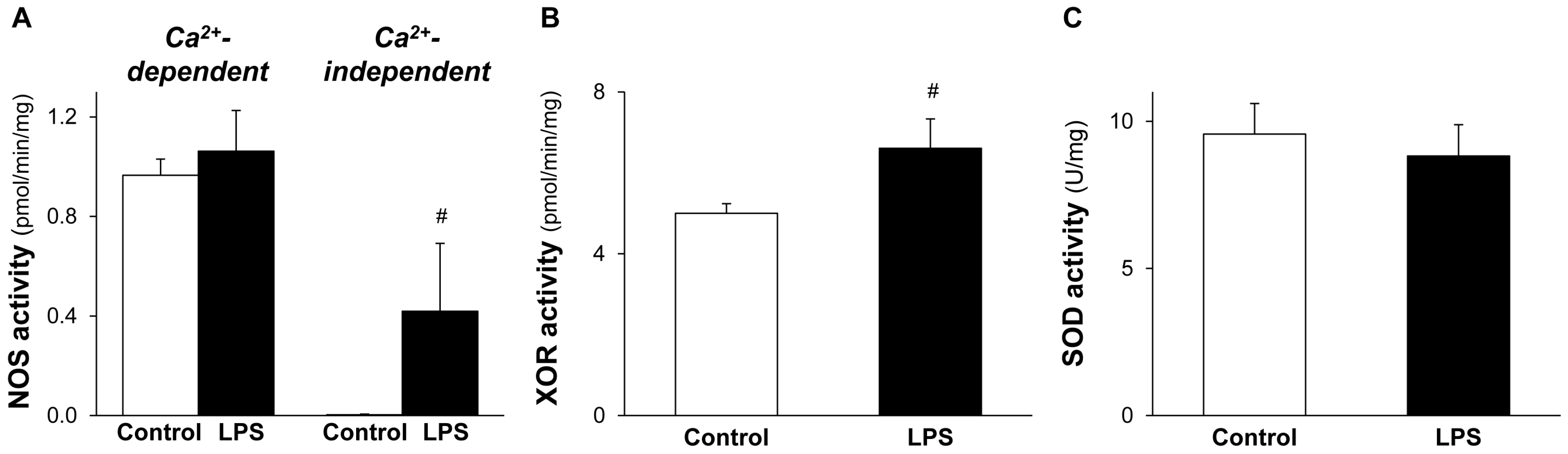
2.4. NOS and XOR Enzymes Contribute to Elevated NO and O2•− Production Induced by LPS
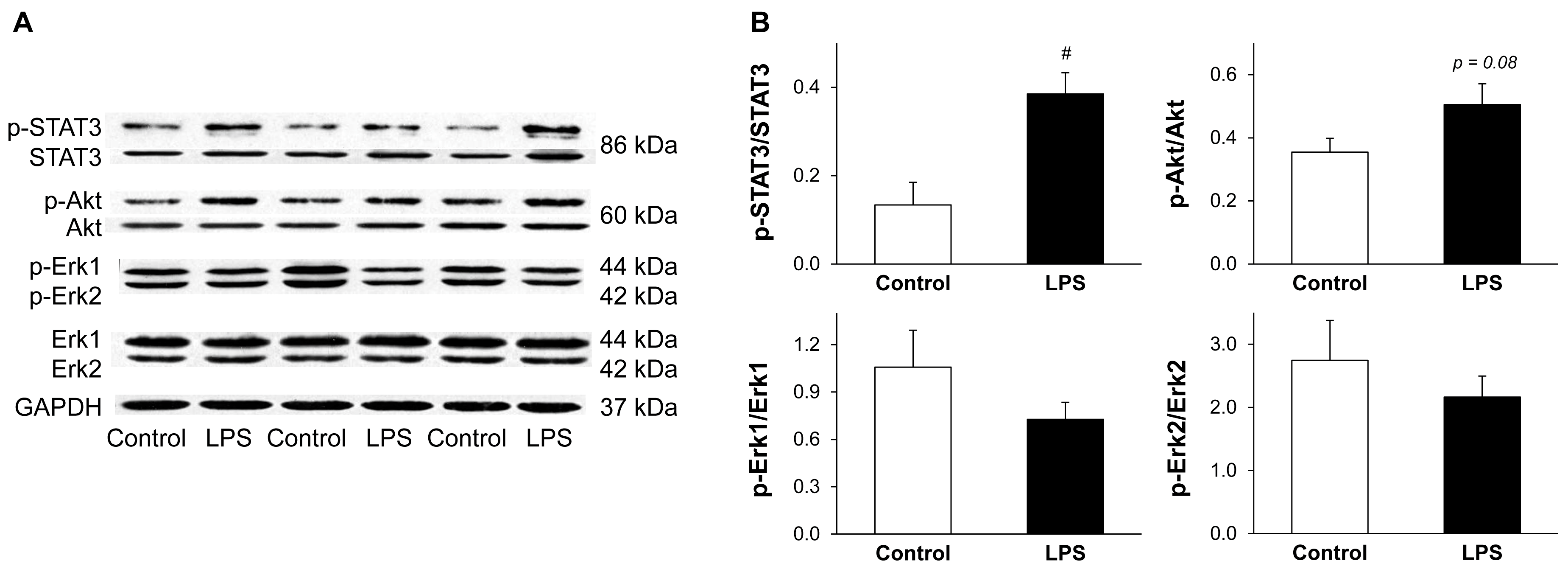
2.5. LPS Pretreatment Results in Enhanced Phosphorylation of STAT3
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Experimental Design and Isolated Heart Perfusion
4.3. Assessment of Cardiac and Serum ONOO− Formation
4.4. Measurement of Cardiac NO and O2•− Levels
4.5. Measurement of Cardiac NO Synthases (NOS), Xanthine Oxidoreductase (XOR), and SOD Activities
4.6. Investigation of Akt, Erk, and STAT3 Activation with Western Blot
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Yellon, D.M.; Downey, J.M. Preconditioning the myocardium: From cellular physiology to clinical cardiology. Physiol. Rev. 2003, 83, 1113–1151. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.B.; Tang, X.L.; Guo, Y.; Xuan, Y.T.; Dawn, B.; Bolli, R. Delayed adaptation of the heart to stress: Late preconditioning. Stroke 2004, 35, 2676–2679. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R. The late phase of preconditioning. Circ. Res. 2000, 87, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Baxter, G.F.; Ferdinandy, P. Delayed preconditioning of myocardium: Current perspectives. Basic Res. Cardiol. 2001, 96, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Grosso, M.A.; Terada, L.S.; Whitman, G.J.; Banerjee, A.; White, C.W.; Harken, A.H.; Repine, J.E. Endotoxin pretreatment increases endogenous myocardial catalase activity and decreases ischemia-reperfusion injury of isolated rat hearts. Proc. Natl. Acad. Sci. USA 1989, 86, 2516–2520. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Sato, C.; Mizoguchi, K.; Yamashita, Y.; Oe, M.; Maeta, H. Lipopolysaccharide triggers late preconditioning against myocardial infarction via inducible nitric oxide synthase. Cardiovasc. Res. 2002, 56, 33–42. [Google Scholar] [CrossRef]
- Wang, E.; Feng, Y.; Zhang, M.; Zou, L.; Li, Y.; Buys, E.S.; Huang, P.; Brouckaert, P.; Chao, W. Toll-like receptor 4 signaling confers cardiac protection against ischemic injury via inducible nitric oxide synthase- and soluble guanylate cyclase-dependent mechanisms. Anesthesiology 2011, 114, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Portnychenko, A.G.; Harmatina, O.Y.; Kotsuruba, A.V.; Moybenko, O.O. The role of nitric oxide in endotoxin-induced cardiodepression. Exp. Clin. Cardiol. 2005, 10, 223–228. [Google Scholar] [PubMed]
- Maulik, N.; Watanabe, M.; Engelman, D.; Engelman, R.M.; Kagan, V.E.; Kisin, E.; Tyurin, V.; Cordis, G.A.; Das, D.K. Myocardial adaptation to ischemia by oxidative stress induced by endotoxin. Am. J. Physiol. 1995, 269, C907–C916. [Google Scholar] [PubMed]
- Csonka, C.; Csont, T.; Onody, A.; Ferdinandy, P. Preconditioning decreases ischemia/reperfusion-induced peroxynitrite formation. Biochem. Biophys. Res. Commun. 2001, 285, 1217–1219. [Google Scholar] [CrossRef] [PubMed]
- Altug, S.; Demiryurek, A.T.; Kane, K.A.; Kanzik, I. Evidence for the involvement of peroxynitrite in ischaemic preconditioning in rat isolated hearts. Br. J. Pharmacol. 2000, 130, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.L.; Takano, H.; Rizvi, A.; Turrens, J.F.; Qiu, Y.; Wu, W.J.; Zhang, Q.; Bolli, R. Oxidant species trigger late preconditioning against myocardial stunning in conscious rabbits. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H281–H291. [Google Scholar] [PubMed]
- Heusch, G.; Boengler, K.; Schulz, R. Cardioprotection: Nitric oxide, protein kinases, and mitochondria. Circulation 2008, 118, 1915–1919. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S. Activation of the protective survivor activating factor enhancement (safe) pathway against reperfusion injury: Does it go beyond the risk pathway? J. Mol. Cell. Cardiol. 2009, 47, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Das, D.K. Molecular mechanism of preconditioning. IUBMB Life 2008, 60, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.; Hua, F.; Liu, X.; Ma, J.; McMullen, J.R.; Shioi, T.; Izumo, S.; Kelley, J.; Gao, X.; Browder, W.; et al. Lipopolysaccharide-induced myocardial protection against ischaemia/reperfusion injury is mediated through a pi3k/akt-dependent mechanism. Cardiovasc. Res. 2008, 78, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Eisen, A.; Fisman, E.Z.; Rubenfire, M.; Freimark, D.; McKechnie, R.; Tenenbaum, A.; Motro, M.; Adler, Y. Ischemic preconditioning: Nearly two decades of research. A comprehensive review. Atherosclerosis 2004, 172, 201–210. [Google Scholar] [CrossRef]
- Pipicz, M.; Varga, Z.V.; Kupai, K.; Gaspar, R.; Kocsis, G.F.; Csonka, C.; Csont, T. Rapid ventricular pacing-induced postconditioning attenuates reperfusion injury: Effects on peroxynitrite, risk and safe pathways. Br. J. Pharmacol. 2015, 172, 3472–3483. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Loukili, N.; Rosenblatt-Velin, N.; Pacher, P.; Feihl, F.; Waeber, B.; Liaudet, L. Peroxynitrite is a key mediator of the cardioprotection afforded by ischemic postconditioning in vivo. PLoS ONE 2013, 8, e70331. [Google Scholar]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Lefer, D.J.; Scalia, R.; Campbell, B.; Nossuli, T.; Hayward, R.; Salamon, M.; Grayson, J.; Lefer, A.M. Peroxynitrite inhibits leukocyte-endothelial cell interactions and protects against ischemia-reperfusion injury in rats. J. Clin. Invest. 1997, 99, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Kawano, T.; Kunz, A.; Abe, T.; Girouard, H.; Anrather, J.; Zhou, P.; Iadecola, C. Inos-derived no and nox2-derived superoxide confer tolerance to excitotoxic brain injury through peroxynitrite. J. Cereb. Blood Flow. Metab. 2007, 27, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Rizvi, A.; Tang, X.L.; Manchikalapudi, S.; Takano, H.; Jadoon, A.K.; Wu, W.J.; Bolli, R. Nitric oxide triggers late preconditioning against myocardial infarction in conscious rabbits. Am. J. Physiol. 1997, 273, H2931–H2936. [Google Scholar] [PubMed]
- Bolli, R.; Bhatti, Z.A.; Tang, X.L.; Qiu, Y.; Zhang, Q.; Guo, Y.; Jadoon, A.K. Evidence that late preconditioning against myocardial stunning in conscious rabbits is triggered by the generation of nitric oxide. Circ. Res. 1997, 81, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Bencsik, P.; Kupai, K.; Giricz, Z.; Gorbe, A.; Pipis, J.; Murlasits, Z.; Kocsis, G.F.; Varga-Orvos, Z.; Puskas, L.G.; Csonka, C.; et al. Role of inos and peroxynitrite-matrix metalloproteinase-2 signaling in myocardial late preconditioning in rats. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H512–H518. [Google Scholar] [CrossRef] [PubMed]
- Imagawa, J.; Yellon, D.M.; Baxter, G.F. Pharmacological evidence that inducible nitric oxide synthase is a mediator of delayed preconditioning. Br. J. Pharmacol. 1999, 126, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R.; Manchikalapudi, S.; Tang, X.L.; Takano, H.; Qiu, Y.; Guo, Y.; Zhang, Q.; Jadoon, A.K. The protective effect of late preconditioning against myocardial stunning in conscious rabbits is mediated by nitric oxide synthase. Evidence that nitric oxide acts both as a trigger and as a mediator of the late phase of ischemic preconditioning. Circ. Res. 1997, 81, 1094–1107. [Google Scholar] [CrossRef] [PubMed]
- Afanas’ev, I. Ros and rns signaling in heart disorders: Could antioxidant treatment be successful? Oxid. Med. Cell. Longev. 2011, 2011, 293769. [Google Scholar] [CrossRef] [PubMed]
- Gelpi, R.J.; Morales, C.; Cohen, M.V.; Downey, J.M. Xanthine oxidase contributes to preconditioning’s preservation of left ventricular developed pressure in isolated rat heart: Developed pressure may not be an appropriate end-point for studies of preconditioning. Basic Res. Cardiol. 2002, 97, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Khadour, F.H.; Panas, D.; Ferdinandy, P.; Schulze, C.; Csont, T.; Lalu, M.M.; Wildhirt, S.M.; Schulz, R. Enhanced no and superoxide generation in dysfunctional hearts from endotoxemic rats. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1108–H1115. [Google Scholar] [CrossRef] [PubMed]
- Faggioni, R.; Gatti, S.; Demitri, M.T.; Delgado, R.; Echtenacher, B.; Gnocchi, P.; Heremans, H.; Ghezzi, P. Role of xanthine oxidase and reactive oxygen intermediates in LPS- and TNF-induced pulmonary edema. J. Lab. Clin. Med. 1994, 123, 394–399. [Google Scholar] [PubMed]
- Nduhirabandi, F.; Lamont, K.; Albertyn, Z.; Opie, L.H.; Lecour, S. Role of toll-like receptor 4 in melatonin-induced cardioprotection. J. Pineal Res. 2016, 60, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Tao, J.; del Monte, F.; Lee, K.H.; Li, L.; Picard, M.; Force, T.L.; Franke, T.F.; Hajjar, R.J.; Rosenzweig, A. Akt activation preserves cardiac function and prevents injury after transient cardiac ischemia in vivo. Circulation 2001, 104, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Talukder, M.A.; Varadharaj, S.; Velayutham, M.; Zweier, J.L. Early ischaemic preconditioning requires Akt- and Pka-mediated activation of enos via serine1176 phosphorylation. Cardiovasc. Res. 2013, 97, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.T.; Guo, Y.; Han, H.; Zhu, Y.; Bolli, R. An essential role of the JAK-stat pathway in ischemic preconditioning. Proc. Natl. Acad. Sci. USA 2001, 98, 9050–9055. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.T.; Guo, Y.; Zhu, Y.; Han, H.; Langenbach, R.; Dawn, B.; Bolli, R. Mechanism of cyclooxygenase-2 upregulation in late preconditioning. J. Mol. Cell. Cardiol. 2003, 35, 525–537. [Google Scholar] [CrossRef]
- Yu, H.M.; Zhi, J.L.; Cui, Y.; Tang, E.H.; Sun, S.N.; Feng, J.Q.; Chen, P.X. Role of the jak-stat pathway in protection of hydrogen peroxide preconditioning against apoptosis induced by oxidative stress in pc12 cells. Apoptosis 2006, 11, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Constante, M.; Layoun, A.; Santos, M.M. Contribution of stat3 and smad4 pathways to the regulation of hepcidin by opposing stimuli. Blood 2009, 113, 3593–3599. [Google Scholar] [CrossRef] [PubMed]
- Chaves de Souza, J.A.; Nogueira, A.V.; Chaves de Souza, P.P.; Kim, Y.J.; Silva Lobo, C.; Pimentel Lopes de Oliveira, G.J.; Cirelli, J.A.; Garlet, G.P.; Rossa, C., Jr. Socs3 expression correlates with severity of inflammation, expression of proinflammatory cytokines, and activation of stat3 and p38 mapk in LPS-induced inflammation in vivo. Mediat. Inflamm. 2013, 2013, 650812. [Google Scholar] [CrossRef] [PubMed]
- Platt, D.H.; Bartoli, M.; El-Remessy, A.B.; Al-Shabrawey, M.; Lemtalsi, T.; Fulton, D.; Caldwell, R.B. Peroxynitrite increases vegf expression in vascular endothelial cells via stat3. Free Radic. Biol. Med. 2005, 39, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G.; Musiolik, J.; Gedik, N.; Skyschally, A. Mitochondrial stat3 activation and cardioprotection by ischemic postconditioning in pigs with regional myocardial ischemia/reperfusion. Circ. Res. 2011, 109, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Csont, T.; Csonka, C.; Onody, A.; Gorbe, A.; Dux, L.; Schulz, R.; Baxter, G.F.; Ferdinandy, P. Nitrate tolerance does not increase production of peroxynitrite in the heart. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H69–H76. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, G.F.; Sarkozy, M.; Bencsik, P.; Pipicz, M.; Varga, Z.V.; Paloczi, J.; Csonka, C.; Ferdinandy, P.; Csont, T. Preconditioning protects the heart in a prolonged uremic condition. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1229–H1236. [Google Scholar] [CrossRef] [PubMed]
- Csont, T.; Csonka, C.; Kovacs, P.; Jancso, G.; Ferdinandy, P. Capsaicin-sensitive sensory neurons regulate myocardial nitric oxide and cgmp signaling. Eur. J. Pharmacol. 2003, 476, 107–113. [Google Scholar] [CrossRef]
- Csonka, C.; Szilvassy, Z.; Fulop, F.; Pali, T.; Blasig, I.E.; Tosaki, A.; Schulz, R.; Ferdinandy, P. Classic preconditioning decreases the harmful accumulation of nitric oxide during ischemia and reperfusion in rat hearts. Circulation 1999, 100, 2260–2266. [Google Scholar] [CrossRef] [PubMed]
- Csont, T.; Szilvassy, Z.; Fulop, F.; Nedeianu, S.; Pali, T.; Tosaki, A.; Dux, L.; Ferdinandy, P. Direct myocardial anti-ischaemic effect of gtn in both nitrate-tolerant and nontolerant rats: A cyclic GMP-independent activation of katp. Br. J. Pharmacol. 1999, 128, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Csont, T.; Pali, T.; Szilvassy, Z.; Ferdinandy, P. Lack of correlation between myocardial nitric oxide and cyclic guanosine monophosphate content in both nitrate-tolerant and -nontolerant rats. Biochem. Pharmacol. 1998, 56, 1139–1144. [Google Scholar] [CrossRef]
- Csonka, C.; Pali, T.; Bencsik, P.; Gorbe, A.; Ferdinandy, P.; Csont, T. Measurement of no in biological samples. Br. J. Pharmacol. 2015, 172, 1620–1632. [Google Scholar] [CrossRef] [PubMed]
- Csont, T.; Bereczki, E.; Bencsik, P.; Fodor, G.; Gorbe, A.; Zvara, A.; Csonka, C.; Puskas, L.G.; Santha, M.; Ferdinandy, P. Hypercholesterolemia increases myocardial oxidative and nitrosative stress thereby leading to cardiac dysfunction in apob-100 transgenic mice. Cardiovasc. Res. 2007, 76, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Downey, J.M.; Hearse, D.J.; Yellon, D.M. The role of xanthine oxidase during myocardial ischemia in several species including man. J. Mol. Cell. Cardiol. 1988, 20, 55–63. [Google Scholar] [CrossRef]
- Beckman, J.S.; Parks, D.A.; Pearson, J.D.; Marshall, P.A.; Freeman, B.A. A sensitive fluorometric assay for measuring xanthine dehydrogenase and oxidase in tissues. Free Radic. Biol. Med. 1989, 6, 607–615. [Google Scholar] [CrossRef]
- Fekete, V.; Murlasits, Z.; Aypar, E.; Bencsik, P.; Sarkozy, M.; Szenasi, G.; Ferdinandy, P.; Csont, T. Myocardial postconditioning is lost in vascular nitrate tolerance. J. Cardiovasc. Pharmacol. 2013, 62, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, G.F.; Pipis, J.; Fekete, V.; Kovacs-Simon, A.; Odendaal, L.; Molnar, E.; Giricz, Z.; Janaky, T.; van Rooyen, J.; Csont, T.; et al. Lovastatin interferes with the infarct size-limiting effect of ischemic preconditioning and postconditioning in rat hearts. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H2406–H2409. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | LPS | |
|---|---|---|
| Animal weight (g) | 307 ± 5 | 301 ± 5 |
| Heart wet weight (mg) | 928 ± 25 | 927 ± 28 |
| Basal heart rate (bpm) | 299 ± 7 | 295 ± 11 |
| LDH release (U/min) | ||
| Before ischemia | 1.1 ± 0.2 | 1.5 ± 0.3 |
| After ischemia | 5.8 ± 0.7 * | 2.3 ± 0.4 # |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pipicz, M.; Kocsis, G.F.; Sárváry-Arantes, L.; Bencsik, P.; Varga, Z.V.; Ferdinandy, P.; Csont, T. Low-Dose Endotoxin Induces Late Preconditioning, Increases Peroxynitrite Formation, and Activates STAT3 in the Rat Heart. Molecules 2017, 22, 433. https://doi.org/10.3390/molecules22030433
Pipicz M, Kocsis GF, Sárváry-Arantes L, Bencsik P, Varga ZV, Ferdinandy P, Csont T. Low-Dose Endotoxin Induces Late Preconditioning, Increases Peroxynitrite Formation, and Activates STAT3 in the Rat Heart. Molecules. 2017; 22(3):433. https://doi.org/10.3390/molecules22030433
Chicago/Turabian StylePipicz, Márton, Gabriella F. Kocsis, László Sárváry-Arantes, Péter Bencsik, Zoltán V. Varga, Péter Ferdinandy, and Tamás Csont. 2017. "Low-Dose Endotoxin Induces Late Preconditioning, Increases Peroxynitrite Formation, and Activates STAT3 in the Rat Heart" Molecules 22, no. 3: 433. https://doi.org/10.3390/molecules22030433